0022-538X/81/040239-10$02.00/0 Vol. 38, No. 1
Isolation and Properties of Moloney
Murine
Leukemia
Virus
Mutants: Use of a Rapid Assay for Release of
Virion
Reverse
Transcriptase
STEPHEN GOFF, PAULA TRAKTMAN, AND DAVID BALTIMORE*
Centerfor Cancer Research and Department of Biology, Massachusetts Institute of Technology,Cambridge,
Massachusetts 02139
Received 25 November 1980/Accepted 7 January 1981
A rapid assay for retroviral reverse transcriptase activity released into the
culture medium by infectedcellswas developed. With the assay,4,000 clonally infected celllines could be tested in a few hours. We have adaptedthe assayfor
use as a screen for the detection of spontaneous viral mutants. Mutants of
Moloney murine leukemiavirus have been isolated which (i) produce a thermo-labilereverse transcriptase, (ii) are temperature sensitive forrelease of enzyme activity,or (iii) can only productively infect cells already producing gag-related polypeptides.The assay has also been useful for the isolation ofnonproducercells infected withvarious replication-defective transforming viruses.
Most knowledge aboutretroviruses has been gained by direct biochemical analyses of virion nucleicacids and proteins. Very little is known about thefunction of each virus-encodedprotein in the life cycle, largely because very few
mu-tantsmappedto specific proteins and affecting
specific functions have been isolated.In the
mu-rine system, there have been
temperature-sen-sitive mutants isolated with impairedgag and
gag-pol
cleavage
and reduced virus release at390C (16, 19). Nonconditional mutants with al-tered protein phenotypes have been described
(15).Therealso have beenreportsofearly
mu-tants found to have temperature-sensitive
po-lymerase and RNase H function (17), and a nonconditionalpol mutant has been described recently (4).Finally,mutantswhichrelease par-ticles which cause no
plaque
formation in theXC assay (XC- mutants) have been described
(5, 9, 10). Few of these mutants have been mappedonthe genome orascribedto
proteins,
and the lesion in most of the
early
and latemutantsremains unknown.Moremutantshave
beendescribed and
mapped
inthe aviansystem. gagprocessing
mutants have beendescribed,
and nonconditionalmutantsingag supportthe
idea that gagproteinsarerequiredforsuccessful
virion assembly (8). The behaviorofa mutant
inp15 confinmsitsprobableroleasthe protease responsible for gag and
gag-pol
cleavage (11).
envmutantshave beendescribed
(7,
14)
whichconfirm the
following
about the envglycopro-tein: its role in
determining subgroup specificity,
its necessary presence for viral
infectivity,
anditsunimportanceinvirionassembly. Lastly, po-lymerasemutantswiththermolabileactivityare
numerous (1, 6), andmutantsinpolmaturation
have been discussed (2).
We have begunconstructingalibrary of
mu-tants of Moloney murine leukemia virus
(M-MuLV) blocked in various stages of the life cycle,which may beusefulinassigningfunctions to the known proteins. To detect mutants, a rapidassay for thevirus-encodedreverse
tran-scriptasewasdeveloped by which several thou-sand clones ofinfected cells could be
assayed
in anafternoon.MATERLA1S AND
METHODS
Celis
and viruses. The NIH/3T3 cell line wasgrowninDulbecco's modifiedEaglemedium contain-ing 10% calfserum. M23 cells were derivedin this
laboratory (15).Thesecellscontainadefective
provi-rusof M-MuLV which encodesonlythe
Pr659M
pro-tein. M-MuLV clone 1was the source of infectious viruspreparationsandwasusedas ourwild-typevirus (3). Rauscher MuLVts29 (R-ts29)was agiftfromS. Aaronson (17). Viruswastitratedbythe XC plaqueassay (13). Samples to be assayed at temperatures other than
370C
were firstpreincubatedatthe indi-cated temperature for45min in mediumcontaining 20 mM HEPES(N-2-hydroxyethylpiperazine-NV-2-ethanesulfonicacid), pH7.35, and thenappliedtothe cellsatthesametemperature.
Rapidreversetranscriptaseassay. Cloneswere
prepared by infection ofNIH/3T3 cellsat a
multiplic-ityofapproximately0.3PFU/cellfor 2 hat37°C.The
cellswerethentrypsinized,counted,and seeded into
96-wellcloning traysat 0.3cellsper well(inavolume 239
on November 10, 2019 by guest
http://jvi.asm.org/
of0.2mlperwell).The cloneswereallowedtoexpand
for10to 12days.Areplicatingdevice (Flow
Labora-tories, Inc.) wasusedtoremove10,ulof the
superna-tantmedium from eachwelland add it to 50
pi
ofareaction cocktail aliquoted into disposable 96-well trays(Dynatech Laboratories).The cocktailgavethe followingfinal concentrations: 50 mM Tris-hydrochlo-ride, pH8.3; 20mMdithiothreitol;0.6mMMnCl2;60 mM NaCl; 0.05% Nonidet P-40; 5 jig of oligodeoxy-thymidylicacidperml;10jig ofpolyriboadenylicacid
perml;and 10,uMa[nP]dTTP (specific activity,1Ci/
mmole).Thereactionwasincubatedatthe
appropri-atetemperature for 1 to 2h,and 10 pd of the mixture
wasspotted directlyontosheets ofdryDEAEpaper
(DE-81; Whatman, Inc.) ina96-spot grid, using the
mechanical replicator. The paper was then washed withgentle rockingatroomtemperature three times
in 500 ml of 2x 0.3 M NaCl-0.03 M sodium citrate for 15min each and then twice in 500 ml of95% ethanol.
