0022-538X/92/010480-09$02.00/0
Copyright X) 1992, American Society forMicrobiology
Functional
Changes
in
Temperature-Sensitive
Mutants
of
the
Adenovirus
Single-Stranded
DNA-Binding Protein
Are Accompanied
by Structural Alterations
MASAYOSHITSUJI ANDGEOFFREY R. KITCHINGMAN*
DepartmentofVirology andMolecularBiology, St. JudeChildren's ResearchHospital,
Memphis,
Tennessee38101-0318
Received5 August1991/Accepted9 October1991
Adenovirus requires the virus-encodedsingle-strandedDNA-binding protein(DBP)toreplicateits DNA.We
have previously shown (M. Tsuji, P. C. van derVliet, and G. R. Kitchingman, J. Biol. Chem.
266:16178-16187, 1991) that theinabilityof threetemperature-sensitive(ts)mutantDBPs
(Ad2+ NDlts23, Ad2ts111A,
andAd5ts125)tosupport DNA replication at thenonpermissive temperaturewasassociated withimpaired ability
to bind to DNA. In thisstudy, we examined thesemutantproteinsfor structural alterations thatmightbelinked to the functional changes. Allthree ts mutants, butnot the wild-type protein, showed different proteolytic cleavage patterns before and after heating at40°C (the nonpermissive temperature), suggesting a possible conformational change during heating. The Ad2+NDlts23 and Ad2ts111A DBPs have single amino acid changes located in a putative zincfingersubdomain(positions282 and280).Inthepresence of zincions,these ts mutants showed significantly increased resistance to inactivation at 40°C.
Surprisingly,
however, thestabilizing effectof zinc was also observed with theAd5ts125 DBP,whichcontainsamutation locatedmorethan
100amino acids from the zincfinger.Other related metalions,such ascobalt,cadmium,andmercury,did not protect the ts DBPs from inactivation at40°C. These resultsindicatethatfunctionalchangesof the ts DBPs in DNAreplication and DNA binding are accompanied by structural alterations in theproteinand thatzinc and
themetal-binding subdomainmayplay animportantrolein thestructureand/or functionofthe DBP.
One of theviralproteins expressed
during
theearly phaseof adenovirus infection isa single-stranded (ss)
DNA-bind-ing protein(DBP). Thisproteinis produced in large
quanti-tiesandis localized predominantly in the nuclei of infected
cells. The DBP performs multiple functions during the
infectious cycle, includingsupportof viralDNAreplication
(reviewed in reference 10), regulation ofviralgene
expres-sion at both the transcriptional (24) and the
posttranscrip-tional (3) levels, determination of host range (14), and
involvement in the virus assemblyprocess(25). Theprotein
binds preferentially to ssDNA with no apparent sequence
specificity and binds to both double-stranded DNA (8, 29,
32) andRNA(1,5, 30). The apparent molecularweightof the
DBP, as estimated by sodium dodecyl sulfate
(SDS)-poly-acrylamide gel electrophoresis (PAGE), is 72,000 (13), but
theprotein is actually composed of529 aminoacids,which
gives a calculated molecular weight of 59,049 (16). This
discrepancy
isprobably dueto ahighdegree ofphosphory-lation (13) andtotheproline-richnature of the protein (16),
leadingtothe anomalous migrationin SDS-containinggels.
Biochemical studies have revealed the presence of two
structurally andfunctionally distinct domainsinthe protein.
