0022-538X/92/052740-08$02.00/0
Copyright ©3 1992, AmericanSocietyfor Microbiology
Structural and Functional
Characterization
of the
Poliovirus
Replication Complex
KURTBIENZ,* DENISE EGGER, THOMAS PFISTER, AND MONICA TROXLER InstituteforMedical Microbiology, UniversityofBasel, Petersplatz 10, CH-4003Basel, Switzerland
Received 18November1991/Accepted 10 February 1992
Two populations of membrane-bound replication complexes were isolated from poliovirus-infected HEp-2
cells bysucrosegradient centrifugation. Thetwofractions show similar ultrastructural features: the replication complex is enclosed ina rosettelike shell of virus-induced vesicles and contains a verytightlypacked second
membrane system (compact membranes). The vesicular fraction, which bands in 30% sucrose, contains
replicative intermediate (RI) and 36S RNA. The fraction banding in 45%sucrosecontains only minuteamounts
of RI and contains mainly 36S RNA, two-thirds of which is encapsidated. In vitro, the two fractions show similarRNAsynthesizing capacities and produce 36Splus-strandRNA. Dissolving themembranes within and around synthetically active replication complexes with sodium deoxycholate abolishes the completion of36S RNAbutstill allows elongationinthe RI. Ourfindingssuggestanarchitecture of thereplication complex that has thenascentplus strandsonthe RIenclosed in thecompactmembranes and the replication forks wrapped additionally inprotein. Plus-strand RNAcanbe localizedbyinsituhybridization withabiotinylatedriboprobe between thereplication complex and therosetteof the virus-induced vesicles. Itwas found that theprogeny RNAstrands aresetfreesoon aftercompletion from the replication complex at the sites where thecompact membraneswithin thereplication complexare inclosecontactwith thesurrounding virus-induced vesicles.
The replication of poliovirus RNA is carried out by the primer-dependent viral polymerase 3DPo' and proceeds asymmetrically in two distinct steps. First, the input plus-strand(genomic) RNA iscopied into aminusstrand, which leads to the formation of the double-stranded replicative form(RF) (1, 24). This reaction canbe studied invitro ina reconstituted system (28) and does not depend on any
cellular component or structure. The next step, however, whichproducesprogenyRNA strands ofpositive polarity, is fully dependent on specialized cellular membranous struc-tures(25). Itstartsfromthe RF andproceedsin thepartially double-stranded replicative intermediate (RI) (11), which is confined in the viral replication complex. The replication complexwasidentifiedby electron microscopy(EM) (3) and foundtobesurrounded by and stretchedoutinsidearosette
of virus-induced cytoplasmic vesicles (6). The structural configuration of the replication complex is essential for ongoingplus-strand RNA synthesis and is maintained by the membranes of the virus-induced vesicles in concert with viral protein 2C (6).
The strict dependence of the functional replication com-plex on cellular membranous structures has hitherto pre-cluded the use of truly reconstituted in vitro transcription systemstostudy viralplus-strandRNAsynthesis. By using appropriate subcellular fractions of infected cells, however, 36Sgenomic RNAcan be produced in vitro (6, 10, 11, 24, 25), and thus at least some of the requirements for this synthetic activitycan be investigated.
In thepresentstudy,weusedsuchsubcellular fractionsto
elucidatetherolesof differentcomponentsof thereplication complex and the surrounding vesicles in viral plus-strand RNA synthesis. By EM immunocytochemistry and in situ hybridization, we visualized the spatial relation of viral macromolecules involved in RNAsynthesis withrespectto
each otherandtothecellular (yet virus-induced)structures.
* Correspondingauthor.
This allowed us to draw conclusions about the functional interaction of the replication complex with its associated membranes. In addition to the known virus-induced vesi-cles, we could identify a second membrane system within the replication complex. This membrane system is very tightly packed and appearsto interact withthe surrounding virus-induced vesicles in the last stepsofcompletion of the
36S progeny RNA. We also found that elongation of
plus-strand RNA at the RI proceeds in the absence of mem-branes, whereas completion and liberation of36S RNAare dependenton membranes.
