0022-538X/81/070229-09$02.00/0 Vol. 39, No. 1
Attachment of Avidin-Coupled Spheres to Linear and Circular
Forms of Mengovirus Double-Stranded RNA
GEORGE B.THORNTON,' DONALD L.ROBBERSON,' AND RALPH B.ARLINGHAUS3* Department of Biology, Abilene Christian University, Abilene, Texas79699,1 and Department ofMolecular
Biology'
andDepartment of TumorVirology,3 The University of Texas System Cancer Center M. D.Anderson Hospital and TumorInstitute, Houston, Texas 77030
Received23January 1981/Accepted1 April1981
Picomavirus-infected cells contain a double-stranded RNA (replicative forn
[RF] RNA), composed of viral genomic RNA hydrogen bonded to cRNA of
similarnucleotide length.Mengovirus RFRNA reacted with a succinimide ester
ofbiotinwasshownby electronmicroscopy to bind avidin-coupled
polymethac-rylate spheres. These binding sites are taken to indicate the presence of VPg
protein molecules by methodspreviouslyappliedto poliovirus RF RNA (Richards
etal., Proc. Natl. Acad.Sci. U.S.A. 76:676-680, 1979). One sphere was bound at
or very near one terminus of linear RF molecules while a second sphere was
boundat asitewhich was 1 to 4% of thegenome length from the other terminus.
Assignment of VPgpositions was limited bythe physicaldimension (ca. 60-nm
diameter) of the heavy metal-contrasted sphere observedbyelectronmicroscopy.
Athirdsite of sphere binding was detected at a lower frequency of occurrence at
asite whichwas 10 to20% of thegenomelengthfromone orthe other terminus.
Circular RF RNA molecules were also detected with two spheres attached at
juxtaposed sites. The termini of the linear RF RNA were located within the
circular structuresby sphere attachmentat sites whichwere very close to and
obscureda
nucleic
acid projection whichwehave described to occur on circularmengovirus RF RNA (D. L. Robberson, M. V. Marshall, G. B. Thornton, and R.
B.Arlinghaus, manuscript submitted forpublication). CsCl gradient fractions of
RNA reacted with
avidin-coupled
sphereswerehighly
enriched incircularstruc-tures.
The
genomic
RNAofpicornaviruses
containsasmall basicprotein
covalently
attachedto its5' terminus(6-8, 10,14,16, 19,20).Thisprotein,
VPg, isalso attachedtothe 5'ends of
intracel-lular forms of viral RNA involvedinviral RNA
replication. Double-stranded
poliovirus
RNA(replicative
form [RF] RNA) contains VPg atthe 5' end of the
plus
and minus strands(6, 13).Onewaytodetect
protein
boundtosuchstruc-turesis
by
useofelectron-opaque
avidin-coupled
spheres (16). In such
experiments,
biotin ischemically
linked to theVPg-RNA
complex.
The high affinity of avidin for biotin is then
utilized in the attachment of the
electron-opaque avidin-coupled spheres to the protein bound to viral RNA.Richards etal. (16) showed
that avidin-coupled spheres were attached to
theplusstrand atonlyoneend ofpoliovirus RF thathadbeen reacted with biotin.
Ourstudies withmengovirusRF have shown
that purified preparations of RF contain both
circular and linear forms of viral double-stranded RNA (M. V. Marshall, R. B.
Arling-haus, and D. Robberson, J. Cell Biol. 70:248a,
1976;Robbersonetal., manuscript submittedfor
publication).
The circulardouble-strandedviralRNAs are of genomic
length,
contain a shortprojection, andarenoncovalent associations of
the linearforms. Similar circularstructures were
firstreported forRF RNAof
encephalomyocar-ditis virus
by Agol
andhiscolleagues (1,2)
andsubsequently shown to be noncovalent
struc-tures (18). In this
study,
theavidin-coupled
sphere
technique
(11)(previously applied
topoliovirusRNA) was usedtodemonstrate that
both ends of biotin-linked
mengovirus
RF canbind
avidin-coupled spheres,
indicating
the pres-ence ofVPg-like
molecules at or near theter-mini,and toidentifythe locationof the termini
of linear RF within the circularstructures.
MATERIALS AND METHODS
Cells and virus. BHK-21clone 13 cellsweregrown andpropagatedinroller bottlecultures,asdescribed
(24). Mengovirus was plaque purified by
Royce
Z. Lockhart, Jr. (Du Pont Laboratories,Wilmington,
229
on November 10, 2019 by guest
http://jvi.asm.org/
230 THORNTON, ROBBERSON, AND ARLINGHAUS
Del.); theinitial stock of viruswasobtained from his laboratory. Inatypicalexperiment,tworollerbottles, having attained75 to90%confluencyof5x 108cells, wereinfected with 10 to50PFUpercell. Fresh me-dium containing 5,ug ofactinomycin D per ml was added2.5h afterinfection,and theisotope (1 mCi-of
[3H]uridine in25ml ofmedium)wasaddedat3h after infection. At5 to 6hafterinfection,the virus-infected cells were harvested by scraping and collected by
centrifugation.