Thepaper wasdried andexposedtoX-rayfilm (XR-5)at-70°Cwithanintensifyingscreen(Dupont
Light-ning Plus).
Infection of replicate trays ofcells.Virus inthe culture medium above clones of cellswaspassedonto recipient cell lawnsasfollows. The recipient96-well trayswereseeded with 0.1 ml ofacellsuspension (104
cells/ml)in mediumcontaining2,ugofPolybreneper
ml. After 1day,10lO ofthe medium in eachcloning
well ofa tray of donor cells was transferred to the recipient trays, usingthe replicating device; the
in-fectedcells were incubated for 1 day, and then the
medium was changed to normal medium (lacking Polybrene).After 1moreday,therecipientcells were
assayedforreversetranscriptase bytherapid
proce-dure.
Conventional reverse transcriptase assays.
Theconventionalreversetranscriptaseassay(12)was carried outbyincubating 50
pl
of the culturemedium to be assayedwith 100jil ofacocktail to give finalconcentrationsasgivenabovefor the rapidassayand
by countingthetrichloroaceticacid-precipitable
ma-terial after filtrationthrough membrane filters
(Mill-poreCorp.). Assaysat32and 39°Cwereperformed by
preincubationof the viruswith0.1M Tris-hydrochlo-ride, pH7.5,attheindicatedtemperatureforvarying times followed byaddition of the remaining
compo-nentsof the cocktail.
Immunoprecipitationofvirus-specific proteinsand
gel electrophoresisof theproteinswere asdescribed
previously (18).
RESULTS
A rapid screen for release of reverse
transcriptaseactivity. Theprotocolfor
assay-ingindividual clonesofcells for releasedreverse
transcriptase activity is shown in Fig. 1. Cells
wereinfected withapreparation of wild-type
M-MuLV and replated into cloning wells; after
growth, partof the culture mediumwas added
to a reverse transcriptase exogenous reaction
mixture with detergent and incubated, and a
portionwasthenspottedontoDEAEpaper.The
RAPID REVERSE TRANSCRIPTASE SCREEN
INFECT CLONE CELLS
VIRUS ~ CELL GROW UP
STRUS MONOLAYER (2-3weeks)
REPLICATE
REPLICATE TRAYOF 1oaI O(32P-TTP-RTCOCKTAIL
(hwbet.2hr. 37C) SAVE CLONES
WASH AUTORAIOGRAPH |..
DEAEPAPER
X-RAYFILM
FIG. 1. Outline of the procedure for assaying
clonedcell linesforreleaseofviralreverse transcrip-tase. Cells were infected with virus, cloned, and grownfor2 to3weeks. Ten microlitersof the culture medium was removed and mixed with an enzyme cocktail.After incubation,the[32P]DNA productwas
boundtoDEAE paper, whichwaswashed and
ex-posedtoX-rayfilm.
paperwas washed andexposedto film for
auto-radiography.
Atypicalassayon 10 traysisshown in Fig. 2.
In the experiment, approximately half of the
wells contained cells(412 of 960);approxiimately
one-third of these (125 of 412) were releasing
reverse transcriptase enzyme. Duplicate assays
of the same cells were reproducible (data not
shown;seebelow).Although the number of cells
in each well at the time of assay was highly
variable, there was no obvious correlation
be-tweenthe number ofcells and the intensity of
the signal. Many of the very dark signals were
produced by only a few cells (<103), whereas
many of the faint signals were produced by
confluent wells (containing 104 cells or more).
Weconcluded that there was a wide range in the
level of enzyme released by cell lines, as has
been notedby others (15). The sensitivity of the
assaywasveryhigh; as few as 100 cells produced
positive signals. The assay shown was completed
inafew hours of work, and the fih shown was
a 12-h exposure.