MATERIALS AND METHODS
Cells and virus and in vivo labeling ofviral RNA. HEp-2 cells and poliovirus type 1 (Mahoney) were grown in sus-pension cultures. Themultiplicity ofinfection for the exper-iments was 30 PFU per cell. To label viral RNA, 5 p.gof actinomycin D (Merck Sharp & Dohme, Rahway, N.J.)per mlwas addedtoinfected cells 30min before the addition of 50 p.Ci of [5-3H]uridine (Amersham, Amersham, United Kingdom) perml. The labelwas added at 2.75 h postinfec-tion and left until the cellswere harvested at 4 h postinfec-tion.
Isolation and in vitro RNA synthesis of replication
com-plexes attached to virus-induced vesicles. Preparation of cy-toplasmic extracts by Dounce homogenization and low-speed centrifugation and isolation of the vesicle-attached replication complexes were described previously (6). In short, cytoplasmic extracts were centrifuged onto a double
sucrose cushion (30% sucrose layered over45% sucrose in
reticulocyte standard buffer) for 1 h at 55,000 rpm in an
SW55 rotor. Thevesicular fractions synthesizedviral RNA whenintroduced into acell-free transcription system (6, 24) containing all four nucleotides and an ATP-generating
sys-tem. To monitor in vitro RNA synthesis, 50 p.Ci of [5,6-3H]UTP (Amersham) per40 p.l was added tothe transcrip-tion system.
2740
on November 9, 2019 by guest
http://jvi.asm.org/
In some experiments,membranes weredissolved by treat-ing the vesicular fraction with 0.5% sodium deoxycholate
(DOC; Difco) for 30 minon ice (5). To test the accessibility of viral RNA, the nativevesicular fraction was treatedwith 100 ,ugof bovine pancreas RNase A(Boehringer Mannheim) perml for 15 min at25°C.
Isolation and characterization of viral RNA. In vivo- or in vitro-synthesized viral RNA was isolated by the
phenol-chloroform-isoamyl alcohol method (23), precipitated with ethanol, redissolved in water, adjusted to 10 mM Tris-HCl (pH
7.4)-i
mM EDTA-2 mM dithiothreitol-1 U of RNase inhibitor from human placenta (Boehringer Mannheim) perp.l,
and loaded on 5 to 20% sucrose gradients in 10 mM Tris-HCI (pH7.4)-i
mM EDTA. Thegradients were centri-fuged for 2.5 h at50,000 rpm in an SW55 rotorand fraction-ated from the bottom, and theradioactivitywasmeasured in aliquid scintillationcounter.Sedimentationcoefficient stan-dardsincluded3H-labeled poliovirus36S RNAisolated frompurified virions (7)and Escherichia coli 16S and 23S rRNA obtained from Boehringer Mannheim.
Biotinylated RNA probes for in situ hybridization. For in situ hybridization on the negatively stained preparationsof vesicles and replication complexes, the RNAprobe utilized for sectioned material was used (26). In short, from a plasmid (pVR106 [22]) containing full-length poliovirus
cDNA, nucleotides 670 to 2099 were excised with BamHI,
cloned intoapGEM-3vector(PromegaBiotec)between the SP6 and T7 promoters, and linearized with SspI at
position
1639 of the insert. The sequence between nucleotides 1639 and 2099 was transcribed in vitro into RNA of negative polarity with 50 U of T7 RNApolymerase (BRL)and 1 mM biotin-11-UTP (Enzo).
MAb. Monoclonal antibodies
(MAb)
against proteins
of the P2 genomic region ofpoliovirus
were characterized previously (3, 19). Theyrecognize an epitope in protein 2C and its precursors 2BC and P2. They will be referred to hereafter as 2C-MAb. Thehybridoma
culture supernatants wereused at a 1:2 dilution forimmunocytochemistry.
EM. Specimens for EM histochemistry, immunocyto-chemistry(IEM), and in situ
hybridization
wereprepared
as described previously (6) by adsorbing the vesicle fractions onto nitrocellulose(Parlodion)-coated
EMgrids.
The prep-arations were negatively stained with 1%phosphotungstic
acid, pH 7.0, before being viewed in a Siemens 102 EM operated at 100or 125 kV.
(i)Detection of viralP2 proteins byIEM. For
immunocyto-chemical labeling,
nonspecific binding
siteson the prepara-tions were blocked with normal goat serum and bovine serum albumin. Next, thegrids
were incubated with 2C-MAb and then with goat anti-mouse antibodiescoupled
to 5-nm colloidal gold (JanssenPharmaceutica)
as describedpreviously (6).