Mild chymotrypsin digestion splits the protein into highly
phosphorylated 26-kDa amino-terminal and nonphosphory-lated 44-kDa carboxyl-terminal fragments (13). The
car-boxyl-terminal domain is capable of binding to ssDNA (13)
and RNA (5) and is fully functional in supporting DNA
replication in vitro (2, 9). Genetic studies have identified
amino acid residues important for nuclear localization (21)
andfor hostrange (16) in theamino-terminaldomain of the
protein. Three highly conserved regions (designated CR1,
CR2,
and CR3) within the carboxyl-terminal domain have*Correspondingauthor.
been found by comparative sequence analysis (12) among theDBPs of various human adenovirus serotypes. Some of the mutations introduced in CR2 (amino acids 322 to 330)
andCR3(aminoacids464 to 475) substantiallydecrease the
protein's ability to bind to ssDNA and to support
adeno-associated virus DNA replication in vivo (22, 23) without
affectingits in vivostability, phosphorylation,orsubcellular
localization. Thus, these two regions of the DBP may be directly involved in DNAbinding.
Three temperature-sensitive (ts) adenovirus mutants
(Ad2+ND1ts23, Ad2tslllA, and Ad5tsl25) that are
defec-tive in DNA replication at the nonpermissive temperature have been isolated, identifying two other regions of the carboxy-terminal domain important for the functions of the protein. The Ad2+NDlts23 (17) and Ad2tslllA (27) muta-tions are in closeproximity (a Leu-to-Phe change at amino acid 282 and aGly-to-Val change at amino acid 280, respec-tively), and these amino acids are flanked by -His-X-Cys-and -Cys-X-His-, which has the potential to coordinate a zinc ion (37) in a fashion similar to that found in "zinc finger" proteins (4, 7). These mutant proteins appear to be stable and properly phosphorylated in vivo at the nonper-missive temperature (27, 31). TheAd5tsl25DBP (6), which has a Pro-to-Ser change at amino acid 413 (16), has been used to identify most of the DBP's functions in RNA metabolism, as well as its function in DNA replication. The mutantprotein produced in vivo at the nonpermissive tem-perature ispoorly phosphorylated(19)andrapidlydegraded
(11, 34), suggesting that the mutation is in a region that is
important for theprotein'sstability. Interestingly, however, functionaldefects caused by thismutation can be corrected
phenotypically by secondary mutations at amino acid 347,
352, or 508 (15,26).Collectively,theseDBP mutantsidentify
fourregions importantforproperfunctioningof the protein.
Wehavepreviously demonstratedthat the three ts mutant
480
on November 10, 2019 by guest
http://jvi.asm.org/
TEMPERATURE-SENSITIVE ADENOVIRUS DNA-BINDING PROTEINS 481
DBPs lose the ability to bind to oligonucleotides when
heated at the nonpermissive temperature, suggesting that
their inability to support DNA replication results from
impaired DNA-binding activity (33). A key question that
remained to be answered was whether the mutations are
located in the sites of the protein that directly interact with ssDNA, thereby causing a functional change without altering the overall protein structure, or whether heating at the
nonpermissive temperature induces a structural alteration
that affects DNA binding and perhaps interaction with the
adenovirusDNA polymerase (20). To address this question,
weexamined the proteolytic cleavage patterns of the three ts mutant proteins before and after heating at the nonpermis-sive temperature and found differences that were indicative ofapossible conformational change. We also investigated a
possible role of zinc ion and the putative metal-binding
subdomain (amino acids 273 to 286) inthefunctions of the
DBPbecause boththeAd2+NDlts23 andAd2ts1llA
muta-tions arelocated within thisregion. In thepresence of zinc,
these two mutant proteins, as well as the Ad5tsl25 DBP,
retainDNA-binding activity over significantly extended
pe-riods at the nonpermissive temperature. MATERIALS AND METHODS
DBPs. The wild-type and three ts mutant DBPs were
purified (29) from KB cells infected with AdSdl301,
Ad2+NDlts23, Ad2tslllA,andAdStsl25with the
modifica-tionsdescribed previously (33).