Isolation andpurificationofmengovirusRF. Virus-infectedcellswereresuspended inahypotonic
buffer solution(0.02 M Tris[pH7.5],0.0036MCaCl2, and0.0014MMgCl2)andallowedtoswellfor5minat
00C.Thecellswerethen mixed withanequalvolume of hypertonic buffer (1.0% Triton X-100, 0.0036 M CaCl2, 0.01 MMgCl2, 0.5 M sucrose,0.25MKCI,0.04 MTris, pH 7.5)and homogenized withaloose-fitting pestle10times. Thebroken cellsuspensionwas cen-trifugedat5,000 rpmin aSorvall SS-34rotorfor 10 min, and the supernatant fluid wasdecanted. Cyto-plasmic RNAwasextracted from thesupernatant fluid by sodiumdodecyl sulfate(SDS)-chloroform-phenol,
asdescribed(15). The RNAwasmade0.2M inlithium acetate(pH 5.1) andprecipitatedbytheaddition of2 volumes of ethanolat-20°Covernight.
Mengovirus RFwasseparated fromsingle-stranded
virus-specific and other cellular RNAs based onits differential solubilityinhigh-saltsolutions (3, 4, 22). Thecytoplasmic RNAwasdissolved in1mlof TLE (10 mMTris-hydrochloride [pH7.5], 0.10MLiCl, 1 mM EDTA), andanequal volume of4MLiClwas addedtothat solution. Theresulting2MLiClsolution wasstoredovernightat4°C.Theprecipitate, contain-ingsingle-stranded RNAs, wasremovedby centrifu-gationat3,000xgfor45min. The2MLiCl-soluble
supernatant fluid,containing thedouble-stranded RF, DNA, and small RNAs,wasmade 1 M inLiClby the addition ofanequalamountof sterile deionizedwater and ethanol precipitated overnight at -20°C. The ethanol precipitate wassuspended in TLE-0.5% so-diumdodecylsulfate, made 2 MinLiCl, and kept at 4°C overnight. The precipitate, if any,wasremoved bycentrifugation,and thesoluble RNA was precipi-tated withethanol and suspended once again in TLE thatwasthen made2M inLiCl. After the third LiCl precipitation, the sample was ethanol precipitated and
suspendedinTLE-0.1% SDS and stored at -70°C. The2MLiCl-soluble RNA (in 0.5 ml of TLE-0.1%
SDS)wasmixed withanequal volume of 5% sucrose-NLE-SDS (10 mM sodium acetate [pH5.5], 100 mM LiCl, 1mMEDTA, and 0.1% SDS) and layered onto a5 to25%sucrose-NLE-SDS gradient. Water (3.5 ml) waslayeredontop of the sample layer and the gradient was placed in a vertical rotor (TVA-865; Du Pont,
Wilmington,Del.) and centrifuged for 75 min at 50,000 rpmat40C.The gradient was collected in 1-ml frac-tionsandassayed forradioactivity, and the peak frac-tions, which sedimented at 20S, were collected and precipitated by ethanol.
DNase Itreatmentof RF.RFobtainedby velocity sedimentation centrifugation asdescribed above was suspended in 0.5mlof NTM (100 mM NaCl, 5 mM
MgCl2, and 10 mM Tris[pH7.8]). DNaseI,obtained from Calbiochem-Hoescht, Behring, La Jolla, Calif.,
was added to a final concentration of50 U/ml and incubated for 30to60minat250C. The reactionwas terminatedby the addition of 50,ul of 0.1 M EDTA (pH 7.0). An equal volume of 5% sucrose-NLE-SDS wasaddedtothe terminated reactionmixture, and the resulting solutionwaslayeredonto a5to25% sucrose-NLE-SDS gradient and subjected to velocity sedi-mentation in a vertical rotor for 75 minat50,000 rpm (4°C),asdescribed above. Thegradientwas fraction-ated, and the peak fractions, which sedimented at20S, werepooled andprecipitated with ethanol.