Isolation of mutants coding for thermo-labile reverse transcriptase. To detect
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.494.266.453.110.314.2]ISOLATION AND PROPERTIES OF M-MuLV MUTANTS 241
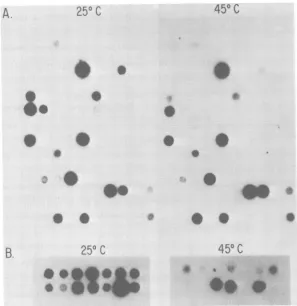
FIG. 2. A typical assay of cloned cell lines for
releasedreversetranscriptase.Ten traysofcellswere
prepared and assayed as outlined in Fig. 1. The
[32P]DNA boundtoDEAEpaperwasexposedto
X-rayfilm for12h.
tantsof M-MuLVwhich encodedathermolabile
reverse transcriptase activity, cloneswere
pre-paredin 20traysasdescribedabove and allowed
to expandfor 10 days at 370C. The cells were
thengrownfor 2moredaysat320C.Portionsof
culture medium were removed and added in
duplicate to an incomplete cocktail mix which
lacked thetemplate, primer,andnucleoside
tri-phosphate.Afterpreincubation for 1 hat45or
250C, the template, primer, and dTTP were
added,andthe reactionwasallowedtoproceed for2h. AnX-rayfilmexposureoftwopairsof
traysis showninFig.3A. Ingeneral, the clones
released temperature-resistant enzyme which
showedsimilaractivityatthetwotemperatures;
a few, however, released enzyme with a much
reduced activity at450C. Avery fewseemed to
be coldsensitive. A total of 13 clones were
se-lected (11 temperaturesensitive and 2 cold
sen-sitive), grown into larger cultures, and reassayed
as described above. Of these 13, 10 were
repro-ducibly thermolabile (Fig. 3B); their behavior
wassimilar to that of the R-ts29, as previously
described (17). No consistently cold-sensitive
clone wasdetected.
Biological propertiesofthe reverse tran-scriptase mutants. Eachofthe 10putatively
mutant clones was grown at
320C,
and eachculture medium was harvested. The released virions were preincubated for 45 min at 32 or
390C
and then titrated in XC plaque assays perfornedatthesame temperature. Of the 10,5 never gave XC plaquesunder any condition
and presumablywereeither uninfectiousdue to the mutation or were XC-negative variants of M-MuLV (5, 9, 10). Ofthe 10, 3 were XC posi-tive, but showednothermolabilityover that of the controlwild-type virus (whichlost 50 to 80%
of itsinfectivityat390C, relativeto32°C, in this
experiment). The XC titers of representative members of this group (6H3 and TSP-11F8)areshown inTable1.Theremaining two
mutants (TSP-11A4 and TSP-6E9) were
ob-viously more thermolabile than the wild type was in the XC assay; in separate experiments, they showed a 40- to 200-fold reduced titer at
thehigher temperature (Table 1). They also had
lower titers than did the wild type after
320C
incubation. Thus, these two mutants, chosen onlyonthe basisofthe invitroreverse
transcrip-tase assay, were also thermolabile by plaque
assay.
Kinetics ofinactivation ofreverse tran-scriptase. Toconfirmthequalitative resultsof therapidassay,conventionalreverse
transcrip-taseassays wereused. Virion harvestswere
col-lected from the mutant cell linesand preincu-bated for
varying
times in buffered mediumattwotemperatures, and then exogenous reverse transcription was allowed to proceed for 1 h.
Three conditionsweretested:
(i)
preincubationandassay at
250C;
(ii) preincubation andassayat
390C;
and (iii) preincubation at390C
andassayat
250C. Examples
ofinactivation curvesfor four of themutants,
along
with controlcurvesofwild-typeM-MuLV andR-ts29, areshown in
Fig.4.
Wild-type viral reverse
transcriptase
wasin-activatedapproximately50% in this
experiment
during preincubation at 25°C; itshowed some
additional loss when
preincubated
at 390C. Inother experiments,the loss of
activity
waslessbut was
generally
10 to 50% and wasslightly
VOL. 38, 1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.494.48.241.63.453.2]242 GOFF, TRAKTMAN, AND BALTIMORE
FIG. 3. IsolationofceU clonescontainingmutantM-MuLV genomesencodingthermolabilereverse tran-scriptase. (A)Ten traysofclonedcellswereassayedintheduplicateat25and450C.
Autoradiograms of
twoof these traysareshown. A totalof13clonesshowingdifferentialactivitywerechosen
from
thisscreen.(B)
Repeat ofthe assay on the 13 clones chosen from the initial screen. Ofthe 13, 10 were
reproducibly
thermolabile.R-ts29 isincludedfor comparison (lowerright sample).
TABLE 1. XCplaqueassayonthermolabilereversetranscriptasemutants'
Expt 1 Expt2
Virus Temp('C) Ratio(32'C/ Temp (°C) Ratio (32'C/
32 39 39°C) 32 39
390C)
Wildtype 3x105 5x104 6 5x 104 2x 104 2.5
TSP-6H3 5x10' 2x101 2.5 5x10' 2x10' 2.5
TSP-11F8 9xi05 2x105 4.5
TSP-6E9 5 x 103 <5x101 100 4 x103 2 x10' 200
TSP-11A4 2 x 103 5x10' 40 2x 103 2 x10' 100
aVirus harvestswerecollectedat32°C. The viruswaspreincubatedattheindicatedtemperaturefor45min
and then titratedatthesametemperature.Tableentriesaretiters inPFU/ml.
greater during preincubation at the elevated
temperature. The activity, when assayed at
390C,
wasconsiderably reduced,independent of the time ofpreincubation. R-ts29 activity wasstable at
250C,
butextremely sensitive toprein-cubation at
390;
1hreducedthe activity 20-foldtoundetectable levelsevenwhenthe assay was performedat25°C. Essentiallynoactivity could
bedetected in any assay carried out at
390C.
