JOURNAL OF VIROLOGY,
JUlY
1990, p. 3463-3470 0022-538X/90/073463-08$02.00/0Copyright© 1990,American Society for Microbiology
The Herpes Simplex
Virus
Us11
Open Reading
Frame
Encodes
aSequence-Specific
RNA-Binding
Protein
RICHARD J. ROLLERANDBERNARD ROIZMAN*
TheMarjorieB. Kovler Viral OncologyLaboratories, The University ofChicago, Chicago, Illinois 60637
Received 21 February 1990/Accepted12April 1990
Herpes simplex virus 1- and 2 (HSV-1 and HSV-2)-infected cell extracts but not uninfected cell extracts
containanRNA-bindingactivity foraninvitro-transcribedsequencefromthedomains of theHSV-1Us11and
a47genes.The transcript of thissequencehasnotbeen detected ininfectedcells. The binding issequenceand
secondary structure specific and protects approximately 95 nucleotides from RNase digestion. Analyses of HSV-1 x HSV-2 recombinants and HSV-1 deletionmutantsmapped the functionnecessaryfor activitytothe
Us11
ora47 open readingframe. The a47genewas excluded, since the RNA-binding activity is alate (Y2)function dependent on viral DNA synthesis for its expression. The
Usll
function is theonly viral function required, since translation in rabbit reticulocyte lysateofaninvitro-synthesizedUs11 mRNAresulted in theappearance of the RNA-bindingactivity. The product of theUs11open readingframe is associated with the
RNAprobe-proteincomplexinasmuchasinsertionofa sequenceencodingin frame 15 additional aminoacids attheC terminusof the
Usll
protein causedacorrespondingdecreasein theelectrophoreticmobility of thebindingcomplex.
Inthispaper wereportthataprotein encoded byan open
readingframe in thesmallcomponentof theherpessimplex virus 1 (HSV-1)genome participates in the formation ofa
sequence-specific RNA-protein complex in vitro. This is the firstreportofanHSV protein withsequence-specific affinity
forRNA. Relevanttothisreport arethefollowing.
(i)The 74 known openreading frames ofHSV-1 (1, 3, 5,
24,25)formseveralgroupswhoseexpression iscoordinately
regulated and sequentially ordered inacascade fashion (10,
11). Among the products of theagenes,the firstgrouptobe expressed, isthe product of the a4gene, designatedas the
infected cellproteinno.4(ICP4). This protein bindstothe 5' untranscribed and transcribed noncoding domains of ,B and 'Yi and _Y2 genes
expressed
later in infection(14).
Because ICP4 binds to transcribed noncoding domains of Y2genes,thatis,genes stringently dependentonviral DNAsynthesis
for theirexpression,thereexisted theformalpossibility that ICP4bindstoRNAaswellasDNA,apossibility that isnot too farfetched in light ofthe discovery of a host protein which binds single- and double-strandedRNA and DNA in sequence-specific fashion(36). Byafortuitous coincidence,
we chose to test the transcripts of the _Y2
US11
(13) openreading frame. While we were unable to show that ICP4 bindstoRNA, wediddiscoverthat
Usll
proteinbinds toatranscript complementarytoitsdomain.
(ii)Theprotein productof the
Usll
genewasfirst identi-fied, using hybrid arrest oftranslation, by Rixon and Mc-Geoch (33) and was shown to be a protein with an Mr of 21,000. Subsequently, usinganantiserumdirectedagainst apeptide sequence predicted by the nucleotide sequence of the
Usll
openreading frame,Johnson etal.(13)showedthat theUsll
protein actuallyconsists ofadoubletwithanMrof 21,000to22,000and that theexpressionofUsll
isregulatedas aY2ortruelategene.Johnson et al.(13)alsoreportedthat
the size ofthe
Usll
proteinvariedbetweentwoof the strains tested-an observation which had been predicted on the basis of the differencesin thesize ofthe openreadingframe found in two different strains of HSV-1 (33, 43). These*Correspondingauthor.
results were consistent with those of Lonsdale et al. (20), whohad earlier shown thatadoublet of proteins withanMr
ofapproximately 20,000 fromanumber of HSV-1 laboratory
strains and isolates passaged a limited number of times
outside the human hostvariedsignificantlyinits electropho-retic mobilityindenaturingpolyacrylamide gels.
(iii)Ourlaboratory and otherlaboratories have reporteda
large numberof HSV-1genesthataredispensable for growth
in cells in culture. Relevant to this report are studies
demonstrating that, with one exception, all of the genes
mappingin the uniquesequences of the S component
(Us),
including theUsll
openreadingframe,aredispensable (19).None of the studies done to date in our laboratory or
reported elsewhere have shown that the
Usll-
HSV-1 strains are deficientfor replication in any of the cell linestested.
MATERIALS AND METHODS
Cells and viruses. HSV-1 strain F [HSV-1(F)], HSV-2 strain G [HSV-2(G)], and deletion mutants R325, R7023, R7039, R3630, and R3631 weredescribed elsewhere (7, 18, 23, 27, 29, 35). HeLacells (American TypeCulture Collec-tion)werepropagated asdescribed elsewhere (14).
Construction andcloningofplasmids. Standardprocedures
wereused in allconstructions described in this report (22).
The structures of plasmids pRB3881, pRB3910, and pRB4027anddetails of their constructionaregiveninFig. 1. Construction of recombinant virus. The recombinant
vi-ruses used in this study (R325, R7023, R7039, R3630, and R3631) and the one recombinant virus constructed for this studywere constructed by double homologous recombina-tion betweenthe intactDNA ofaparentvirus andplasmid DNA in which nonhomologous sequences and a selectable markerweresandwichedbetweenflankingsequenceswhose homologswerepresentin the sameorder andorientationin
the genomic DNA. The selectable marker was the HSV-1 thymidinekinase(tk)geneasdescribedbyPostand Roizman (29), except that the natural promoter of the tk gene was
replaced bythea27promoter(19). RecombinantR3630was
constructed by replacementof thecx47andUs11geneswith
3463
Vol. 64,No. 7
on November 10, 2019 by guest
http://jvi.asm.org/
3464 ROLLER AND ROIZMAN
A[
2f~
ji
B "Yin -M Deletion
-R7023
1 R7039 _
R325 U
R3631 U
C
DI
gl usl1o
F-*
(H-S2 G gD> US9 USi1
2 a4 a22 US3 US5 gE a47 x4
a. c' U c a
- r~zz~~
USi 1
a47
1111 Oris RNA
Sal SacSac Bam Rsa,Bst Rsa Sac Eco
pRB421
T7
2 SmaBst Sty Sma ThaPst
U- 1 ---, pRB3881
: ' ' ~~~~~~~SP6
3 Eco BamSac Xho Xho
pRB3910
iSal Ava Bar Sac Ect
pRB421 5bp Synthetic Tag , 07bpAva/Sac Fragment 2m/AvaCompatibleEnds|- ; va/SacCompatibleEnds
Sal Bam,Sac Eco
4, Bam/Sac 1-cutpRB421
3 US1O(truncated)
< USll(tagged)
Sal ,p,5jac§ Eco
pRB4027
FIG. 1. Sequence arrangement of the HSV-1 genome and S componentand constructionofplasmids.(A)Schematic diagram of the HSV-1 genome in the prototype arrangement, showing the unique sequences (lines) flanked by inverted repeats (boxes). (B) Expansion of the S component of the genome. Bi, Extent of deletions intheshortcomponent of deletion mutants usedinthese studies. Inthe genome of R7023, theScomponent is frozenin an invertedorientation. Thedeletion extends beyond the L-Sjunction intotheinvertedrepeatsflankingtheScomponent.B2,Genetic map of the short component for comparisonwithBi. (C)Expansionof theregion oftheS componentcloned aspRB421andconstructionof plasmidspRB3881and pRB3910.Cl, Restrictionandgeneticmaps ofpRB421 (23). Verticalbarsindicatetranscription initiation sites; thin lines indicated transcribed noncoding domains; solid boxes indicate open reading frames ot47,
Us11,
andUs10.