Proteolytic digestions. DBP preparations were diluted to
0.5mg/ml in TEMG buffer(10 mMTris-HCl[pH8.0], 1 mM
EDTA, 2 mM 2-mercaptoethanol [2-ME], 10% glycerol)
containing 100 mM NaCl. These samples were either
di-gesteddirectly orheated to 40°C for 30 min prior to
diges-tion. Reactionswere set up onice by mixing 2
R1
of DBPs (1,ug) and4 ,ul ofproteolytic enzymes (1 ng/,lp) in thebuffers
described below. Tosylsulfonyl phenylalanyl
chlorometh-ylketone-treated trypsin and tosyl lysine chloromethyl
ke-tone-treated chymotrypsin (both from Sigma) digestions
wereperformed in bufferP (40 mMTris-HCI [pH 8.0], 1 mM
EDTA, 2 mM 2-ME, 10%glycerol)containing0.5 MNaClat
30°C for15and 20min,respectively,andstoppedwith 0.5p.1
of100 mM phenylmethylsulfonyl fluoride. Staphylococcus aureus V-8 protease (Pierce) digestions were done in 100
mM phosphate buffer (pH 7.8) at 30°C for 30 min and
terminated by
heating
at90°C
for 5min.Thermolysin
(Sig-ma)digestionsweredone in20 mMTris-HCl(pH8.0-2 mM
CaCl2-1.4
mM 2-ME-5%glycerol
at30°C
for 15 min andstopped with 2 ,ul of50 mM EDTA.
Papain
(Sigma)
diges-tions were performed in buffer P at
30°C
for 15 min andstopped with 1
pl
of50 mM iodoacetamide. Proteinase K(BethesdaResearch Laboratories) digestionswerealso
per-formed inbufferPat
30°C
for15min andstopped
with 0.5 ,ulof100 mM
phenylmethylsulfonyl
fluoride.Proteolytic
frag-ments were separated by SDS-PAGE
(18)
and visualized with the Bio-Rad Silver Stain kit.Gelmobilityshiftassay.Detailsof the
gel
shiftassayusing
an 84-mer DNA have been described
previously (33).
Briefly, the wild-type and ts DBPs, either untreated or
treated as described
below,
were mixed withspecified
amounts of
32P-labeled
84-mer DNA.Samples
were incu-bated on ice for at least 60 min and thenelectrophoresed
througha1%agarose
gel
in TBE(45
mMTris-borate,
1mMEDTA [pH 8.3]). Gels were
partially
dehydrated
andex-posed to Kodak XAR film with Cronex
Lightning-Plus
intensifying screens.
Stabilizing effect of DNA. The DBPs (200 ng) in TEMG buffercontaining 20 mM NaCl and 200
jig
of bovine serum albumin per ml were either heated to 40°C for various periods and then mixed on ice with 5 ng of 32P-labeled 84-mer DNA or mixed first with 5 ng of 32P-labeled 84-mer DNA onice and then incubated at 40°C for various periods. Freeandprotein-bound DNAs were separated at 4°C by thegelmobility shift assay.
Stabilizing effect of antibody. A monoclonal antibody against DBP (B6 antibody in reference 28) was purified from hybridoma culture supernatants by affinity chromatography
by using an anti-mouse immunoglobulin G (IgG)-Sepharose
4B column. Four sets of reactions were carried out with 200 ngof DBPs in TEMG buffer containing 20 mM NaCl and 200 ,ugof bovine serum albumin per ml. In the first, DBPs alone were incubated at 40°C forvarious periods and then mixed with5 ngof
32P-labeled
84-merDNA. In the second and thirdsets of reactions, DBPs were mixed with 200 ng of either
unimmunized mouse IgG (Sigma) or purified B6 IgG, the
mixtureswereheated at40°C for various periods, and 5 ng of
32P-labeled 84-mer DNA was added. In the fourth assay,
DBPs were heated at 40°C for various periods and then
mixed with200 ngofB6IgG and S ngof32P-labeled 84-mer
DNA. Reactions were analyzed at 4°C by the gel mobility
shift assay.