The reversetranscriptase released by each of
the mutant clones was morethermolabile than
the wild-type enzyme. The enzymes produced
bytwoof the clones(TSP-11A4andTSP-6E9)
were extremely temperature sensitive; like R-ts29,
they
were rapidly inactivated bypreincu-bationat39°C (Fig. 4). Theyalsoshowed almost
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.494.103.400.79.385.2] [image:4.494.54.453.453.548.2]ISOLATION AND PROPERTIES 243
WILD TYPE
100 100
50 5 3 450
*1
00 15 30 50
TSP 6E9
00- _ 100
0
\\ o --,
50_ .s 50
0 15 30 50
R-TS21
TSP 6H
.A
9 the virions released in that period were har-vested andbanded
by isopycnic centrifugation.
.~--_ Whenthe labeled proteins in the virions were
>0
analyzed
bypolyacrylamide
gel electrophoresis(Fig.
5),
cellsinfected with wild-type virus werefound to have released large amounts of virion
proteins
atbothtemperatures; the most abun--- dant protein byfarwasp30,a productof the gag50 gene. Neither clone 6E9 nor clone
TSP-13 6H3 was temperature sensitive for release of
° virions (data not shown), although both
pro-duced athermolabilereversetranscriptase
activ-ityand one, inaddition,wastemperature
sensi-tive for plaque formation (Table 2). Thus, re-verse transcriptase mutants are not always af-...._. fected in release. Two other clones, however, f
(TSP-11A4
andTSP-11F8)
were temperatureon
1 30 50
TSP IIA4 TSPIIF8 o o
100__100 o o
50 50
0 15 30 50 0 15 30 50
Preincubation (minutes)
FIG. 4. Heat-inactivation curvesforreverse tran-scriptasereleasedfrom wild-typeM-MuLV-infected cells, R-ts29-infected cells, andfourmutant clones. Culture medium fromeach clone waspreincubated
for varyingtimes and thenassayed forafixedtime
(1 h). The conditions were: bothpreincubationand assayat390C(-); preincubationat39°Candassay
at25°C (A); and bothpreincubation andassayat
25°C (0).Eachofthefourclones releasedareverse
transcriptaseactivitymorethermolabile than thatof
the wildtype.
noactivity when assayed at 390C. Two others
wereless sensitive topreincubation,with
activ-ity fallingslowly during preincubationandassay
at 390C. Clone TSP-11F8 produced enzyme
which was particularly resistant to a 30-min
preincubation at 390C if the assay was then
performedat250C. Thus,theenzymefromthis
clone was not irreversibly inactivated by the
preincubation. It is noteworthy that the two
viruses containing the most thernolabile
en-zymes were the same twowhich showed
tem-peraturesensitivityinthe XCplaqueassay
(Ta-ble2).
Associated phenotypic changes in
tem-perature-sensitive reverse transcriptase
mutants. Four of the mutant cell lines were
testedfor additional effects of themutationon
theassemblyofvirions. Cellswerelabeled
[image:5.494.54.243.57.355.2]over-night (at32or390C) with[35S]methionine,and
TABLE 2. Summary ofproperties of four thermolabile reverse transcriptase mutantsa
Raidrse
Rapidre-~transcrip-
Reverse
Vhion
VrsVr
transcrip-
verse XCXCa8yassay taset8 heatet rlrelease taseassayinactiva-TSP-6H3 tsb +(very low) ts +
TSP-11F8 ts + ts ts
TSP-6E9 es ts ts +
TSP-11A4 Ls ts ts ts
aData weretaken from Table 1 and Fig. 3 through 5. ts,Temperature sensitive.
320 390 320 390 320 390 320 390
FIG. 5. Analysis ofvirionproteins released into the culturemediumbyclonesexpressingthermolabile
reversetranscriptase. Cells were labeled with
["SI-methionineat 32or390C, and the virions released into the medium werepurified. Theproteins were
separated by electrophoresis through polyacrylamide gels and detected byfluorography. Theposition of
the viralprotein p30 is indicated.