a47 and Oris RNAtranscripts initiate within the invertedrepeat sequence; Us11 andUs10transcripts initiate withinUs. Restriction enzyme abbre-viations: Sac, SacII; Bam, BamHI; Bst,BstEII; Rsa, RsaI, Sal, Sall;Eco,EcoRI. Thelisting of RsaI sitesis notcomplete; only the relevant sites are indicated. C2, Construction of pRB3881 probe template. The202-ntRsa-Rsafragment containing 158 nt ofUs11
5'-untranslated sequence and 44 nt of5'-flanking nontranscribed sequence was cloned into the HincIl site of pUC8 and then subclonedas aHindIII-EcoRIfragmentintopGEM-3Z. Thin lines indicate sequences transcribed by T7 RNA polymerase; the open box indicates the minimumregionessential for binding (see text). Forprobe transcription, plasmidwaslinearized at thePstIsite and transcribed with T7 RNA polymerase toproduce a245-nt probe. Restrictionsitesused togeneratedeleted probes: Bst,BstEII;Sma, SmaI; Sty, StyI; Tha, ThaI; Pst, PstI. 5' deletionswere made by collapsing the template between thepolylinkerEcoRI site and the appropriate site intheinsert.3'deletions were made bylinearizing pRB3881 at sites within the insert. C3, Construction ofpRB3910 plasmid template for
Us11
mRNA. The650-base-pairBstEII-SacIIthecx27-tkgene inahomologousrecombination between the genome ofa tk-negative virus anda plasmidcontaining the
a27-tk gene sandwiched between the appropriate homolo-gous sequences. The recombinant was selected by
plating
the progeny of the cotransfection in medium containing methotrexate, which selects for the tk+ progeny. Recombi-nant R4027 was constructed by cotransfection of intact R3630 DNA andpRB4027 plasmid DNA containinganinsert in the
Usll
open readingframeas described in Results. In this instance, the tk gene was replaced by the HSV-1 sequences contained inpRB4027.In vitro transcription. Plasmid templates were prepared
and transcribed with SP6 and T7 RNA polymerases as recommended by Promega Biotec, except that all RNAs were capped during synthesis with the cap analog GppG (New England BioLabs) as described previously (28). La-beled binding probe was synthesized with 10 1Ci of [32P]CTP ina 10-,ul reaction mixture supplemented with 50 ,uM unlabeled CTP. Reaction products were separated on
denaturing4% acrylamideureagelsand visualizedby auto-radiography of the wet gels. The band corresponding to full-length probe was excised, and RNA was eluted by soaking overnight in 0.5 M ammonium acetate-1 mM EDTA-0.2% sodiumdodecyl sulfate. Eluted RNA was pre-cipitated with ethanol in the presence of 10 pLg of carrier glycogen and suspended inwater.
Capped synthetic
Usll
mRNA for in vitro translationwas transcribed from EcoRI-linearized pRB3910 by the same procedure, except that the concentration of all NTPs was 0.5 mM andonly 0.5 ,uCi of [32P]CTPwasused asatracer. The product was quantitated by DE-81 filter assay as described elsewhere(22).Assays for RNA-binding activity. For gel shift assays, labeledprobe (10,000to20,000 cpm) was treated with 5 plg of nuclear protein extract in 7
[l
ofbinding buffer (200 mM NaCl, 20 mM Tris [pH 7.9 at 4°C], 2 mM MgCl2, 0.5 mM dithiothreitol, 0.2 mM EDTA, 0.2 mMphenylmethylsulfonylfluoride, 20%[vol/vol] glycerol) for 20min at room temper-ature, followed by addition of heparin to 5 mg/ml and reaction for a further 10 min. Onemicroliterofloadingdye (100%glycerol, 0.4%bromphenol blue, 0.4% xylene cyanole FF)was then added, and samples were applied to a nonde-naturing 4%acrylamide gelpoured and run in 0.5x TBE(45 mM Tris, 45 mM boric acid, 1.25 mM EDTA). Protection
fragment of pRB421 containing theUs11 openreading frame, 76nt of5'-untranslated sequence, and 90 nt of3'-untranslatedsequence wasexcised, converted to blunt ends by treatment with the Klenow fragment and dNTPs, and cloned into the SmaI site ofpGEM-3Z. Linearization with EcoRIrestrictionenzymefollowedby transcrip-tion withSP6RNApolymeraseproducedUs11mRNA. Restriction site abbreviations: Eco, EcoRI; Bam, BamHI; Xho, XhoI. (D) Construction ofpRB4027 encodingtagged Us11 protein. Two in-serts were prepared asfollows. (i) The 107-base-pairAvaIl (Ava)-Saclfragment of pRB421 (D1) encodingUs11codons111to 146 was excised and purified from a low-melt agarose gel. (ii) Synthetic oligonucleotides (94 and 95 nt) wereannealedtogeneratean insert encodingUs11 codons 147 to 161followedby asequenceencoding amino acids 71 to 85 of HSV-1 ICP4 and a UAA stop codon. The annealed double-stranded insert had overhanging BamHI- and AvaII-compatible ends. These inserts were coligated (D2) into SacI/BamHI-cutpRB421 togenerate pRB4027 (D3). The newUs11 openreadingframe encoded acompleteUs11plus tag. The insertion of the tag introduced a stop codoninto theUs10open reading frame, truncatingit such that only 110 codons should be translated. The construction also resulted in the duplication of the sequences between the BamHI and Avallsites.
r---
---U.Y6
---F e
I
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.66.298.77.415.2]HSV RNA-BINDING ACTIVITY 3465 assays were identical, except that before addition of
heparin,
reaction mixtures were digested with 5 U of RNase T1 for 10 min.