Bondingofavidin-coupledspherestoRF. Av-idin-coupled sphereswereboundtoRF moleculesas describedby Richardsetal.(16). Briefly, RF prepared asdescribedwassuspendedin0.1Msodium bicarbon-ate toyieldafinal concentration of 25,ug/ml. A
100-pl
sampleof the RFsolutionwasmixed with 10,lIof dimethyl formamidecontaining 100Mug of theN-hy-droxysuccinimidylesterof biotin(kindly provided by JerryManning, University ofCalifornia, Irvine) and incubated at 4°C for 3 h. This mixture was then
dialyzedovernight against 1 MNaCl-0.1 M HEPES
(N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid) (pH 7.5). To the dialysatewasadded30,ul of the
avidin-coupled spheres (20mg/ml), and the suspen-sionwasincubatedat40C for24h. Thepreparationof avidin-coupled spheres boundtoRFwasthenlayered onto aCsCl stepgradient (2.45 ml of CsCl in0.1 M HEPES [pH7.5],p = 1.52; 2.45ml ofCsCl in0.1 M HEPES[pH7.5],p =1.23) andsubjectedto centrifu-gation inanSW50.1rotor at45,000 rpmfor22 hat
40C.Theavidin-coupled spheresandspheres bound tothe RFmolecules formed an opalescentband ap-proximately one-third of the distance from the top of thegradient. Theopalescent band contained 77% of theuridine-labeled RFRNA; theremaining 23%was found in the pellet. The band wasremoved with a syringe, dialyzedagainstTNEbuffer (10 mM Tris[pH
7], 100mMNaCl, and1 mMEDTA), and examined byelectronmicroscopy.
Electronmicroscopy.Samplesfrom the CsCl gra-dient fractions containing RF reacted with poly-methacrylate sphereswerediluted in0.1MNaCl-0.1 MTris[pH8.0]-0.01 M EDTAtoafinal RNA con-centration of 0.5 Mug/ml and prepared for electron microscopy by usingthe aqueous basic protein
tech-nique of Kleinschmidt as described previously by Davisetal.(5). Gridswerestained with uranyl acetate asdescribed(17) and rotary shadowed with Pt:Pd (80: 20) after which theywereexamined in a Philips 300 electron microscope. Photographswere taken on 35-mmflrm,andlengths of projectedimagesof the mol-eculesweredetermined withaNumonics 1224 digitizer interfaced withaHewlett-Packard 9825A computer.
RESULTS
Purification ofmengovirus RF.
Mengovi-rusdouble-stranded RNA, termed RF, was
iso-latedfrom infected BHK-21 clone 13 cells and
purified essentially as described previously by
Spector
andBaltimore (22).Theisolationof the RF is basedonitssolubility in2MLiCl.Men-govirus RF, subjected to three LiCl
precipita-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
o fO~~~~~~~~~~o
0 10 2 00 1 0 3
andthelatterwas8entrifugedinaparallelgradient.(B)The peak(A)was
d+
DNaseI
centrifuge0 onti
TREATMENT(LIt
i 04 It II
10 I
I
6-0
4~~~~14
00
20
0 10 20
0o
10 20 30FRACTION NUMBER
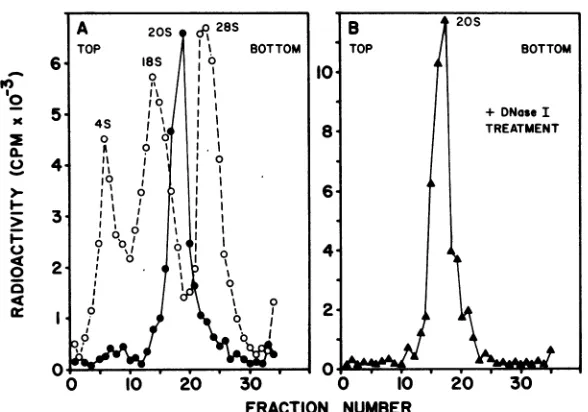
FIG. 1. Sucrose density gradient centrifugation of mengovirus-specific RF RNA. (A) ViralspecificRNA waslabeled withf8HJuridinein thepresenceofactinomycinD,and 2 MLiCi-soluble RNAwasappliedtoa
5to25%linear-sucrosegradient. Theviral RNAtracing(@)wascoplottedwithacellular RNAmarker(0),
and the latterwascentrifuigedinaparallel gradient. (B) The208peak(A)waspooled,DNaseI-treated, and
centrifugedon a5to25% linearsucrosegradient.
tions, sedimented as a
homogenous
peak at arateof 20S
(Fig.
1A). Treatment ofRF,
whichhad been isolated fromasucrosegradient, with
DNase I hadnoeffectonitssedimentationrate
(Fig.
1B).
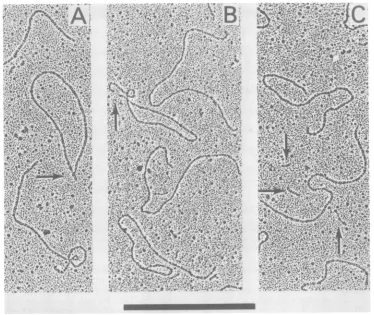
DNase-treated mengovirus RF molecules
were isolatedasdescribed above and examined
by
electronmicroscopy.