CI-w
0
0
38, 1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.494.257.441.277.582.2] [image:5.494.278.421.401.580.2]TRAKTMAN,
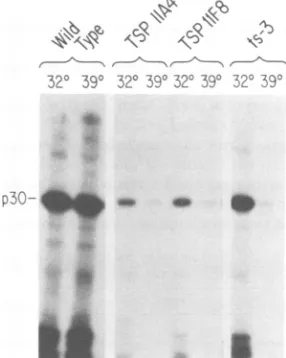
sensitive for release of virus (Fig. 5); much less
p30 wasdetected in harvests atthe high
tem-perature. Their behavior was similar to that of
M-MuLV ts3 (Fig.5). In asimilarway, the
R-ts29-infected line was temperature sensitive for release (data not shown). Thus, a number of independentreversetranscriptasemutants have
beenfound tobe,inaddition, defectivein their
abilitytoassemble orreleasevirions.
Isolation ofmutants temperature sensi-tive for release of reverse transcriptase. The reverse transcriptase assay could also be
used todetectandisolatemutations in retroviral
proteins other than the reverse transcriptase
itself. To detectclones with conditional
muta-tionsin any gene affecting assemblyorrelease
ofvirus, we chose to assaythereverse
transcrip-tasereleasedateach of two temperatures.Cells
were infected as above and distributed to 10
traysofcloningwells.Thecloneswere expanded
for20daysat320C, and the culture mediumwas assayedat250C (withoutapreincubation); then the mediumover the clones was changed, and after 2 daysat 390C, a second portion was as-sayed at 39°C. Comparison of the two assays revealed approximately 11 clones (out of 200
producing reversetranscriptase)whichreleased
less activityatthe elevatedtemperature; seven
of these provedtobeconsistently temperature
sensitive for release upon retesting. No
con-sistentlycold-sensitive clones weredetected. Preliminary characterization ofmutants
temperaturesensitive forrelease. The
viri-ons of seven clones which were temperature
sensitive for release were tested biologically. Culture mediumwasharvested fromeach ofthe
sevenclones after incubation for 1dayat
320C.
These harvests were then titrated in an XC
plaque assay carried out at either 32 or
390C.
Wild-type virus showed a high titer at either
temperature (Table 3). Four of the mutant
clones either released viruswhichwas XC
neg-ative in the assay ateither temperature or
re-leased a verylow titerof viruswhich was
unde-TABLE 3. XCplaque assay on temperature-sensitive virus harvested at32°C
Virus
Tempa
(°C)
Ratio(320C/
32 39
Wild type 4 x105 2 x105 2 TS1OC7 5 x 104 <103 >50
TS 1E6 8x 104 3 x103 27
TS4D2 1.4x 104 2x103 7
a
Virus
harvestswerecollected
fromcell
lines after1dayofgrowthat320C.Thevirus waspreincubated attheindicated temperaturefor45minandtitrated
atthesame temperature. Table entries aretiters in
PFU/ml.
tectable inourassays.Theremaining three
mu-tantclones released virions whichwerecapable
of producing plaques at 320C but which
pro-ducednoorfewer plaques when assayedat390C
(Table 3). Thus, the three producer cell lines carrying these viruses were temperature
sensi-tive for release of virion reverse transcriptase,
and, moreover, the virus released at the
permis-sive temperature was itselfunabletocomplete
aninfectionatthe nonpermissive temperature.
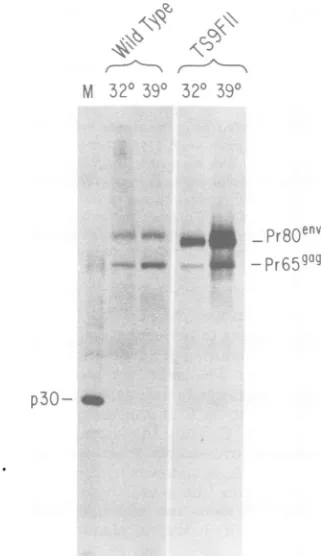
The viral proteins synthesized by the seven
cell lines were analyzed by metabolic labeling
with[3S]methionine, immunoprecipitation,and
polyacrylamide gel electrophoresis. The
prod-uctsof thegag gene
(Pr65`)
and the envgene(Pr80env) weredetected in all of the clones; the
onlyabnormalitywas alarge overproduction of
Pr8Oenv at the elevated temperature by one of
the XC-negative clones (TS 9F11) (Fig. 6). No
other gross changes in the size orlevel of
pro-duction of the viral proteins were detected
among themutants.
Isolation of mutants in thegag gene.
Us-M 320 390 320 390
I-Pr8Oenv
-Pr659°9
p30-FIG. 6. Envelopeglycoprotein produced by the TS
9F1 Iclone. The indicatedcells were labeled at 32 or
39°C with [35S]methionine, andextracts were
pre-pared. Viral envelope glycoprotein was immunopre-cipitated. The proteinswereanalyzed by
electropho-resis andfluorography.
VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.494.265.426.319.597.2]ing therapid assay,clonalisolates of M-MuLV canbetested for theirabilitytoreplicate intwo different host cells. Ifone of the hosts already
carries adefective provirus which provides one
oftheviral functions, thenmutants inthat func-tion wouldbe detected ashostrange mutants,
able to grow onthe hostcarrying the function
and not onnormal cells. Hosts expressing the
env function could not be used to isolate env
mutants because they are not superinfectible,
but hosts producinggag orpolproteins could, in principle, be used toisolate mutants in the gagorpolgene.One such hostcell wasavailable
to us in the form of the cell line M23 (15), a
derivativeof NIH/3T3 cells carryingadefective M-MuLV
provirus
and synthesizing only Pr659"9. Preliminary mapping (unpublished data)suggeststhat theprovirusinM23 contains a1.4-kb deletionmappingintheenvgene,near the 3' endof the viralgenome.Thiscell line hasbeen found tosynthesizeashortviralRNA(15)
which
apparently
directs synthesis of onlyPr65V"';
nopol- orenv-relatedprotein has been detected. Because this host cell is superinfecti-ble, inprinciple
itwould bepossible to isolategag mutantswith thishost, which wouldgrow
by complementation but which wouldnot rep-licateinnormal cells.
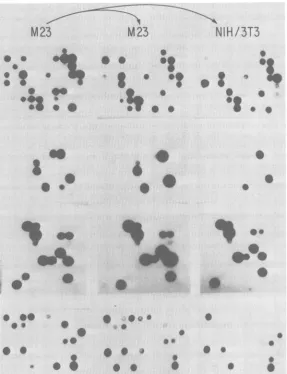
Accordingly, M23 cells were infected with wild-typeM-MuLV anddistributedto 10
cloning
trays asdescribed above. After15days at
370C,
the cells were assayed for release of reverse
transcriptase;
at the same time, culture fluids werepassedtofreshM23cellsand in duplicate tofreshNIH/3T3cells.After 2days, thesecells
were also
assayed.
Two clones were detected which releasedtranscriptase activity
and re-leased virus whichproductively
infected M23 cells but whoseyield
did not infectNIH/3T3
cells
(Fig. 7).
Analysis ofthe viralproteins synthesized by thetwoclones showednoalterations from wild-typecontrols (datanotshown). Analysis of the
virusreleased from theseclones by XC plaque
assay,however, showedadistinct
difference
be-tweenthe clones and controls andconfirmed the
hostrangephenotype of thesetwomutants
(Ta-ble 4). The supernatants were titrated on two hosts: M23 cells and NIH/3T3 cells. Wild-type virus, as well as wild type virus grown on M23
cells,
showed identical titers on these two hosts.The virus releasedbythe two clones showed a
hightiteronM23andavery low titeron
NIH/
3T3 cells (Table 4). In addition, the mutant
plaque numberson M23cells gave a linear
re-sponsetodilution;their
plaque
numberson3T3cells, however, dropped off very rapidly with
dilution of the stock, suggesting that multiple
hits were required to initiate an XC plaque. The data were consistent with the notion that two genomes were present in the virus preparations
and that both must infectthe same cell to
pro-duce a plaque. The virus would yielda higher
andlinear titeron M23 cells because thesecells alreadycontainedone of the required viruses as anendogenousprovirus.
Isolation
of transformed nonproducer cell lines.A commonprobleminthe analysisofreplication-defective
transforming viruses is theisolationofatransformedcell clone which is free
of helper virus.Wemadeuseof the rapid reverse transcriptaseassay to facilitatethis procedure. Amixed stockof AbelsonMuLVwith its helper M-MuLV was collected from a producer cell. NIH/3T3
cells
wereinfected at a lowmultiplic-ity (0.7 PFU/cell; 0.15 focus-forming units per cell),trypsinized, and clonedinmicrotiter trays. Aftergrowth for13days,the clones were assayed forreleasedreverse transcriptaseactivity.Those clones scored as negative for release ofreverse transcriptasewerethen scored
visually
for mor-phologicaltransformation.
In thisway, several transformed nonproducerlines
wererapidly iso-lated. In one representative experiment, 4% of thetotal clones showed the desired phenotype.DISCUSSION
The reverse transcriptase assay described is
rapid andinexpensive; several thousand clones can be screened in a
day
by
one person. Theassayis
remarkably sensitive,
allowing
detectionofactivityfrom asfewas a 100cells.The screen isdirectly useful forcloningvirusesandfor the isolation of
transformed nonproducer
clones fromamixed virus stock ofatransforming virus
and its
helper.
Inaddition,
the screen isex-tremely
helpful
in theidentification of variant clonesproducingtemperature-sensitive
or hostrange mutantsof retroviruses.Wehaveisolated
asmall number ofmutantsof threetypes;more
of each class andof other classescould
easily
be isolatedfor furtheranalysis.