Preparation of cellular extracts. Small-scale nuclear ex-tracts (used for experiments shown in Fig. 5) were prepared asdescribed by Lee et al. (16) from HeLa cells infected with 10PFU per cell and harvested 20 h after infection. Large-scale nuclear extracts (used for all other experiments) were prepared as previously described (14).
Sephacryl S300 chromatography. One milliliter of nuclear extract was fractionated on a 55-ml column of Sephacryl
S300 run in binding buffer at a linear flow rate of 8
cm/h;
0.5-mlfractions were obtained.Preparation ofcompetitor nucleic acids. Competitor RNAs were synthesized and purified in the same way as probe RNA, except that reaction mixtures contained all NTPs at 0.5 mMwithout label, and the product was excised from gels
byusingcomigrating labeled product as a standard. Purified
competitor RNAs were quantitated by
A260
Competitor DNAs were purified from low-melt agarose gels as described elsewhere (22) and quantitated by coelectrophoresis with DNAsof known quantity.In vitrotranslation. Synthetic
Us11
mRNA (0.3 ,g) was translated in a 20-,ul reaction mixture by using a kit obtainedfromPromega Biotecaccording to the protocol supplied with the kit, except that after the suggested incubation period, unlabeledmethionine was added to 100 puMand incubation
wascontinuedfor afurther 30min. After the reaction, the
mixtureswerediluted to 50
,lA
with binding buffer and passed over a 1-ml Sephadex G-50 spin column which had beenpreequilibrated inbinding buffer.
RESULTS
Infected-cell extracts contain an RNA-binding activity.
pRB3881 (Fig. 1C) contains 5'-transcribed noncoding se-quences of the Y2
US"1
open reading frame. Uniformlylabeled in vitro transcripts of each strand of the HSV-1 sequences inpRB3881 werereacted with apartiallypurified preparation of ICP4 derived from Bio-Gel A.5m
chroma-tography ofcrude HSV-1-infected cell nuclear extract (P. McAtee and B. Roizman, unpublished results), and the
reaction mixtures were electrophoretically separated on a
nondenaturing polyacrylamide gel. Only one ofthe RNA
probes,i.e.,thatcorrespondingto theoppositestrand of the
Usll
open reading frame 5'-untranslated sequence, exhib-ited a slowerelectrophoretic
mobility(Fig.
2A,lanes 1 and 2).Digestion ofthe mixture withproteinase Keitherbefore or afterthe reaction(Fig. 2A,
lanes 3 and 4,respectively)
causedthe RNA
probe
to movewith normalmobilityonthegel,
showing
thattheretardation oftheprobe
wasduetotheformation ofanRNA-protein
complex.
Because thedegreeof retardationappearedtobetoosmall to be caused by a
protein
aslarge
as ICP4 and becauseaddition of anti-ICP4
antibody
to the incubation failed to causeafurther decrease in themobilityof theprobe,ashas been shown for ICP4-DNA complexes (14), it seemed un-likelythat the retardationof theprobe
was dueto ICP4.RNA-bindingactivity issecondary structure
specific.
RNAprobe transcribed from
pRB3881
wasalways
purified
as a single tight band fromdenaturing
polyacrylamide
gels.
Innondenaturing gels, however, it
migrated
as twospecies.
The
faster-migrating
species
composed
70% ofthe label andformed a
relatively
diffuseband,
whereas theslower-mi-grating
formcomposed
30% of the total and formed arelatively tight band
(Fig. 2A,
lane1).
Elution of eitherA B-
-ProltK~~
E Co E.2)
A.5m
E S S 3 E , ¢~~E c
Protein > >
I.Q cL <I )
;K3: <
_
_
_ ^
_ _
.w.. _
..:..._
I.
-u
42
h
123 4 1 2 3 1 2 3 4 5
FIG. 2. Demonstration of bindingto specific probe and protec-tionof probe fromRNaseT1 digestion.(A)Autoradiographicimage ofpRB3881 T7 transcript probe reacted as describedin Materials and Methods with binding buffer (lane 1), 7
[lI
ofBio-Gel A.5m fraction(lane 2),7,ulofA.5mfractionpreviouslydigested with 200p.g
ofproteinaseK(Prot. K)permlfor30minat25°C (lane 3),or7 ,ulof Bio-GelA.5mfraction subsequentlydigested withproteinase K (lane 4). (B)Autoradiographic image of pRB3881 T7transcript probe reactedwith 7 ,ul of Bio-GelA.5mfraction(lane 1), 5 ,ulof HSV-1(F)-infectedcell nuclearextractfollowedbyRNaseT,
diges-tion (lane 2), or 5 jg of HSV-2(G)-infected cell nuclear extract followedbyRNase T1 digestion (lane 3). Arrowheads indicate the positions ofspecific RNase-resistant RNA-protein complexeswhich wereexcised from the gel.(C)Autoradiographic image ofdenaturing electrophoresis of RNAs isolated from the complexes shown in panelB. Lane 1,pRB3881 T7transcript probe RNA; lane 2,RNA fromA.5mfractionbinding complex;lane3,protected RNA frag-ment from HSV-1(F) binding complex; lane 4, protected RNA fragment from HSV-2(G) complex; lane 5, MspI-digested, end-labeled pGEM-3Z molecular weight (MW) standards (fragment lengths shownatright).species from gel slices followed
by
electrophoresis
on an-othernondenaturing gel
resulted in thereappearanceofbothspecies in the same ratio (not
shown).
Theprobe
species
were interconvertible and
apparently
represent different secondary structuresadopted by the RNAprobe.
Only
the slowerspecies
formedcomplexes
with the infected cell proteins (Fig. 2A, lanes 1 and 2;Fig.
3B, compare lanes 1 and 2through 14).RNA-binding
activity
issequencespecific.
The RNAs tested todate includetranscripts
of both ends of theunique
shortregion ofthe HSV-1 genome and sequences from the
5'-untranslated
region
ofthe HSV-1 late geneencoding
the a genetrans-inducing
factor(aTIF).
Inall,
severalkilobasesof RNA sequences have beentested,
and no other sequenceVOL. 64, 1990
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.353.533.73.382.2]3466 ROLLER AND ROIZMAN
B
CompetitorRNA Competitor DNA
Pst 3881 t,nci {,Ta3881 Vc.cI.r i.