Several formswereob-served
(Fig. 2):
(i)linear RF molecules(Fig. 2A,
B, and
C), (ii)
circular moleculespossessing
ashortprojection
(Fig.
2A,arrow),
and(iii)
linearmolecules with their ends
opposed
(Fig. 2B,
ar-row, and
C,
upperpart).
Although
somemole-cules smaller than
genomic
length
werepresent(Fig.
2C, arrows),
thepreparation
of RFap-peared to be rather
homogeneous
inlength
ineach of the three
forns.
Beforesphere
attach-ment and
CsCl
gradient
fractionation(see
be-low),
the RFpreparation
contained 7.1% circularduplexes
withaprojection,
6.0% linearduplexes
withopposed
ends,
andonly
1.0%circulardu-plexes withoutadetectable
projection;
atotal of298moleculeswasexamined
by
electronmicros-copy.
Binding
ofavidin-coupled spheres
toRFmolecules. The
genome-linked
protein,
VPg,
has been found to be
covalently
linked to thevirus-specific RNAs of
poliovirus (6,
7, 10, 16,25), the genomicRNAs of foot-and-mouth
dis-easevirus(9,20),
encephalomyocarditis
virus(8,
10), mengovirus
(14),
and calicivirus(21).
Inthese
studies,
bothbiochemical andelectronmi-croscopictechniqueshave beenemployed. Using
the avidin-coupled sphere method ofRichards
etal. (16), weinitiallysought to locate VPgon
mengoviruslinearRFmoleculesandcircular RF
molecules. Since VPg is expectedtobe found at
or near the termini, its presence would then allowustoidentifytheterminiof themolecules
in thecircularforms,a majorobjectivein these
studies.
An excess of theN-hydroxysuccinimidylester
ofbiotin was reacted with mengovirus RF by
meansoftheprotein VPg. Themengovirus
RF-VPg-biotin complex was thenboundto
avidin-coupledpolymethacrylate spheres,asdescribed
above.Electron microscopic examinationofthe
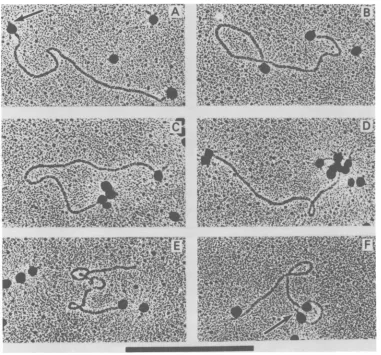
RF-bound spheres revealed several classes of
linear molecules: (i) full length RF molecules
possessing spheresonbothtermini (Fig. 3A and
B), (ii)molecules possessingasinglesphere on
oneendandmultiple spheresontheother(Fig.
3C), (iii) molecules possessing multiple spheres
on both ends (Fig. 3D), (iv) molecules ofless
than genomiclength possessingasingle sphere
(Fig. 3E), and (v) infrequently occurring
mole-culespossessing intemallyboundspheres (Fig.
3F).Itshouldbeemphasized that when multiple
sphere attachment was involved, only one
sphere of theaggregate wasfoundtobein direct
physical contact with the RF RNA molecule.
Furthernore, in linear RF molecules, it was
frequently observed thataspherewasattached
at oneterminuswhileasecondsphere attached
on November 10, 2019 by guest
http://jvi.asm.org/
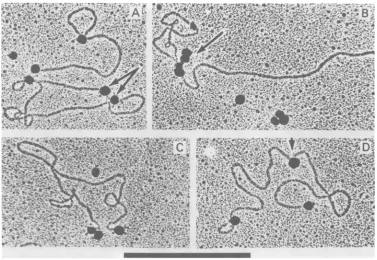
[image:3.493.102.396.62.268.2]232 THORNTON, ROBBERSON, AND ARLINGHAUS
FIG. 2. ElectronmicrographsofmengovirusRFRNA. Both linear and circular RF RNA moleculeswere detected. The circular molecules typically contained ashortprojection (indicated by the arrow inA). In
addition, linearmolecules withopposedendswerealsodetected(indicated bythearrowinB;upperpart
of
C).Double-strandedmolecules much shorter than genomelengthareindicatedbythearrowsin C.Bar, 1,m.
verynearbutnotpreciselyatthe other terminus, i.e.,oneterminuswasseentoextendbeyond the site of sphere attachment (Fig. 3A and F,
ar-rows). This terminus extends approximately
1.0% of thegenomelengthbeyond the boundary of the sphere. Furthermore, a second site of
sphere attachmentappearedatadistance of10 to 20% of the genome length from one of the
termini(Fig. 3Datright and Fig. 3F) in
approx-imately 6% of the linear duplexes and13% of the
circular duplexes with one or more attached
spheres, which may reflect the presence ofan
additionalVPg-like moleculeor someother
pro-teinboundtotheRF RNA.