(ii) Detection ofRNAwithRNase-gold.For RNAdetection withRNase-gold(2),the same types of
specimen
and block-ing reaction utilized for IEMwere used. To visualizeRNA,
gridswere incubatedon RNase
coupled
to 15-nmgold
(E-Y
Laboratories, SanMateo,Calif.).Carewastakento
keep
all solutions as RNase free aspossible.
(iii) Doublelabelingfor RNAand P2 proteindetection. The RNase-goldand the 2C-MAbwere mixedand
applied
tothepreparation. Afterbeingwashed, the
grid
was treated with 5-nm gold-labeled anti-mouseantibody
andnegatively
stained asdescribedabove.
(iv) EM in situ hybridization. The vesicle fractions de-scribed abovewerealsoused for in situ
hybridization,
which was performed essentially as describedpreviously
forul-dpm 45% ves. 40000 20000 dpm 30% ves. *20000 *10000 30 fractions
FIG. 1. RNAcontent of vesicular (ves.)fractions isolated from poliovirus-infected HEp-2 cells. The cells were labeled with
[3H]
uridine from 2.75 to 4 h postinfection, and RNA ofthe vesicular fractions was analyzed, afterphenol extraction, by centrifugationthrough5to20% sucrosegradients.The30% vesicularfraction(A) contains 36S RNA and RI sedimenting at 27 to 30S. The 45% vesicular fraction (A) contains 36S RNA and a small shoulder (fractions20 to22)ofpresumably RI RNA.
trathin sections
(26).
Some modificationsproved
to bees-sential to retain ultrastructural
preservation
of thenative,
and thus rather
fragile,
vesicles. After in vitrotranscription,
thevesicular fractions werewashed onceby
centrifugation
andresuspended
inphosphate-buffered
saline(PBS).
UVcross-linking
minimized the loss of RNA in thesubsequent
steps. The material was then adsorbed to EM
grids,
andhybridization
wasperformed
for 2 h at58°C by
floating
thegrids
on10-,ul drops
of ahybridization
solutioncontaining
4x SSC
(lx
SSC is 0.15 M NaClplus
0.015 M sodiumcitrate),
13.5%formamide,
and thebiotinylated riboprobe.
Afterthe
grids
werewashed withPBS,
biotin detectionwasperformed
immunocytochemically
with anti-biotin antibod-iescoupled
to 6-nmgold
(AURION, Wageningen,
TheNetherlands).
As anegative control,
an unrelatedbiotiny-lated RNA
probe
(vaccinia
virusthymidine kinase)
wasused.
RESULTS
Content ofviral RNA
species
intwovesiclepopulations.
Thepoliovirus
replication complex,
attached tovesicularmem-branes,
isfound intwodistinctpopulations
of vesicles(6);
it bands in 30 or 45% sucrose and is referred to asthe 30 or45%
fraction,
respectively.
Weinvestigated
possible
differ-encesincontentof viral RNAby
phenol
extraction followedby
sucrosegradient
centrifugation
of in vivo [3H]uridine-labeled RNA.Figure
1shows that the30% fraction contains 36Sgenomic
RNAinadditiontoRIsedimenting
at27to30S. The 45% fractions containedlargely
36Sgenomic
RNAand asmallamountofmoreslowly
sedimenting
RNA ofpresum-ably
RIconfiguration.
RNase treatmentafter
phenol
extraction indicated thatin both vesiclepreparations,
no double-stranded RNA wasdetectable
(not
shown).
If the vesiclepreparations
weretreatedwith RNase Abefore
phenol
extraction(Fig. 2),
the 36SRNA inthe30% fractionwasfoundtobefully
accessible to the enzyme, whereas the RI wasprotected.
In the 45%fraction,
abouttwo-thirds of the 36S RNAwasfoundtobeprotected
against
RNase(compare
thecorresponding
36Son November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.315.552.77.235.2]dpm
36 - 23 -16S 40000
-30000
20000
-10000
0-0 1 0 20 30
fractions FIG. 2. RNase treatment ofa30%vesicular fraction(A)digests practically all of the 36S RNA but leaves the RI intact. RNase treatmentofa45%vesicular fraction (A)digestsabout one-third of the 36S RNA, leaving the remainder intact because it is encapsi-dated (data not shown). RNA was analyzed as described in the legend toFig. 1.
peaks in Fig. 1 and 2). The protected RNA of the native, non-phenol-extracted45%fractionsedimentedat
150S
(datanot shown) and thus wasencapsidated invirions.