-CO t:30G
qI'O1- _%£tCMCn
.
~~~~CM
M v iC13 -> w C) CO L. n [image:4.612.129.460.77.247.2]1t1112 13 4 15 1
FIG. 3. Autoradiographic images of complexes formed betweenA.5m fraction protein andpRB3881 T7transcript probe RNA in the
presenceofvarious competitor RNAs andDNAs. Arrowheads indicate thepositionsof theRNA-protein complexes. (A) Competition with
specific (lanes 3 to7) and nonspecific (lanes8 to17)RNAtranscripts. Lane 1, Probe without protein;lane2, probeand proteinwith no
competitor. (B) Competition with specific (lanes 3to8)and nonspecific (lanes9to 14)double-stranded DNAcompetitors. Lane 1, Probe without protein; lane 2, probe and protein without competitor.
has been found to bind. Transcripts of various truncated versions of the HSV-1 sequences in pRB3881 probe have
also been tested forbinding. 5' deletions upto theStyI site boundaswellasthecomplete probe,but additional deletions to the3'-mostSmaI site abolishedthecapacity of the RNA tobind. We have been unabletodeleteanysequencesfrom the 3' end and retain binding ability; linearization of pRB3881 with ThaI yielded a nonbinding transcript shorter by only7nucleotides (nt).The distance between the 3'-most SmaI site and the ThaI site is 87nt,showingthat the smallest fragment capable of binding isatleast this size.
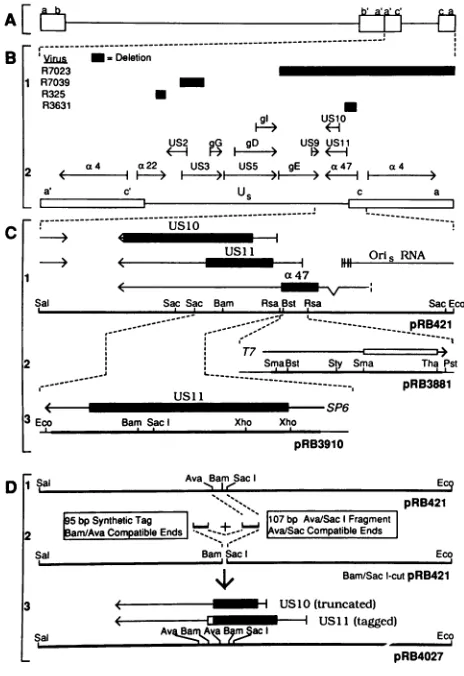
Sequence specificity oftheRNA-binding activitywasalso
demonstrated in competition assays (Fig. 3A). Specific
probe was mixed with various amounts of unlabeled
com-petitor RNAs, either specific (Fig. 3A, lanes 3 to 7) or
nonspecific (Fig. 3A, lanes 8 to 17). The nonspecific
com-petitors used were (i) the T7 transcript of ThaI-linearized pRB3881 and (ii) a 269-nt T7 transcript of the promoter-containing PvuII fragment of pGEM-3Z which also was
nonbindinginthe mobility shiftassay.The salient features of
the resultwereasfollows. First, the specific competitorwas
mosteffective, but allof the competitors decreased binding to some extent, and indeed it appears that the specific
competitor isonly three- tofivefold better than the others. The smalldifferencemaybe deceptive in that onlyafraction
of the 3881 competitoriscompetitive because of secondary structure. The two nonspecific competitors, on the other
hand, formed single, discrete bands in nondenaturing gels and may be completely competitive. If this is taken into
account, the specific competitor was 10- to 20-fold more
effective. Second,alargemassexcessofspecific competitor
(about 300-fold) was required before competition was
ob-served. The reasonfor this is unclear, since labeled probe
was apparently inexcess overbinding activity (i.e., only a
little morethan half of the labeled probe was bound inthe
absence ofcompetitor).
Competitionswere also done with both specific and
non-specific double-stranded DNA to determine whether the activity is RNA specific. Neither the 240-base-pair EcoRI-PstIinsertfragment of pRB3881 (specificcompetitor)northe
379-nt PvuIIfragment of pGEM-3Z (nonspecificcompetitor) competed effectively for the RNA-binding activity (Fig.3B).
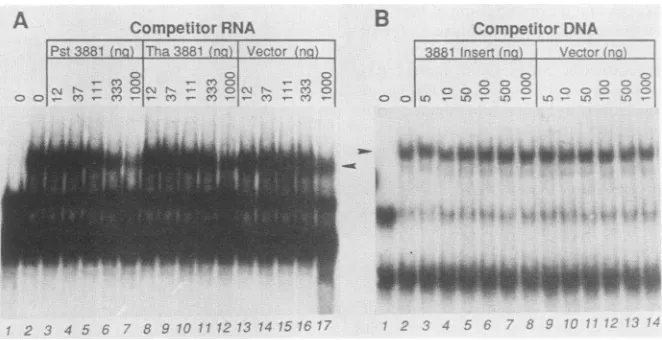
RNA-binding activity is present only in infected cells. Nu-clearextractsfrom infected and uninfectedcells were chro-matographedon aSephacryl S300gelfiltrationcolumn,and column fractionswere assayedforRNA-binding activity by gel mobility shift(Fig. 4). RNA-binding activitywaspresent infractions of infected cellextract (Fig. 4A), peaking near
the void volume and tailing well into the column. This activitywas absent in all fractionsof uninfected cellextract (Fig. 4B), indicating that some function necessary for this activityis either induced orencodedby HSV-1.
AnalysesofRNA-binding activityincrude nuclear extracts. Thegel shiftassay described above didnotpermitassayof RNA-binding activity in crude nuclear extract, since
com-peting nonspecific binding (e.g., by heterogeneous nuclear ribonucleoproteins)masked the shift. We found that itwas
possible toassaytheRNA-binding activity in crude extract by a protection assay as described by Leibold and Munro
(17),inwhich thebindingreactionwasfollowedby digestion
with RNase T1, which destroyed nonspecific complexes. Mixtures ofprobeand either A.5mfraction (Fig. SA,lane4)
orcrude HSV-1-infected cell nuclear extract(Fig. SA,lane 5) resulted in the formation of two complexes between proteinandprotectedRNAwhichwerenotpresenteitherin the absence ofprotein (Fig. SA,lane2)orinthepresenceof
mock-infected cellprotein (Fig. SB, lane 1), confirming that the binding and protecting activity is either induced or
encodedbyHSV. Theamount of theuppercomplex varied
from one experiment to the next, and it was occasionally
absent(e.g., Fig. SB,lane3). Bandsseen evenintheabsence
ofprotein presumably resulted from self-protection of the probe due to secondary structure. HSV-2(G)-infected cell extracts also reactedwith the RNA probe and yielded two specific complexes (aswithHSV-1, only the faster-migrating complex was invariably present) which migrated slightly moreslowlythan their HSV-1counterparts(Fig.SB,lane 2). RNA-binding activity requiresthe product of theU1llopen
readingframe. Inpreliminary experiments,HSV-1 x HSV-2 intertypic recombinants were used to map the viral gene
responsible for the mobility difference between the HSV-1 and HSV-2 complexes to the unique sequences of the S
component. We thentested four deletion mutations ingenes
mappingin the S component (Fig. 1B). Nuclearextracts of
A
J. VIROL.
i
-0
31- 'L, A&MAAn
W**oioiow*WF,Iwwio:w
K
on November 10, 2019 by guest
http://jvi.asm.org/
HSV RNA-BINDING ACTIVITY 3467
A A HSV-1
SephacrylS300 Fraction
to NDC-e
**
O-N.