Effects of various metal ions on DNA-binding activity of
DBP. Solutions (15 p.l) containing 250 ng of DBP, 10 mM
Tris-HCl(pH 8.0), 1 mM EDTA, 2 mM 2-ME(omittedwhen
CoS04 was used), 20 mM NaCl, 0.2 mg of bovine serum
albumin per ml, 10% glycerol, and either zinc acetate,
CoS04, CdCl2, HgCl2, MnCl2, or MgCl2 at 2 mM were
incubated at 40°C for the periods indicated in the figures.
Sampleswerechilledonice,and EDTA wasadded to a final
concentration of2.5 mM. DNA-binding activitywas tested
at 4°C with 5 ng of 32P-labeled 84-mer DNA in the gel
mobility shiftassay.
EM. Samples (10 ,ul)for electron microscopy (EM)
con-tained 0.1 ,ug ofM13mpl8ssDNA and 4
jig
ofDBPs(either unheated or heated at 40°C for30 min) in 10 mMTris-HCl(pH 8.0-1 mM EDTA-2 mM 2-ME-20 mM NaCI. The
sampleswereeitherdiluted with90p.l of 50% formamidein
100 mMTris-HCI (pH8.0-10mMEDTA, mixed with 2.5,ul
of 0.5% cytochrome c, and spread on distilled water or
dilutedwith 90 p,l of0.25 Mammoniumacetate, mixed with
2.5
,u1
of 1% cytochrome c, and spread onto a 0.025 Mammonium acetate hypophase. Materials on the surface
were picked up with carbon-coated grids, stained with
alcoholic uranyl acetate, rotary shadowed with
platinum,
and examinedbyaPhilips 301 EM. RESULTS
Proteolytic digestion patterns. We
previously
demon-strated (33) that ts DBPsAd2+NDlts23,
Ad2tslllA,
andAdStsl25 have wild-type
activity
at30°C
in both theoligo-nucleotide-binding and the in vitroDNA
replication
assaysbutirreversibly lose bothactivities
by
heating
at40°C.
Themechanism
underlying
this change could be either a smallstructural change confined to the
DNA-binding
site in theprotein or a
global
conformationalchange
that makes theprotein no longer functional. One method of
distinguishing
between thesetwo
possibilities
isexaminationof themutant DBPs before and afterheating
at40°C
forsusceptibility
toproteolyticenzymes, asthe
accessibility
ofanenzymetoapotential cleavage
site would differas aprotein
undergoes
aconformational
change.
VOL.66, 1992
on November 10, 2019 by guest
http://jvi.asm.org/
Chymotrypsin
Trypsin
V8
protease
M
wt
tslllA ts23 ts125
Hdeating
r- r-, r--r-at
400C 30'
+-97
-66
6-M
wt
tsIllA
ts23
ts125
_ +-_ + -+ -+
M
wt
tslllA
ts23 tsl25
_-+
_-+ _+-_
+no_ W.. - N_ _ - we
,,--a
urn
_
m - -_
so
40 mm
Thermolysin
_ " -"
_m _wo&do
Papain
NW
to 40a
m
so
Proteinose
K
am a_ -_
so
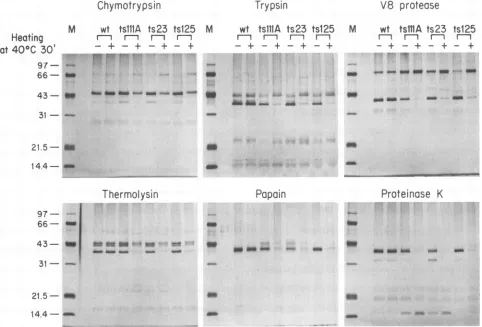
FIG. 1. Proteolytic digestionpatternsofwild-type (wt)and threetsmutantDBPs. The DBPs(1 ,ug,either unheatedorheated at40°Cfor
30min)weredigestedwitha1/250(wt/wt)dilution ofenzymeat30°C,and theproducts wereanalyzed bySDS-PAGE. Lanes Mcontained
markerproteins, whose sizesaregivenonthe left in kilodaltons.