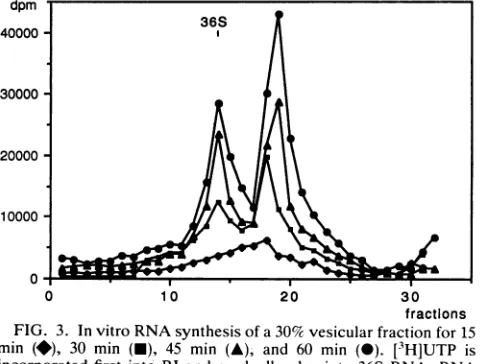
Transcriptional activity of the 30and45%fractionsinvitro. The amount and type of RNA synthesized in vitro were monitored for both vesicular preparations. The 30% frac-tions incorporated [3H]UTP at first into RI and gradually
also into 36S RNA (Fig. 3). The 45% fractions, which contained predominantly 36S RNA, showed kinetics of RI and36S RNAsynthesis in vitro (Fig. 4) similar to those of the30% fraction. Afterapproximately60min, both systems became exhausted. The invitro-synthesized36S RNA of the 30% fractionaswellasthatof the45%fractionwasfoundto befullysensitivetoRNase,whereas theRI ofboth fractions wasprotected (not shown). Thus, during invitro
transcrip-tion, both fractions react to RNase treatment in a manner similarto that offreshlyisolated 30% fraction.
The combined data from Fig. 1 to 4 are interpreted to
0 10 20 30
fractions FIG. 3. Invitro RNAsynthesis ofa30% vesicularfraction for15 min (*), 30 min (-), 45 min (A), and 60 min (-). [3H]UTP is incorporatedfirst into RI andgradually also into 36S RNA. RNA
wasanalyzedasdescribed inthe legendtoFig. 1.
dpm
40000 36S
30000-
20000-10000
0
0 10 20 30
fractions FIG. 4. Invitro RNAsynthesis ofa45% vesicular fractionfor15 min(O),30 min(O),45 min(A), and 60min(0). Thekinetics and pattern of plus-strand RNA synthesis are very similar to those obtained with30%fractions. RNA wasanalyzedasdescribedin the legendtoFig. 1.
meanthat the 30% fraction, since it contains RI,synthesizes
RNA in vivo and continues the synthesis in vitro, whereas the 45% fraction seems to restart its plus-strand RNA
synthesisin vitro.Whether thishappens aselongation, e.g., on a small amountof RI (trailing shoulder in Fig. 1) or as initiationon RFor RIcannotbe determinedatpresent(see Discussion).
Ultrastructural aspects of the 30 and 45% vesicles. As statedearlier (6),the poliovirus replication complex is func-tionalonly if, bymeans of viral protein 2C, it is associated with the membranes of the virus-induced vesicles. In the currentinvestigation, we found that the virus-induced
vesi-cies
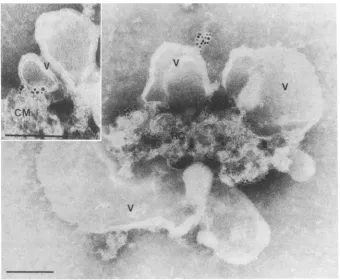
are not the only membranesystem associated with the replication complex. A second type ofmembrane was ob-served in thereplication complex of bothvesicularfractions(Fig. 5).These membranes are tightly packed and appear as small (approximately 50-nm), clustered, vesiclelike bodies
(compact membranes). Like the membranes of the virus-induced vesicles, these compact membranes were labeled with2C-MAb(Fig.5). The label was foundpredominantlyat theborder of thereplication complexatthesiteswhere there is intimate contact between the compact membranes of the replication complex and the membranes of the virus-induced vesicles.
The ultrastructural aspects of the compact membranes within the replication complex are the same in the 30 and 45% fractions and also do not change during in vitro tran-scription. In ultrathin sections of poliovirus-infected cells, the compact membranes appear tightly coiled and do not have a clearly defined shape (data not shown).
DOCtreatmentof vesicularfractions. Totestwhetherthe replicationcomplex-associated membranes described above are involved in the observedprotection of the RI, afreshly isolated, RI-containing 30% fraction was treatedwith DOC for 30 min at 0°C. This rendered the RI partially RNase susceptible,asthe RIcould now be convertedbyRNaseinto an 18S core (10) (Fig. 6). After phenol extraction, the 18S corewasRNase sensitive(data notshown), which indicates that this 18Sstructure is derived from theRI (andnot from the RF) and that it is enclosed inpresumably proteinaceous material.