Aremarkablefinding
implicit
in oursuccessful isolation ofthesemutantsisthehigh
frequency
of spontaneous mutations which arise in puta-tively
wild-type
stocksofM-MuLV.Thermola-bile and
temperature-sensitive
mutants werefound in screens ofonly200 to 300
clones;
thefrequency of recovery of mutants was in the
range of 3 to 5%. Thishigh
frequency
ofappear-ance of mutations has been noted
previously
(15). Some step in the life
cycle
ofM-MuLV,
and
presumably
many otherretroviruses,
must be highlymutagenic;
likely
candidates for this step are thetranscription
of theprovirus
intoon November 10, 2019 by guest
http://jvi.asm.org/
246 GOFF, TRAKTMAN, AND BALTIMORE
FIG. 7. Isolationofhost rangemutants.M23 cellswereinfectedwithM-MuLV, cloned, andassayedfor reversetranscriptase(left panels). The virusreleased by these cellswasthen passedtofresh M23 cellsor to
freshNIH/3T3 cells. After2days, these cells were also assayed (middle andrightpanels). Clonesreleasing
viruscapable ofinfecting M23 cells but notNIH/3T3 cells were selected for further study.
thegenomic RNAand thereversetranscription of the RNA into circular double-stranded DNA.
Bothof these stepsarethoughttobe carriedout
by nonproofreading polymerases and maywell havehigherror rates.Theremaythus be several
mutations introduced into each retrovirus
ge-nomeeach time it ispassedfrom celltocell.
The mutantsisolated in ourfirst screen are probablydefective in the pol gene itself because
the enzyme is thermolabile in vitro by two
as-says. It isnevertheless possible that mutations
inadjacent viral proteins could reduce the
sta-bilityofthe enzyme. It isinterestingthatthepol
mutantsoften,butnotobligatorily,havedefects
in assembly and release of virions. These
mu-tants raise the
possibility
thatthepolymerase
proteinor aprecursorof the enzyme is involved
in virion assemblyatthe cell surfaceasis also suggested by other data (P. Traktman and D. Baltimore,manuscript inpreparation).
The mutants recovered in thesecondscreen
could potentially lie in any viral gene, or even
potentiallyinacellular gene, neededfor
assem-bly; the high frequencyof appearance of these
mutations would argue for a viral origin, and
those that have a temperature-sensitive yield
mustbe in the virus. Theonly gross alteration
inamajorviralproteinwastheoverproduction
of theglycoproteinPr8Oenvatthenonpermissive
temperature inone clone. All of the other
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.494.110.397.77.451.2]ISOLATION AND PROPERTIES OF M-MuLV MUTANTS 247
TABLE 4. XCplaque assay onvirus preparations harvested from clonedcell linesa
Celllawn Virus prepn Dilution
NIH/3T3 M23
Wildtype grownonM23 cells 1:100 TMTCb TMTC TMTC TMTC
1:300 500 500 500 500
1:1,000 200 200 200 200
1:3,000 100 100 100 100
1:10,000 75 47 37 45
1:30,000 22 16 12 15
1:100,000 6 6 4 3
Clone 8G4 1:100 100 200 TMTC TMTC
1:300 23 13 500 500
1:1,000 2 1 150 150
1:3,000 2 2 64
-1:10,000 0 0 30
1:30,000 0 0 6
1:100,000 0 0 0 1
Clone M16 1:100 100 100 TMTC TMTC
1:300 13 17 300 300
1:1,000 100
-1:3,000 0 0 43 40
1:10,000 0 0 14 12
1:30,000 0 0 4
-1:100,000 0 0 0 1
Wild-type virus grown on 1:1,000 105 57
NIH/3T3cells 1:10,000 13 4
1:100,000 1 0
1:1,000,000 0 0
a
Virus
harvests fromthe indicatedcell
lines were diluted and used to infect either NIH/3T3cells
orM23 cells. Table entriesareplaquescounted per6-cmdish.bTMTC, Too manytocount.
tants presumably either carry point mutations
which inactivate the function of a particular
protein without altering its size or alter
unde-tected minorproteins.
Mutations of our thirdclass,which are
com-plementedbythe defectiveprovirusof M23cells,
presumably lie in the gag gene. In M23 cells
these mutants replicatenormally, but inNIH/
3T3 cellsthey donot.Apparentlyrecombination
did notoccur in the initial infection becausein
preliminary experiments two RNA sizes were
still detectable in the cells: full-size RNA and
the deleted M23 RNA (unpublished
data).
Sim-ilarly,among theraresuccessfullyinfected
NIH/
3T3 cells therewere some reverse
transcriptase-positive clones which also carried both RNAs.
Thus bothgenomescanbereleased and
appar-ently must be transmitted to a newly infected
cell if that cell istobecomeaproducerof virus.
The virus thus showsahightiteronM23cells,
which already provides one of the needed
ge-nomes, and avery low titer on
NIH/3T3
cells.The fact that the mixed virus stock shows
mul-tihit kinetics on NIH/3T3 cells suggests that
both genomes
rarely
arepackaged
intoonevir-ion or, morelikely, that both genomes in a virion
arerarely convertedtofunctionalproviruses.
Weareattemptingtoclonethesemutantsfree
of their M23helper byscreening NIH/3T3cells
infectedat alowmultiplicityforthesynthesis of
viralproteins. We can thenanalyze themutants
biochemicallyand determine the exact nature of
the defect in thegag gene.