-%t*_w*F**X*****sX
fn v r, co m,
Oj~~~~~~~~.).
-n,rXa,SNC .co v ' coQKNB
00 LL
E E.0 COC'
( LE E >
eC>
°'N>_ED
tLB Mock
SephacrylS300 Fraction
;;'4C,Lt CS rS e , =
[image:5.612.64.297.76.533.2]<lCvvesrccrvvv- n_ ;,,
FIG. 4. Autoradiographic images of complexes formed between
pRB3881 T7 transcript probe and Sephacryl S300 fractions of HSV-1(F)-infected cell nuclear extract (A) or mock-infected cell
nuclearextract(B). The position of RNA-protein complex migration
wasdetermined by running acomplexformed between the probe
and the A.5mfraction(markedwitharrowheads)inpanelsAandB
(lanes labeled A.5m). PanelB, lane S300,showscomplexes formed
between probe and protein pooled from fractions 39to 49 of the
fractionationdepictedinpanelA.
two of these, mutants R7023 (Fig. 5A, lane 7) and R3630 (Fig. SB,lane4),failedtobindthe RNAprobe.Thecommon
property of both mutants is the absence of(x47 and Us11
openreadingframes.
To determine which of these two open readingframes is required,wetookadvantageof the observationthat
Usll,
aY2gene, requiresfor itsexpressionviralDNAsynthesisand
is not expressed in the presence ofphosphonoacetate, an
inhibitor ofDNAsynthesis, whereas x47 isexpressedin the
presenceof this inhibitor. Sinceextractsof cells infected and
C z
z-+ E:Uu iL 'C E
o c o+ 1:_
I) o: ti U)(
Z j I
^
i!
42 -1
D
t--
r(> C-)
-n
*Wu .
1 2 3 4 5 6 7 8 1 2 3 4 5
FIG. 5. Autoradiographic images of complexes formed between pRB3881 T7 transcript probe and various protein samples in gel shift (panelA, lanes 1 and 3) and RNaseT, protectionassays (all other lanes).Thepresence of protein or state of infection of the cells from whichextractwas prepared is indicated above each lane. The genes deletedinviralmutants R325,R7023, R7039, and R3631 are shown in Fig. 1B. Viral mutant R3630carries the same deletion as R3631 but is tk negative. (A) Positions of the HSV-1-specific protected complexesareindicatedbyarrowheads. Note that although all lanes arefromthe same autoradiograph, only lanes 1 to 4 were adjacent in the original. (B)Positionsof the HSV-1- and HSV-2-specific com-plexes areindicated byarrowheadsmarked 1 and 2, respectively. The lanes were not adjacent in the original autoradiograph. PAA, Phosphonoacetic acid. (C)Lane 1, no protein; lane 2, protein from rabbitreticulocyte lysatein which noexogenousmRNA was trans-lated; lane 3, protein from rabbit reticulocyte lysate in which pRB3910 SP6-transcribed Us11 mRNA was translated; lane 4, protein fromHSV-1(F)-infectedcell nuclear extract. Thepositionof thebinding complex is indicatedwith an arrowhead. Thelaneswere not adjacent in the original autoradiograph. (D) Comparison of complexes formed in extracts of HSV-1(F)- and R4027-infected cells. Positions oftheHSV-1(F)-andR4027-specificcomplexesare indicated byarrowheads.
maintained in the presence of phosphonoacetate didnotbind the RNA probe (Fig. 5B, lane 5), we conclude that the
RNA-binding activity requires
Usll.
The
Usll
open reading frame product is the only viral protein required for RNA-binding activity. Invitrotranscrip-tion-translation was used to demonstrate that the
Usll
functionis theonlyviral functionrequiredfor the formation of the RNA-protein complex. In vitro transcription of EcoRI-linearized pRB3910 (Fig. 1C) with SP6 polymerase produced a710-nt mRNA. When translated in rabbit reticu-locyte lysate, thismRNAproducedaproteinwhichmigrated
in denaturing polyacrylamide gel electrophoresis as a dou-blet with an Mr of approximately 22,000 to
23,000
(not shown), as expected forUsll
protein (13).
In an initialexperiment,incubationof
binding
probewithlysate
in whichUsll
had been translated didnot result in the formation of anyspecific complex.Thisprovedtobe dueto aninhibitory
factor in the translation
mix,
sincemixing
the crudetrans-VOL.64, 1990
*4 .4
$-.4.;'Al
;?, -T .1 I..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.326.555.76.322.2]3468 ROLLER AND ROIZMAN
lation product with HSV-1-infected cell extract also pre-vented complexformation. Passage of the translation mix-ture over a Sephadex G-50 spin column in binding buffer removed theinhibitory factor and resulted in complex for-mation with the RNA probe (Fig. SC, lane 3). Complexes
were notformed with protein fromlysate in which
nothing
(Fig. SC, lane 2) or an irrelevant mRNA (either Brome mosaic virus RNA or synthetic HSV-1 aTIF mRNA; not shown) had beentranslated.
The
Usll
open reading frame product participates in the RNA-binding complex. To determinewhether theUs11
pro-teinparticipates in thebinding complex,weinserted into the 3'terminus ofthecoding domainof
Usll
asequencecodinga15-amino-acid peptide in framewith the
coding
sequenceofthegene, asillustratedinFig. 1D. Thechimericgene,cloned aspRB4027, was recombined into the genomeof
recombi-nant
R3630
in which the a-47 andportions oftheUs11
openreading frame were replacedby thethymidine kinasegene.
Previous studies have shown that the 15-amino-acid se-quencecontainsanepitope foramonoclonalantibodyH1091 to ICP4 (12).