Together with the findings illustrated in Fig. 2, these
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.69.309.77.242.2] [image:3.612.320.564.78.239.2] [image:3.612.68.308.530.712.2]~~~U1
FIG. 5. Electronmicrographofapoliovirusreplicationcomplexsurroundedbyvirus-induced vesicles(V)andcontainingasecond, compact
membranesystem (arrowheads). Immunocytochemical labelingwith2C-MLAb and 5-nm colloidal gold shows thatboth membrane types are
labeled. Themicrographshowsa45% vesicular fraction. Nomorphologicaldifferences from the 30% fractions could be found. Bar,100 nm.
findings indicate that themature36S RNA is located outside of thereplication complex,whereas thenascentplusstrands of the RI are sequestered by membranes. DOC treatment converts the replication complexinto amembrane-deficient
dpm 36 - 23- 16S
200 -_ _ _ -
-20000
t
U0 10 20 30
fractions FIG. 6. RNasetreatmentofa30%vesicularfractiondigests only 36S RNA and leaves the RI intact (A) (Fig. 2). The RI can be digestedto an 18S coreonly afterDOC treatment (-), indicating that membranes protect thegrowing plus strands. RNAwas
ana-lyzedas described in thelegendtoFig. 1.
matrix containing the RIwith protruding, and thus
RNase-sensitive, nascentRNA strands.
Since the 30 and45% fractionsare
morphologically
and in vitro alsofunctionally identical, thefunction(s) of therepli-cation complex-associated membranes during in vitro viral RNA synthesis was investigated with 45% vesicular frac-tions.
DOCwasaddedtoan invitrotranscriptionsystemafter 30 minofongoing in vitroRNAsynthesis. Duringfurther RNA
synthesis, the amount ofradioactivity increased in the RI,
whereas theamountof36SRNAremainedstableatthe level reached when DOC was added (Fig. 7). Thus, it can be concluded that themembranesof the
transcriptionally
activereplication complexes not only sequester the nascent plus
strands but alsoareinvolved in their detachment in
becom-ing36S RNA.
Ultrastructural analysis of
replication complexes
after treatmentwithDOCshows that thevirus-inducedvesiclesas well as the compact vesiclelike structures within therepli-cation complexhave completelydisappeared
(Fig. 8).
This indicates that these compact structures indeed consist of membranes. The membrane-deficient matrix of thereplica-tion complexstillcontains P2protein atits
periphery.
Role of membranes and location ofprogenyvirus RNA in thereplicationcomplexasrevealedbyRNase-gold histochem-istryand insituhybridization. The observations
presented
so far indicate that the replication complex is a membrane-containing structure that encloses the RItightly
and ison November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.138.470.79.398.2] [image:4.612.54.296.522.683.2]dpm
36 23-16S
30000 - l l
20000-10000 L
0'
0 10 20 30
fractions FIG. 7. Invitro RNA synthesis of a45% vesicular fraction was allowed to proceed for 30 min, and RI and 36S RNA were made (A). To a parallel tube (O), DOC was added after 30 min of in vitro synthesis, and the reaction was allowed to proceed for another 15 min; only RIcontinued to incorporate [3H]UTP,whereas radioac-tivity did not increase in the 36S moiety. RNA was analyzed as described in thelegend to Fig. 1.
surrounded by a rosette of virus-induced vesicles. On the basis of the ultrastructural data(Fig. 5), it seemed to us that the resistance of the nascent plus-strand RNA against RNase reported above might be mediated not by the rosettes of large, virus-induced vesicles (which do not seemtobe in an extremely tight arrangement) but by the densely packed compact membranes within thereplication complex.
Toidentifythe membrane systemprotectingthe RI andat thesame timeto obtainmore information about the topog-raphy of thereplicationcomplex,weexploredthe location of
the 36S RNA after in vitro RNA synthesis, adopting the following rationale. Since only the 36S RNA and not the RI was found to beaccessible to RNase, only the 36S RNA can be located by histochemistry and in situ hybridization. The observedlocation of the 36S RNA would then be indicative of thetightness of the two membrane systems.