Severalunusualfeaturesrequirefurther com-ment.Therewere numerousunattachedas well
asattachedspheres which possessedsmall
pro-tuberancesthatweredistinguishedfrom the
pro-jections on circular structures. A small
protu-berance is evident on the single sphere in the
lowerrightcornerof Fig.3C.Thesourceof these
protuberances is unknown, but it should be
noted thatvery smallRNAs havebeenobserved in
preparations
ofmengovirus
andmayaccountforthese
protuberances
(manuscript
submittedfor
publication).
Theseprotuberances
werenotobservedin the
poliovirus
studies describedby
Richards et al.
(J.
Manning,
personal
commu-nication). A second unusual
phenomenon
wasthe
clumping
ofavidin-coupled spheres.
Theclumps of
avidin-coupled
spheres
wererarely
foundexcept when boundto
mengovirus
RF. Arandom sampling of 212 spheres in the
back-groundof theCsCl
gradient
fractioncontaining
RFrevealedthat 73%occurredas
single
nonag-gregated spheres, 17% as aggregates of two
spheres, 6% as aggregatesofthree
spheres,
2%asaggregates of four spheres,and 2% as aggre-gates of five spheres. The significance of the
clumps ofspheres boundtooneend of the RF
remainstobe determined.However, the
clump-ing of spherical aggregates of antibodies was
reported by Wuet al. (25) in antibody-treated
2,4-dinitrophenol derivatives of
poliovirus
RFJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.62.436.67.382.2]VOL. 39,1981
(seebelow).
Having demonstrated thatbothtermini,
bear-ing VPg, could be labeledwith the
avidin-cou-pled spheres, itwasofinterest todeterminethe
location of thetermini in circular forms of
men-govirus RF. As mentioned above, mengovirus
circular RF molecules possessed a single short
projection per circle. In 73% of the molecules
scored (sphere-bound circles)theprojection was
obscured. The termini of the RF
molecules,
therefore, are at or in close proximity to the
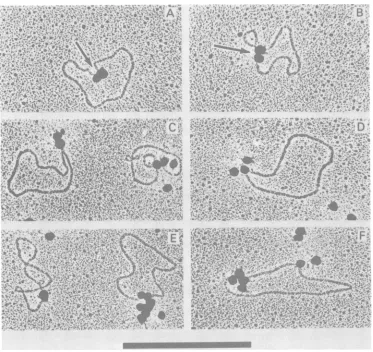
projection.Figure 4A presents a typical circular
RFmolecule withtwospheresattached in close
proximity but lying on the circular contour of
MENGOVIRUS RF RNA 233
the RFmolecule. Most circularformshad two
spheres attached (Fig. 4A and B). There were
also circular formspossessing a single attached
sphere (Fig. 4D and E), multiple attached
spheres (Fig. 4E), and
internally
boundspheres(Fig. 4F).Inthose circular structures possessing
twoor more spheres which indicated the
posi-tions of the termini of linear RF within the
circularstructures,itwasfound that the termini
were physicallyseparated by only a small
un-measurable distance on the circular contour
(Fig. 4A, B, C,and F). This would indicate that
in the circularforms of RF, the VPg molecules
at the termini of the linear molecules are not
FIG. 3. Electronmicrographs ofmengoviruslinear RF RNA molecules with attachedpolymethacrylate spheres. (AandB)A single sphereis attachedneareach terminusof the linear molecules. Thearrow (A) depictsoneterminus whichprotrudes slightly beyondtheboundary ofthesphere. (C) An aggregate of four spheres is attached to one terminus and a single sphere is attached to the otherterminus of the linear molecule.(D) Twospheresareattachedatoneterminusandanaggregateof six spheres is attachednearbut not atthe otherterminus,whereasingle sphereis attached. (E)Asphereis attached atonlyoneterminus. (F) Onesphere isattachedneareach terminusaswellas asinglesite locatedonthe linear molecule. The arrow(F)depictstheterminusofthe molecule whichprotrudesbeyondtheboundary of the sphere. Bar,1,tm.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.58.439.216.571.2]234 THORNTON, ROBBERSON, AND ARLINGHAUS
FIG.4. Electronmicrographs of mengoviruscircular RF RNA molecules with attachedelectron-opaque polymethacrylate spheres. Samples wereprepared for electron microscopy by the aqueous technique of Kleinschmidtdescribedpreviously byDavisetal.(5)and contrastedbyrotaryshadowingwith Pt:Pd.(Aand B) Two spheres are attached at apparently adjacent sites indicated by arrows on each ofthe circular molecules. (C)Twospheresjoinedin tandem appearattachedat asinglesiteonthe circular molecules. (D
andE[atleft])A single sphereis attachedtothe circular molecules. (E [at right])Anaggregateofseven
spheres is attachedat asinglesiteofthe circular molecule. (F)An aggregateof four spheresis attachedat onesite, andasingle sphereisattachedat asecond siteonthesamecircularmolecule.Bar, I,um.
juxtaposedbutdo lieinclose proximity toone
another.