ACKNOWLEDGMENIS We thank Owen Witte forhelpfuldiscussions.
ThisworkwassupportedbycontractN01-CP-53562 from theDivision of Cancer CauseandPrevention,National Cancer Institute, and by Public Health Service grant CA-14051(core granttoS.E.Luria)from the NationalCancer Institute. S.P.G. isa Jane CoffinChilds Memorial Fund Fellow. D.B. isan
AmericanCancerSocietyResearch Professor. LITERATURE CITED
1. Alevy, M. C. andP. K.Vogt.1978.Tspolmutantsof
avian sarcomaviruses:mappingand demonstration of single-cyclerecombinants.Virology87:21-33. 2. Eisenman,R.N.,W. E.Mason,and M.Linial. 1980.
Synthesis and processing ofpolymerase proteins of wild-typeandmutantavianretroviruses. J.Virol. 36: 62-78.
3. Fan,IL, andM. Paskind. 1974. Measurement ofthe sequencecomplexityofclones ofMoloneymurine leu-VOL. 38, 1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.494.50.448.87.375.2]kemia virus 60 to 70S RNA: evidence for a haploid genome. J. Virol. 14:421-429.
4. Gerwin, B. J., A. Rein, J. G. Levin, R. H. Bassin, M. B.Benjers, S. V. S. Kashmini, D. Hopkins, and B. J.O'Neill. 1979. Mutant of B-tropic murine leukemia synthesizing an altered polymerasemolecule. J. Virol. 31:741-751.
5. Hopkins, N., and P. Jolicoeur. 1975. Variants of N-tropic leukemia virus derived fromBALB/c mice. J. Virol. 16:991-999.
6. Linial, M., and W. S. Mason. 1973. Characterization of twoconditionalearly mutants of Rous sarcoma virus. Virology 53:258-273.
7. Mason, W., and C. Yeater. 1977. A mutant of Rous sarcomaviruswith a conditional defect in the determi-nant(s) of viral host range.Virology 77:443-456. 8. Mason,W.S.,C.Yeater, T. V.Bosch, J. A. Wyke,
and R. R. Friis. 1979. Fourteentemperature-sensitive replication mustants of Rous sarcoma virus. Virology 99:226-240.
9. Rapp, U. R., and R. C. Nowinski. 1976. Endogenous ecotropic mouse type C virusesdeficient in replication andproduction of XC plaques. J. Virol.18:411-417.
10.Rein, A.,B.I.Gerwin,R. H.Bassin, L. Schwaim, and G. S.Schialovsky. 1975. Areplication-defective var-iant of Moloney murine leukemiavirus. I. Biological
characterization.J.Virol.25:146-156.
11.Rohrschneider,J.M., H.Diggelman, H. Ogura, R. R. Friis, and H. Bauer. 1976. Defective cleavage of a precursor polypeptide in atemperature-sensitive
mu-tant of aviansarcoma virus.Virology 75:177-187. 12. Rosenberg, N., and D. Baltimore. 1978.Theeffect of
helper virus on Abelsonvirus-induced transformation oflymphoid cells. J. Exp. Med.147:1126-1141.
13. Rowe, W.P.,W.E.Pugh,and J.Hartley.1970.Plaque assaytechniques for murine leukemia viruses.Virology 42:1136-1139.
14. Scheele, C. M., and H. Hanafusa. 1971. Proteins of helper-dependent RSV. Virology 45:401-410. 15. Shields,A.,0. N.Witte,E.Rothenberg,and D.
Bal-timore. 1978.High frequency of aberrantexpression of Moloney murine leukemia virus in clonal infections. Cell 14:601-609.
16.Stephenson,J.R., and S. A. Aaronson.1973. Charac-terizationoftemperature-sensitive mutantsof murine leukemia virus.Virology 54:53-59.
17.Tronick,S.R.,J.R.Stephenson,I.M.Verma, and S. A. Aaronson. 1975.Thermolabile reverse transcriptase of a mammalian leukemia virus mutant temperature sensitive in its replication and sarcoma virus helper functions. J. Virol.16:1476-1482.
18.Witte,0.N.,N.Rosenberg,M.Paskind,A.Shields, and D. Baltimore.1978.Identification ofanAbelson murine leukemia virus-encoded protein present in transformed fibroblast and lymphoidcells.Proc. Natl. Acad. Sci.U.S.A. 75:2488-2492.
19.Wong,P. K.Y.,L. J.Russ,and J. A.McCarter. 1973. Rapid selecti e procedure forisolation of spontaneous temperature-:?nsitive mutants ofMoloney leukemia virus.Virology 51:424-431.

![FIG. 2.prepared[32P]DNAreleasedray A typical assay of cloned cell lines for reverse transcriptase](https://thumb-us.123doks.com/thumbv2/123dok_us/1482123.100847/3.494.48.241.63.453/prepared-dnareleasedray-typical-assay-cloned-lines-reverse-transcriptase.webp)