InFig. 9, RNA in general was identified by a histochem-ical reaction using RNase coupled to colloidal gold. In Fig. 10,viral 36S plus-strand RNA was localized specifically by in situ hybridization with a biotinylated riboprobe comple-mentary to a partof theP1 genomic region. Both reactions clearly show their respective targets located within the rosetteof thevirus-induced vesicles, thus demonstrating the rather loose and in any casepermeable configuration of the vesicular rosette and suggesting that the central compact membranes are the protecting agents of the RI. It is note-worthythat the 36S RNA is set free at several distinct sites at the periphery of the replication complex. This can be explained by assuming a topography of the viral RNA replication in which the template moves through the repli-cation complex and has the progeny RNA strands fixed at defined sites.
DISCUSSION
From poliovirus-infected cells, two populations of mem-brane-bound replication complexes can be isolated. One population bands in 30% sucrose and actively synthesizes
36S plus-strand RNA in vivo and in vitro. The fraction banding in 45% sucrose contains capsid precursors and virions (20) and besides encapsidated and free 36S RNA contains little, if any, RI. In vivo, the 45% fraction is therefore considered to be transcriptionally silent ("burnt out") but contains progeny RNA in the process ofbecoming
encapsidated. In vitro, it is capable of resuming RNA
I ..
*,
..4~f
V-kN .
.0-FIG. 8. ElectronmicrographofaDOC-treatedreplicationcomplex(45% fraction). The membranesof the virus-induced vesiclesaswell
asthe compact membranes within thereplicationcomplexaredissolved. Theimmunocytochemical label with 2C-MAb isatthe surface of thecorestructureof the replicationcomplex. Bar, 100nm.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.65.306.78.239.2] [image:5.612.125.514.456.696.2]FIG. 9. Avesicle (V)-bound replication complex (RC) as in Fig. 5 but doubly labeled with 2C-MAb (small gold grains) and with RNase-gold (large gold grains). The 36S RNA is located between the virus-induced vesicles and the replication complex. Bar, 100 nm.
synthesis and producing 36S RNA (Fig. 4). It is not clear whether in vitro the plus-strand RNA is elongated only on a small amountof RI or whether there is true initiation, which could take place either on preexisting RI or on RF, which could already be present in small quantities in vivo orcould have been freshly synthesized in vitro.
The two typesof viral replication complexes show similar ultrastructural features. They are enclosed in a rosettelike shell of virus-induced vesicles, which are attached to the replication complex by means of viral protein 2C (6). In the present investigation, we identified a second, very compact membrane system within the replication complex itself (Fig. 5). It encloses tightly the RI, which in concert with viral proteins represents the actual plus-strand RNA-synthesizing machinery. This was shown by our DOC experiments: removal of all membranes with DOC leaves the core of the replication complex, which still incorporates [3H]UTP. The finding that under these conditions [3H]UTP was incorpo-rated only into RI and not into 36S RNA indicates that elongation takes place but that no completed 36S RNA is released. Theelongation proceeds forquite some time, albeit at a reduced rate compared with that of non-DOC-treated replication complexes or intact cells.
IfDOC-treated cores of replication complexes devoid of membranes are treated with RNase A, the nascent plus strands at the RI areexposed to the action of the enzyme, whereas the partially double-stranded template with the replication forks remains protected from the RNase by a substance removable by phenol. RNase treatment of non-DOC-treated, transcriptionally active replication complexes
FIG. 10. Avesicle(V)-bound replication complex(RC)(45%fraction)onwhich insituhybridizationwithabiotinylatedRNAprobewas performed.Theprobe is visualized with6-nmgold-coupledanti-biotin antibodies. Thehybridization signal, localizingthe 36SRNA,is found onthesurface of thereplication complexbut within therosetteofthe virus-inducedvesicles.(Inset)Highermagnification.Thehybridization signalis found wherethe compact membranes(CM)of thereplication complexarein closecontactwith thevirus-induced vesicles.Bars,100 nm.
.:-...0.4.
.. -low
.,P:
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.132.472.396.676.2]digests only36SRNA whileleavingthe entire RI intact. This means that freshly completed 36SRNA is rapidly set free. Since the completed 36S RNA is found by RNase-gold as well as by in situ hybridization within the rosette of virus-inducedvesicles but outside thereplication complexper se, it is possible that the compact membranes within the repli-cation complex have to interact with the virus-induced vesicles for the detachment of the 36S RNA from the template.