Therecombinant virus (R4027)selectedfor thethymidine kinaseminus phenotypeexpressed aproteinwhichmigrated
more slowly in denaturing polyacrylamide gels than the authentic protein, and, moreover, after transfer to a
nitro-cellulose sheet, the protein reacted with the monoclonal
antibody H1091 (not shown). Extracts ofcellsinfected with R4027bound to the RNA(Fig. SD, lane2), and, moreover, the mobilities of the T1-resistant complexes were slightly
less than those of complexes fromHSV-1(F)-infected cells, consistent with the slightly larger size of the tagged
Usll
protein. This strongly suggests that the RNA-protein
com-plex contains
Usll
protein.RNA-binding activity protects a large piece of the probe from RNase digestion. Thedeletionstudies described above
suggestedthat the minimum RNAprobesize wasin excess of 87nt. Itseemedunlikelythateach of these 87or more nt isrequired for recognitionby
Usll
and,giventhesize oftheUsll
protein, it seemed doubtful thatthis entire sequence could be in contact with theprotein. Totestthelatteridea,wedetermined the size of the RNA fragment protected from RNase T1 digestion by HSV-1 and HSV-2 RNA-binding
activity. The RNA probe was incubated with
HSV-1(F)-infected cell nuclear extract and HSV-2(G)-infected cell
nuclearextract,digested with RNase T1,and
electrophoret-icallyseparated in a nondenaturing polyacrylamide gel along with undigested probe-A.5m protein complex (Fig. 2B).
Binding complex bands (Fig. 2B; indicated by arrowheads) were excised from the gel, and RNA was purified and
subjected to electrophoresis on an 8% denaturing
urea-polyacrylamide gel along with intact probe and molecular
weightstandards. The results of RNase T1digestions (com-pareFig. 2B, lane 2, with lanes 3 and 4) indicated that HSV-1 and HSV-2 RNA-binding activities protected fragments of
identicalsize and that the fragments were 95 to 100 nt in size.
DISCUSSION
Our studies indicate that
Usll
encodes a protein whichparticipatesin and is necessary for sequence-specific binding to anin vitro RNA transcript of an HSV-1 DNA sequence.
Severalaspects of these results require further comment. The
Usll
openreading frame encodes a protein predicted to have a translated Mr of approximately 17,800.The,
pre-dictedaminoacidcompositionis that of an extremely basic
proteinwith apredicted pl equal to 11.1(by the method of
Silleroand Ribeiro
[39]).
Itsstructure is unusual in that theC-terminalhalf of the
protein
iscomposed
of tandem reiter-ations of the sequenceArg-X-Pro.
The number of these repeats varies(33,42)
andprobably
accountsfor the differ-encein the sizes ofUsll
proteins
of different HSV-1 strains(20). Wewere
perhaps
fortunate inourchoice of HSV-1 and HSV-2strainsforintertypic mapping
inasmuchasHSV-1(F)
specifies
arelatively
smallUsll
(20)
andcomplexes
fromstrains
specifying
alarger protein might
notbedistinguish-ablefromthose formed
by HSV-2(G).
The
relatively large
size of theprobe fragment protected
fromRNase
digestion by
thebinding complex (95
to 100nt)
may suggestthatthe
binding complex
containsmorethanasingle
moleculeofUsll
andperhaps
contains otherproteins.
Theobservation thatasmall
change
in the size of theUsll
polypeptide produced
asignificant mobility
shift in thecomplex, however,
suggests thatUsll
islikely
to be themajor
protein
constituent ofthecomplex.
Thechromato-graphic
properties
of theRNA-binding activity
onSephacryl
S300(activity peaks
in the voidvolume)
werecharacteristicofa
protein
muchlarger
than thatpredicted
from the size ofthe
Usll
openreading
frame.However,
the nuclearextractcould be fractionated with ammonium sulfate such that the
RNA-binding
activity
ran as adiscrete,
lower-molecular-weight peak
inSephacryl
S300 columns(work
inprogress).
These results and the observation that the
RNA-protein
complex
wasonly
minimally
retarded relative to that ofunbound RNA suggest that
(i)
Usll
existsininfectedcells inlarge
aggregateswhichreadily
dissociateintomonomers orsmall multimers and
(ii)
theU.11
containedinRNA-protein
complexes
consists ofaspecific
but smallnumberofmono-mers.
Thefunction ofthe
Usll
openreading
frame inthe viralreproductive cycle
isunknown. MacLeanetal.(21)
reported
that the
protein product
oftheUsll
gene binds to DNA cellulose and is elutedonly
with ahigh
saltconcentration,
consistent with a role as a
DNA-binding protein.
It has further beensuggested (21)
thattheUsll
protein
is theMr
21,000
to22,000
-yprotein reported
to interactspecifically
with the HSVasequence(6). Thissequenceis locatedatthe terminiof the S and L components ofHSV DNA
(41)
and containsseveralcis-acting
sites, including
thoserequired
forcleavage
of concatemers of viral DNA intounit-length
molecules and
packaging
ofthe viral genome(reviewed
inreference 34). Several lines of evidence suggestthat
Usll
doesnotplay
arole incleavage
andpackaging
anddoesnot,in
fact,
bind viral DNA in vivo.(i)
TheUsll
openreading
framecan be deletedfrom the genomewith no apparentill effects for virus
growth
incell culture(18, 23).
(ii)
Adetailedstudy
of thefactors which bindtothe HSV-1asequence(4) showed no evidence ofbinding
ofUsll
protein.
(iii)
Anti-body
localizationexperiments
(21) show thatUs1l
protein
isdistributed
throughout
both the nucleus and thecytoplasmof infected cells and is concentrated in the nucleolus. Thisdistribution is more consistent with
affinity
for RNA thanwith
affinity
for DNA inasmuch as viral DNA is excludedfrom thenucleolus(30, 32)and incytoplasmispresentonly
in
capsids
(34). It is not, however, surprising that anex-tremely
basic protein such asUsll
might bindnonspecifi-cally
to DNA, or any other polyanion for that matter, invitro.
Thesecondarystructurespecificityof
Usll
binding mighthave oneoftwogeneralcauses. It may be that the binding
site canbe recognized only insingle-stranded form, and in
nonbinding
RNAisomers this site existsatleast partlyinastem-loop
structure.Alternatively,
binding byUsll
mayJ.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HSV RNA-BINDING ACTIVITY 3469
require the presence of a particular secondary structure element not present in the nonbinding forms of the probe. It is formally possible that
Usll
recognizes some secondary RNA structure element(s) regardless of its sequence.The probe to which
Usll
specifically bound in these experiments is complementary to the a47 andUsll
mRNAs. The sequences protected by the RNA-binding activity are not present in any of the previously mapped transcription units, and sensitive RNAse protection assays on infected cell RNA failed to detect a corresponding transcript (work in progress). These observations suggest two mutually exclu-sive alternatives. First, theU.11
RNA-binding activity may act as a negative regulator of RNA sequences to which it binds, and the probe sequences may be transcribed but fail to accumulate in significant amounts in cells which expressUsll.