Linearandcircular dimersofmengovirusRF RNAhave alsobeendetectedatalow frequency
ofoccurrence (D. Robberson, M. Marshall, G.
Thornton, and R. Arlinghaus, manuscript
sub-mitted for publication). These species also re-actedwithavidin-coupled spherestogive struc-tures presented in Fig. 5. The circular dimers frequently containedtwoor moresites of sphere
attachment (Fig. 5A), whereas linear diners
most frequently were seen to contain sites of
sphere attachment ator nearthe termini (Fig.
5C)aswellasatasiteinternally locatedonthe molecule (Fig. 5D). A circular monomer was
occasionally seenattachedtothe terminus ofa
linear moleculethroughone ormore
avidin-cou-pledspheres(Fig.5B). In such cases, the
projec-tionon thecircular RFmoleculewasobscured
bysphere attachment and, byinference, must
belocatednearthesite ofsphere attachment.
Table 1summarizes thefrequenciesof
differ-ent structures observed in the avidin-coupled
sphere boundRFpreparation.Ofsignificanceis
the fact that 63% of the linear RF molecules
possessed spheres on both ends, indicating at
least oneVPgmolecule is at or neareach
ter-minus.However,98% of the linear RFmolecules
had one or more spheres attached. Of the
cir-cular forms, 98% of the molecules contained
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.68.441.65.417.2]MENGOVIRUS RF RNA 235
FIG. 5. Electron micrographs of mengovirus circular and linear oligomeric RF RNA molecules with attachedpolymethacrylate spheres. (A)The circular dimerformhastwospheres attached near the same site
(indicatedby arrows)andasingle sphereattachedat aremotesite. A linearmonomerwithasphere at each terminus is shownatthetopofthemicrograph. (B)Acircularmonomerisjoinedto alinear monomerthrough anaggregateof fourspheres in theregion indicated byarrow. (C) A linear dimer has one sphere attached near oneterminus andtwospheres, close to each other, attached near the other terminus. (D) One sphere is attachedateach terminus andonesphere is attachedat aninternal site indicated by thearrow on the linear dimer molecule.Bar,1
tum.
spheres, 43% contained one sphere, and 50%
containedtwospheres.Itshould beemphasized
that
sphere
attachmentnearly always occurredat asite thatobscured the projection. Also, the
frequency
of circular forms in these preparationswas
significantly
higher than in preparationsnotreacted with biotin-bound avidin-coupled
spheres. One possible explanation for this
occur-rence is that the reaction withthe
avidin-cou-pled spheresmayhave inducedagreater
stabil-ityin circular forms, preventing their
dissocia-tionuponspreadingfor theelectronmicroscopy.
DISCUSSION
We have been ableto demonstratethe
pres-enceofprotein, presumably VPg, onboth
ter-mini ofmengovirus RF by the avidin-coupled
sphere technique of Manning et al. (11).
Pre-vious studieswith this method ofpoliovirusRF
(16) revealedapreferentiallabelingofonlyone
end of the RFmoleculewith theavidin-coupled
spheres, the endpossessing the 5' terminus of
thepositivestrand. Wu etal. (25),
however, by
using adifferent method
(that
ofreacting
anti-2,4-dinitrophenol antibodies with the
2,4-dini-trophenol-derivatized VPg of RF molecules),
wereabletolabel both ends of
poliovirus
RF. Itwas suggested by Richardset al. (16) that the
inabilitytolabel the 5' end of the negative strand
of the poliovirus RF might be due to one of
several
factors,
themostprobable
of whichwere(i) that thepolyadenylic acidsequence onthe 3'
end of the positive strand is longer than the
polyuridylic acid sequenceon the 5'endofthe
negativestrand, whichmight maskVPgatthe
5' end of thenegativestrand, preventing
esteri-ficationwithbiotin,and (ii) that there are two
different VPg
molecules,
one capable ofbeing
esterified tobiotinmore
readily.