At the samesite of thisputative interaction,the presence of
protein
2C(and
the other2C-containing
viral P2proteins)could consistentlybe demonstrated by
immunocytochemis-try (Fig. 5 and 9). Its presence in the replication complex even after DOC treatment indicates that it is not only
membrane associated but also boundto other components. This is in agreement with the findings that it is RNA associated and that it attaches viral RNAto the surface of the virus-induced vesicles
(6).
The 2C-RNA association is consistent with thefinding
that2C containsNTP-binding (8) aswellashelicase(12)motifs.Itmightwell be that 2C is also involved in the liberation of the RNA from the replicationcomplex
andpossibly
in preparatory steps forencapsidation(16).
Whatever theexactfunctions of P2proteins2BC, 2B, and 2C
might
ultimately be, the action ofone or more of them wasfoundtobe essential for viralplus-strandRNAsynthe-sis
(6, 13, 15);
forhepatitis
Avirus(9, 14), rhinovirus (17),and
poliovirus (21) replication
in cellculture;
and for expres-sion ofcytopathic
effects(5).
Since these P2protein-medi-ated processes are interconnected
(4),
one might speculatethat these
proteins
donotfunctionontheirownbut functiononly in concert with each other and/or otherviral or even cellular proteins. This would also explain why, despite numerousattempts, e.g.,bymutationalanalysis,it is stillnot
possible
to attribute abiochemicallydefined roletoeach of theseproteins.
In our in vitro system, encapsidation of progeny RNA could not be observed. This is in contrastwith a recently
published
report(18)
andmight
beduetoseveralexperimen-taldifferences. Our in vitro system isatranscriptionsystem, not a coupled transcription-translation system, and it was
kept going
foramuch shorter time. Animportant aspect of thisstudy
could be thefinding
that the 36S RNA is setfree from the replication complex immediately after itscomple-tion. This is in agreement with the
finding,
demonstratedbyin situ hybridization on infected cells, that a pool of free
genomic
RNAwaspresent in thevicinity ofthe replicationcomplex (27).
Inthe invitro system, thisRNAthenmightbe lost for encapsidation, because the RNA-synthesizing enti-ties are rather diluted in vitro compared with the in vivo situation.The data presentedhereallowustopropose thefollowing model ofthe poliovirus replicationcomplex. The replication
complex
contains a compact membrane system which en-closes the RIwith its nascent plus-strand RNA. The repli-cation forks are in additiontightlyenclosed in protein. The compact membranes are involved in the liberation of thecompleted 36S RNA from the RI and from the replication
complex.
The progeny strandsaresetfreeatdifferentpoints onthereplication
complex's surface, i.e.,atthesites where thereplication
complex is attached to the virus-induced vesicles.During
RNAsynthesis, theRI would thushave to move(turn)
through, ortogether with, the replicationcom-plex
with respect tothe virus-induced vesicles.ACKNOWLEDGMENTS
Wethank Bea Herzog for excellent technical assistance. This work was supported bygrant 31-29910.90 from the Swiss National Science Foundation. M.T. wassupported byagrantfrom the Freiwillige Akademische Gesellschaft Basel, Basel, Switzer-land.
REFERENCES
1. Baltimore, D. 1968. Structureof thepoliovirusreplicative inter-mediate RNA. J. Mol. Biol. 32:359-368.
2. Bendayan, M., and E. Puvion. 1983. Ultrastructural detection of RNA:complementarityofhigh-resolution autoradiography and ofRNase-gold method. J. Ultrastruct. Res. 83:274-283. 3. Bienz, K., D. Egger, and L. Pasamontes. 1987. Association of
polioviral proteins of the P2 genomic region with the viral replication complex and virus-induced membrane synthesis as visualized by electron microscopic immunocytochemistry and autoradiography.Virology 160:220-226.
4. Bienz, K., D. Egger, Y. Rasser, and W. Bossart. 1980. Kinetics and location ofpoliovirus macromolecular synthesis in correla-tion to virus-inducedcytopathology. Virology 100:390-399. 5. Bienz, K.,D.Egger,Y.Rasser, andW.Bossart. 1983.
Intracel-lular distribution ofpoliovirus proteins and the induction of virus-specific cytoplasmic structures. Virology 131:39-48. 6. Bienz, K., D. Egger, M. Troxler, and L. Pasamontes. 1990.
Structuralorganization ofpoliovirus RNA replicationis medi-ated by viral proteins of the P2 genomic region. J. Virol. 64:1156-1163.