To test this hypothesis, we must construct a virus in which theUsll
RNA-binding activity is not expressed but in which the probe sequences and surrounding sequences are not grossly disrupted. None of theUs11-
deletion mutants meet this requirement.Second, the probe sequences used here may not be transcribed in vivo and are merely a substrate analog. While this is an attractive hypothesis, the observation that the 5' transcribed noncoding domain of the gene contains the target sequences of the gene product is in fact too much of a coincidence. To probe this hypothesis further, wehave RNA affinity studies for the
Usll
RNA-binding activity in prog-ress.The function of the product of the
Usll
open reading frame is not known, but several possible functions which could result from binding of RNA late in infection have been excluded. Specifically, (i) host protein synthesis is decreased grossly late in infection bya viral function other than that of the vhs gene (encoded by the UL41 open reading frame), which degrades RNA upon entry of the virusintothe cell(8, 15, 31).Usll
is not involved in the late degradation of host RNA inasmuch as the cells infected with the wild-type parent and the(x47-Usll deletion mutant R3631 accumulate equivalent amounts of cellular(P-actin)
and viral(aTIF and glycoprotein C) mRNAs (data not shown). (ii) HSV encodes several transcripts, including those encoding the single-stranded DNA-binding protein (UL29) (9), the UL38 open reading frame (2), and possibly thymidine kinase (9), which terminate at more than one polyadenylation signal, and HSV-1 encodes or induces a factor which increases the efficiency with which at least one of these signals is used (26). ThatUsll
is not a necessary component ofthis factor is suggested by the equivalent use ofthe UL29 and UL38 polyadenylation signals in HSV-1(F)- and R3631-infected cells (work in progress). (iii) Immunohistochemical studies (21) showed thatUsll
is associatedwith thenucleolus ofthe infected cell, suggesting a possible association with ribo-somes. HSV infection has a profound effect on ribosomal RNA metabolism (42), and a viral protein with sequence-specific affinity for ribosomal RNAcould play a role in the modulation of translation or turnover of specific mRNAs; components of a cellular system which differentially de-grades mRNAs have been found to be ribosome associated (37, 38). While no differences betweencellsinfectedwith the parent and R3631 mutant have been noted, experiments on this point are still in progress.ACKNOWLEDGMENTS
These studies were aidedbyPublic Health Servicegrantsfromthe National Cancer Institute (CA47451) and the National Institute for
AllergyandInfectious Diseases(AI124009 and A11588-11). R.J.R. is apostdoctoral fellow of the American Cancer Society.
LITERATURE CITED
1. Ackermann, M., J. Chou, M. Sarmiento, R. A. Lerner, and B. Roizman. 1986. Identification by antibody toa synthetic peptide of aprotein specified by a diploid gene located in the terminal repeats of the L component of herpes simplex virus genome. J. Virol. 58:843-850.
2. Anderson, K.P., R. J. Frink, G. B. Devi, B. H. Gaylord, R. H. Costa,and E. K.Wagner. 1981. Detailed characterization of the mRNA mapping in theHindIll fragment K region of the herpes simplex virus type 1 genome. J. Virol. 37:1011-1027.
3. Chou, J., and B. Roizman. 1986. The terminal a sequence of the herpes simplex virus genome contains the promoter of a gene located inthe repeat sequences of the L component. J. Virol. 57:629-637.
4. Chou, J., and B. Roizman. 1989. Characterization of DNA sequence-common and sequence-specific proteins binding to cis-acting sites for cleavage of the terminal a sequence of the herpes simplexvirus 1 genome. J. Virol. 63:1059-1068. 5. Chou, J., and B. Roizman. 1990. The herpes simplex virus 1
gene forICP34.5, which maps in inverted repeats, is conserved in several limited-passage isolates but not in strain 17syn+. J. Virol.64:1014-1020.
6. Dalziel, R. G., and H. S. Marsden. 1984. Identification of two herpes simplex virus type 1-induced proteins which interact specifically with the a sequence of herpes simplex virus DNA. J. Gen. Virol. 65:1467-1475.
7. Ejercito, P.M., E. D. Kieff, and B. Roizman. 1968. Characteri-zation of herpes simplex virus strains differing in their effecton social behavior ofinfected cells. J. Gen. Virol. 2:357-364. 8. Fenwick, M. L. 1984. The effects of herpesviruses on cellular
macromolecular synthesis, p. 359-390. In H. Fraenkel-Conrat and R. R. Wagner (ed.), Comprehensive virology, vol. 19. PlenumPublishing Corp., New York.
9. Holland, L. E., R. M. Sandri-Goldin, A. L. Goldin, J. C. Glorioso, and M. Levine. 1984. Transcriptional and genetic analysis of the herpes simplex virus type 1 genome: coordinates 0.29 to 0.45. J. Virol. 49:947-959.
10. Honess, R. W., and B. Roizman. 1974. Regulationofherpesvirus macromolecular synthesis.I. Cascade regulationof the synthe-sis of three groups of viral proteins. J. Virol. 14:8-19. 11. Honess, R. W., and B. Roizman. 1975. Regulationofherpesvirus
macromolecular synthesis: sequential transitionofpolypeptide synthesis requires functional viral polypeptides. Proc. Natl. Acad. Sci. USA72:1276-1280.
12. Hubenthal-Voss, J., R. A. Houghten, L. Pereira, and B. Roiz-man. 1988. Mapping of functional and antigenic domains ofthe a4 protein of herpes simplex virus 1. J. Virol. 62:454-462. 13. Johnson, P. A., C. MacLean, H. S. Marsden,R.G. Dalziel, and
R. D. Everett. 1986. The product ofgeneUs11 of herpes simplex virus type 1 is expressed as a true late gene. J. Gen. Virol. 67:871-883.
14. Kristie, T. M., and B. Roizman. 1986. a4, the majorregulatory protein of herpes simplex virus 1, is stably and specifically associated with promoter-regulatory domains of a genesand of selected other viral genes. Proc. Natl. Acad. Sci. USA 83: 3218-3222.
15. Kwong, A. D., J. A. Kruper, and N. Frenkel. 1988. Herpes simplex virus virion host shutoff function. J. Virol. 62:912-921. 16. Lee, K. A. W., A. Bindereif, and M. R. Green. 1988. A small-scale procedure for preparation ofnuclear extracts that support efficient transcription and pre-mRNA splicing. Gene Anal. Tech. 5:22-31.