Both of thesepossibilities may, in
fact,
beresponsible
forat-tachment of onlya single sphere topoliovirus
RF. Infact, the studies described in thispaper
wereinitiallyundertaken,inpart, because
men-govirus has been shown topossess a short
se-quence of
polyadenylic
acid on its 3' terminus (12,23).Withthis inmind,
wefelt thatwecould label bothtermini of themengovirus
RF withouthavingtocontend witha
long
polyadenylic
acid39,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.493.58.435.63.323.2]236 THORNTON, ROBBERSON, AND ARLINGHAUS
TABLE 1. Frequencies of mengovirus RFRNAs with attachedavidin-coupledpolynethacrylate
spheresa
No.of %Of
to-RF RNA mole-
-cules
Genomic-lengthmolecules
Linearforms 43 45
Circular forms 44 46
Opposedends 8 9
Genomic-lengthlinears
Nospheres 1 2
Sphereson oneendonly 15 35 Spheres on both ends 27 63
Genomic-lengthcircles
One sphere,projection absent 19 43 Twoadjacent spheres, 13 30
projection absent
Two adjacent spheres, 9 20 projection absent (loops)
Spherespresentonprojection 2 5
Nospheres 1 2
aThe frequencies of
internal
sphere attachment mentioned in the text have been omitted from this tabulation.sequence similar to that found in poliovirus
RNA. Asreported here, 65% of the mengovirus
RF molecules werelabeled atboth
termini,
incontrast tothe labeling of 13% of the RF
mole-cules ofpoliovirus (16). Althoughwewereable
tolabel both endsofapicornaviralRF
possess-ingashortpolyadenylicsequence,itcannotbe
ruledoutthat therearechemicallydifferent VPg
moleculespossessing different labeling affinities
(see
below).
Infact,
Kingetal. (9) haverecentlydemonstrated that thereare twochemically
dif-ferent VPg's in foot-and-mouth disease virus
RNApreparations.
Wu et al. (25) have described a class of RF
molecules possessingaproteinaggregate on one
end of poliovirus RF and have termed these
complex molecules.Mostoftheseprotein
aggre-gates could be removed by banding the RF in CsCl-guanidine hydrochloride gradients. These
additionalproteinmolecules were not,therefore,
covalentlybound.
We report here the occurrence ofcomplexRF
moleculesin preparations of mengovirusRF and
note that there appears to be a preferential
labeling of one or the other end of these
mole-cules with two or more spheres; however, the observation thatonly onesphereof anaggregate
lies in contact with the RF molecule suggests
thatonlyoneVPg molecule hasreactedwith a
sphereoraggregateofspheres at this site.Other
VPg molecules may infact be nearby but are
notdetectedas aresult of physicalexclusion by
the sizes ofspheresthat were utilized. Whether or not the VPg or protein molecules on both ends of the RF are the same or different remains
to be determined. It is also possible that one
typeofVPg molecule,orprotein, has agreater
affinityfor one end of the RF molecule than the
other.
A second major objective of this work was
locating the termini of linear RF molecules
within the mengovirus circular structures
pre-viously described (Marshall etal.,J. Cell Biol.
70:284a, 1976) (Fig. 2).We havepresented
evi-dence that the sphereswhich are boundto the
circularstructuresobscure a smallprojectionwe
have previously observed to occur on circular
molecules(Marshalletal.,J. Cell. Biol.70:284a,
1976). The observation that the spheres are
joinedtothe circularRF moleculesat two
dis-tinct butclosely separated pointsand that most circles possess two or more suchspheres lying
sideby sidesuggeststhe close relative position
of theVPg's but excludes the
possibility
that theVPgmolecules are
juxtaposed
withinmostcir-cularstructures.
Another observation which must be
ac-counted for is the increasedfrequencyof circular
RF RNAinthe
preparations
reacted withavi-din-coupled spheres.Itappearedthat the
reac-tion increased thestability of the
circles,
possibly
by
cross-linking
theVPg
moleculesoreffecting
a more stable structure which is abletowith-stand the spreading forces during preparation
forelectronrnicroscopy.
ACKNOWLEDGMENTS
This research was supported by Public Health Service grantsCA-25465, CA-16672 and CA-16527 from the National InstitutesofHealth and grantsG-429and G-841 from The Robert A.Welch Foundation. G.B.T.wassupported byfunds from the Research Council ofAbilene ChristianUniversity.
We thank Susan Berkley, CarolynPeterson, and James Syrewiczforexcellenttechnical assistance and Rebecca Bazer forassistance in manuscript preparation. We especially thank Jerry Manningforprovidingreagents withwhichtodo the biotincross-linking and the avidin-coupled sphere binding.
LITERATURE CITED
1.Agol, V.,Y.Drygin,L.Romanova,and A.Bagdonev.
1970.Circularstructures inpreparationsofreplicative formofEMCvirus RNA. FEBS Lett 8:13-16.
2. Agol,V. I., L.L. Romanova,I. M. Cumakov, L. D. Dunfevskaya, and A. A. Bogdanov. 1972.Circularity andcross-linkinginpreparations ofreplicativeformof encephalomyocarditis virus RNA. J. Mol. Biol. 22:77-89.
3. Baltimore,D., and M. Girard. 1966. An intermediatein
thesynthesis of poliovirus RNA. Proc. Natl. Acad. Sci. U.S.A.56:741-748.