7. Dernick, R., and P. Bilello. 1974. Absence of glycoproteins in poliovirus particles.J. Gen. Virol. 25:125-132.
8. Dever, T. E.,M. J. Glynias, andW. C. Merrick. 1987. GTP-bindingdomain: three consensus sequence elements with dis-tinctspacing.Proc. Natl. Acad. Sci. USA 84:1814-1818. 9. Emerson, S. U., C. McRill, B. Rosenblum, S. Feinstone, and
R. H. Purcell. 1991. Mutations responsible for adaptation of hepatitis A virus to efficient growth in cell culture. J. Virol. 65:4882-4886.
10. Etchison, D.,and E.Ehrenfeld. 1981.Comparison of replication complexes synthesizing poliovirusRNA.Virology 111:33-46. 11. Girard, M. 1969. In vitro synthesis of poliovirus ribonucleic
acid: role of thereplicative intermediate. J. Virol. 3:376-384. 12. Gorbalenya,A.E.,E. V. Koonin, and Y. I. Wolf. 1990. Anew
superfamilyofputative NTP-bindingdomains encoded by ge-nomesof small DNAand RNA viruses. FEBS Lett. 262:145-148.
13. Johnson, K. L., and P. Sarnow. 1991. Three poliovirus 2B mutantsexhibitnoncomplementable defects in viral RNA am-plificationanddisplay dosage-dependentdominance over wild-typepoliovirus. J.Virol. 65:4341-4349.
14. Lemon, S. M.,P. C. Murphy, P. A. Shields, L.-H. Ping, S. M. Feinstone,T.Cromeans, and R. W. Jansen. 1991. Antigenic and geneticvariation incytopathic hepatitisAvirus variants arising during persistentinfection: evidence for genetic recombination. J.Virol. 65:2056-2065.
15. Li, J.-P., and D. Baltimore. 1988. Isolation of poliovirus 2C mutants defective in viral RNA synthesis. J. Virol. 62:4016-4021.
16. Li, J.-P.,and D. Baltimore. 1990. Anintragenic revertant of a poliovirus2Cmutanthasanuncoating defect. J. Virol. 64:1102-1107.
17. Lomax,N.B., and F.H. Yin.1989. Evidence for the role of the P2 protein of human rhinovirus in its host range change. J. Virol. 63:2396-2399.
18. Molla,A.,A. V.Paul,and E.Wimmer. 1991.Cell-free, de novo synthesisofpoliovirus. Science 254:1647-1651.
19. Pasamontes, L., D. Egger, and K. Bienz. 1986. Production of monoclonal and monospecific antibodies against non-capsid proteinsofpoliovirus. J. Gen. Virol.67:2415-2422.
20. Pfister,T.,L.Pasamontes, M.Troxler,D. Egger, and K. Bienz. Immunocytochemical localization of capsiol-related particles in subcellular fractions ofpoliovirus infected cells. Virology, in press.
21. Pincus, S. E.,D. C. Diamond, E. A. Emini, and E. Wimmer.
on November 9, 2019 by guest
http://jvi.asm.org/
1986. Guanidine-selected mutants of poliovirus: mapping of point mutationstopolypeptide 2C. J. Virol. 57:638-646. 22. Racaniello, V. R., and D. Baltimore. 1981. Cloned poliovirus
complementary DNA is infectious in mammalian cells. Science 214:916-919.
23. Sambrook,J., E. F. Fritsch, and T. Maniatis. 1989.Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
24. Takeda, N., R. J. Kuhn, C.-F. Yang, T. Takegami, and E. Wimmer.1986. Initiation of poliovirus plus-strand RNA synthe-sis in a membrane complex of infected HeLa cells. J. Virol.
60:43-53.
25. Takegami, T., B. L.Semler,C.W.Anderson, and E. Wimmer.
1983. Membranefractions active in poliovirus RNA replication containVPgprecursorpolypeptides. Virology 128:33-47.
26. Troxler, M., L. Pasamontes, D. Egger, and K. Bienz. 1990. In situhybridizationforlight and electron microscopy. A
compar-ison ofmethods for the localization of viral RNA using biotiny-lated DNA and RNA probes. J. Virol. Methods 30:1-14. 27. Troxler, M., T. Pfister, D. Egger,and K. Bienz. Submittedfor
publication.
28. Young, D. C., D. M. Tuschall, and J. B. Flanegan. 1985. Poliovirus RNA-dependent RNA polymerase and host cell protein synthesize products twice the size of poliovirionRNA in vitro. J. Virol. 54:256-264.