17. Leibold, E. A., and H. N. Munro. 1988. Cytoplasmic protein binds in vitro to a highly conserved sequencein the 5' untrans-lated region of ferritin heavy- and light-subunit mRNAs. Proc. Natl. Acad. Sci. USA85:2171-2175.
18. Longnecker, R., and B. Roizman. 1986.Generation ofan invert-ing herpes simplex virus 1 mutant lacking the L-S junction a sequences, an origin of DNA synthesis, and several genes including those specifying glycoprotein E andthea47gene. J.
VOL. 64, 1990
on November 10, 2019 by guest
http://jvi.asm.org/
3470 ROLLER AND ROIZMAN Virol.58:583-591.
19. Longnecker, R., and B. Roizman. 1987. Clustering of genes dispensablefor growth in cell culture in the small componentof the herpes simplexvirus 1genome. Science 236:573-576. 20. Lonsdale, D. M., S. M. Brown, J. H. Subak-Sharpe, K. G.
Warren, and H. Koprowski.1979. Thepolypeptide and theDNA restriction enzyme profiles ofspontaneous isolates of herpes simplex virus type 1fromexplantsof human trigeminal, supe-riorcervicaland vagus ganglia. J. Gen.Virol.43:151-171. 21. MacLean, C. A., F. J. Rixon, and H. S. Marsden. 1987. The
products of gene Usll of herpes simplex virus type 1 are DNA-binding and localize to the nucleoli ofinfected cells. J. Gen. Virol. 68:1921-1937.
22. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual. ColdSpring Harbor Laboratory, ColdSpring Harbor, N.Y.
23. Mavromara-Nazos, P., M. Ackermann, and B. Roizman. 1986. Construction and properties ofaviableherpessimplex virus 1 recombinant lacking the codingsequences of thea47gene. J. Virol.60:807-812.
24. McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan, M.C. Frame, D. McNab, L. J. Perry, J. E.Scott, and P. Taylor. 1988. ThecompleteDNAsequenceofthelong uniqueregionin the genome of herpes simplex virus type 1. J. Gen. Virol. 69:1531-1574.
25. McGeoch, D. J., A. Dolan, S. Donald, and F. J. Rixon. 1986. Sequence determinationandgeneticcontentoftheshortunique region ofthe genome of herpes simplex virus type 1. J. Mol. Biol. 181:1-13.
26. McLauchlan, J., S. Simpson, and J. B. Clements. 1989. Herpes simplex virus induces a processing factor that stimulates poly(A) siteusage. Cell 59:1093-1105.
27. Meignier, B., R. Longnecker, P.Mavromara-Nazos,A. E. Sears, andB.Roizman. 1988. Virulence and establishment of latency by genetically engineered deletion mutants of herpes simplex virus1. Virology 162:251-254.
28. Nielsen, D. A., and D. J. Shapiro. 1986. Preparationof capped RNAtranscripts using T7 RNApolymerase.NucleicAcids Res. 14:5936.
29. Post, L. E., and B. Roizman. 1981. A generalized technique for deletion of specificgenesinlarge genomes: a gene 22 of herpes simplex virus 1isnotessentialforgrowth. Cell 25:227-232. 30. Puvion-Dutilleul, F., E. Pichard, P.Sheldrick,F. Arratric, and
E.Puvion. 1985.Appearance ofhost-specificnucleolar proteins in intranuclear 'dense bodies' following herpes simplex infec-tions. Eur. J. Cell Biol.39:458-468.
31. Read, G. S., and N. Frenkel. 1983. Herpes simplex virus
mutantsdefective in thevirion-associatedshutoff of host
poly-peptide synthesis andexhibiting abnormalsynthesis ofa (im-mediateearly)viralpolypeptides.J. Virol. 46:498-512. 32. Rixon, F. J., M. A. Atkinson, and J. Hay. 1983. Intranuclear
distribution of herpes simplex virus type 2 DNA synthesis: examination by lightandelectron microscopy. J. Gen. Virol. 64:2087-2092.
33. Rixon,F.J.,andD.J.McGeoch. 1984. A 3' co-terminalfamily ofmRNAs from the herpes simplexvirustype1 shortregion: twooverlapping readingframes encode unrelatedpolypeptides one of which has a highly reiterated amino acid sequence. NucleicAcids Res. 12:2473-2587.
34. Roizman,B.,andA. E. Sears. 1990.Herpes simplexviruses and theirreplication, p. 1795-1894. In B. N.Fields, D. M. Knipe, R. M.Chanock,M. S.Hirsch,J. L.Melnick,T.P.Monath,and B. Roizman (ed.), Fields' virology, 2nd ed. Raven Press, Publishers,NewYork.
35. Roizman, B., and P. G. Spear. 1968. Preparation of herpes simplexvirus ofhightiter.J. Virol. 2:83-84.
36. Roller,R.J.,A. L.McCormick,andB. Roizman. 1989. Cellular proteins specificallybindsingle-anddouble-stranded DNA and RNA from the initiation site of a transcript that crosses the originofDNAreplicationofherpes simplexvirus 1. Proc.Natl. Acad. Sci. USA 86:6518-6522.
37. Ross,J., and G. Kobs. 1986. H4 histonemessengerRNAdecay in cell-free extracts initiates at or near the 3' terminus and proceeds3' to 5'.J. Mol. Biol. 188:579-593.
38. Ross,J.,G.Kobs,G.Brewer,and S.W. Peltz. 1987.Properties of theexonucleaseactivitythatdegradesH4histone mRNA.J. Biol. Chem. 262:9374-9381.
39. Sillero, A., and J. M. Ribeiro. 1989. Isoelectric points of proteins: theoretical determination. Anal. Biochem. 179:319-325.
40. Vlazny, D. A., A. Kwong,and N. Frenkel. 1982. Site-specific cleavage/packagingofherpes simplexvirus DNA and the selec-tive maturation of nucleocapsids containing full-length viral DNA.Proc. Natl.Acad. Sci. USA 79:1423-1427.
41. Wadsworth, S., R. J. Jacob, and B. Roizman. 1975. Anatomy of herpes simplex virus DNA.II.Size, composition,and arrange-mentofinverted terminal repetitions. J. Virol.15:1487-1497. 42. Wagner, E. K., and B. Roizman. 1969. Ribonucleic acid
synthe-sis in cells infected with herpes simplex virus. I. Patterns of ribonucleic acid synthesis in productively infected cells. J. Virol.4:36-46.
43. Watson,R.J., and G. F. Vande Woude.1982.DNAsequenceof animmediate-earlygene (IEmRNA-5)ofherpes simplex virus type 1.NucleicAcidsRes.10:979-991.
J. VIROL.