4. Bishop, J. M., and G. Koch. 1967. Purification and characterization of poliovirus-induced infectious dou-ble-strandedRNA.J.Biol.Chem.242:1736-1743.
5. Davis,R.W.,M.Simon,and N.Davidson.1971.
Elec-tron microscopic heteroduplex methods formapping regions ofbase sequence homology innucleic acids. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
MethodsEnzymol.21:413-438.
6. Flanagan, J. B., R. F. Petersson, V. Ambros, M. Hewlett, and D. Baltimore. 1977. Covalent linkage of a protein to a definednucleotide sequence at the 5' terminusofvirionandreplicativeintermediate RNAs ofpoliovirus. Proc. Natl. Acad. Sci. U.S.A. 74:961-965. 7. Golini, F., A. Nomoto, and E. Wimmer. 1978. The genome-linked protein ofpicornaviruses. IV. Difference in theVPgs of EMC virus and poliovirus as evidence that thegenome-linked proteins are virus coded. Virol-ogy 89:112-118.
8. Hruby,D.E.,and W. K. Roberts.1978. Encephalomy-ocarditis virus RNA. HI. Presence of a genome-associ-atedprotein.J. Virol. 25:413-415.
9. King,A. M. Q., D. V. Sangar, T. J. R. Homes, and F. Brown. 1980. Heterogeneity of genome-linked protein offoot-and-mouth disease virus. J. Virol. 34:627-634. 10.Lee,Y.F.,A.Monoto,B.M.Detjen, andE.Wimmer.
1977. Aproteincovalently linked topoliovirus genome RNA. Proc.Natl. Acad. Sci. U.S.A. 74:59-63. 11. Manning,J. E., N. D. Hershey, T. R. Broker, M.
Pellegini, H. K. Mitchell, and N. Davison. 1975. A newmethod of in situ hybridization. Chromosoma 53: 107-117.
12. Marshall, ILV., and R. B.Arlinghaus.1976. Polyad-enylicacid in the genomic RNA of mengovirus. J. Virol. 19:743-745.
13. Nomoto, A.,B. M.Detjen,R.Pozzati,and E. Wimmer. 1977. The location ofpolio genome protein in viral RNAs and its implications for RNAsynthesis.Nature (London) 268:208-213.
14. Perez-Berloff,R., and M. Gander.1978.In vitro
trans-lationofmengovirus RNAdeprivedof theterminally linked(capping?) protein.FEBSLett.96:306-312. 15. Perry,R.P.,J.LaTorre,D. F.Kelly,and J. R.
Green-bert.1972.On thelability ofpoly(A)sequences during
extraction of messenger RNA from polyribosomes. Biochim.Biophys.Acta254:493-495.
16. Richards, 0.C.,E.Ehrenfeld, andJ.Manning.1979.
Strand-specificattachment of avidin-apheres to double-stranded poliovirus RNA. Proc. Natl. Acad. Sci. U.S.A. 76:676-680.
17. Robberson, D.,Y.Aloni,andG. Atiardi.1971.Electron microscopicvisualization of mitochondrial RNA-DNA hybrids.J. Mol. Biol.55:267-270.
18. Romanova, LI.,and V. I.Agol.1979.Interconversion oflinearand circular forms of double stranded RNA of encephalomyocarditisvirus.Virology 93:574-577. 19.Rothberg,P.G.,T. J. R.Harris,A.Nomoto,and E.
Wimmer. 1978. 04-(5'-Uridylyl)-tyrosineis the bond between thegenome-linked protein and the RNA of poliovirus. Proc. Natl. Acad. Sci. U.S.A. 75:4868-4872. 20. Sanger, D. V., D. J.Rowlands, T. J. R. Harris, and F.
Brown.1977.Aprotein covalently-linkedto foot-and-mouthdisease virus RNA.Nature(London) 208:648-650.
21. Schaffer, F. L, D. W.Ehresmann,M. K.Kretz, and M.E.Soergel.1980.Aprotein, VPg, covalentlylinked to36S calicivirus RNA. J. Gen. Virol.47:215-220. 22. Spector,D., and D.Baltimore.1975.Polyadenylicacid
onpoliovirusRNA.ILPoly(A) on intracellular RNAs. J.Virol. 15:1418-1431.
23. Spector,D., and D. Baltimore. 1975.Poly(A) of men-govirus RNA. J. Virol. 16:1081-1084.
24. Syrewicz,J.J., R. B.Naso, C. S.Wang, and R. B. Arlinghaus. 1972. Purification oflarge amounts of murineribonucleicacid tumor virusesproduced inroller bottle cultures. Appl. Microbiol. 24:488-498.
25. Wu, M.,N. Davidson, and E. Wimmer. 1978. An elec-tronmicroscopic study of theproteins attached topolio virus RNA and its replicativeform(RF). Nucleic Acids Res. 5:47114723.
VOL. 39,1981