Thewild-type and threemutantDBPs, either unheated or
heatedat40°C for 30min, weredigested separately with six
proteolyticenzymeshavingarangeofsubstratespecificities,
fromhighlyspecific (trypsin and V-8 protease)tononspecific (proteinase K). With each of the six enzymes, all three ts
mutantproteins yieldedatleastonecharacteristic band that was different between the heated and unheated samples, while the wild-type protein produced identical digestion patterns under thetwoconditions (Fig. 1). For chymotryp-sin, V-8 protease, trypsin, and thermolysin, lower-molecu-lar-weight peptide fragments were produced when the un-heated but not heated DBP was used as a substrate,
suggesting thatacleavage sitewasrendered inaccessible by heating. Withpapain and proteinase K, digestion of the DBP to lower-molecular-weight fragments was observed follow-ing heatfollow-ing, indicatfollow-ing increased accessibility to these
en-zymes. Thus, the ts mutants, butnotthe wild-type protein, probably undergo a conformational change at the nonper-missive temperature. This does not necessarily mean that theproteinsaretotally denatured: thetsDBPs thathad been heatedto40°C yielded specificcleavage fragments (Fig. 1), whereas those heated to 90°C for10 min did nothave such specific fragments, presumably because ofa more random conformation (data not shown). The unheated ts mutant
DBPs and those heated at 30°C for 30 min had digestion patterns similartothatof wild-type DBP (datanot shown), suggesting that the mutations do not significantly alter the
proteins' overallstructuresaslongastheyareproducedand kept atthe permissivetemperature. Interestingly, digestion of the unheated ts DBPs with the less specific proteolytic
enzymes papain and proteinase K yielded a predominant cleavage fragmentwithamolecularweightofapproximately 39,000. The production of such a fragment suggests the
presence of a rigid, compactly folded region within the protein, probably corresponding to a part of the carboxyl-terminal domain. Loss of thisfragment with heatingto40°C indicates that the mutations are located at sites that are
crucial for maintenance of the compactly folded structure. Stabilizationofts DBPsby DNA and antibody. Ifa struc-tural change in the mutant proteins is the cause oftheir ts phenotype, interaction of the DBP with other macromol-eculesmight interferewithconformationalchanges, protect-ing the proteins from functional inactivation atthe
nonper-missive temperature. Two macromolecules, an 84-mer ssDNA and an anti-DBP monoclonal antibody (B6), were
chosento test thishypothesis.
The wild-type and three tsDBPs wereheatedat 40°C for various periods and then mixed with 32P-labeled 84-mer DNA orfirst mixed with 32P-labeled 84-merDNA and then incubated at40°C. The DBP-DNA complex and free DNA wereseparatedat4°C byagarosegelelectrophoresis (Fig. 2). The ts DBPs existing free in solution were all rapidly inactivated at 40°C, but the proteins that hadboundto the 84-mer DNAremained associated withthe DNAat40°C for 31
-21.5- II 14.4 -
-97
66 t
43
31
21.5- _m
14.4 -_
amam 6m- MO-am--o
43- IW
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.70.552.82.409.2]TEMPERATURE-SENSITIVE ADENOVIRUS DNA-BINDING PROTEINS 483
A B C
Inc. time r- I
1-i
I lat 400 C 0 5 10 20 10 20 40 60 80 (min)
_ _ .. _ __- _. Al
DBF DNA
- _
__
_ _AmA B - C D E
-,- - IrIC 1
Inc. time
at400C 0 1020 30 40 01020 3040 0 1020 304010203040
(min) t
DNA-DBP-Ab- iI I
DNA-DBP-iJIItS -_
wt
freeDNA-
0
to IIA
DNA-DBP-Ab-- t " DNA-DBP-*
0
free
DNA-
*tea
DNA-DBP-Ab-DNA-DBP- *
seW'.
50d4.
SO
*-
freeDNA- _ M S_IIIr 11 Tl
Inac4tOnC
0 5 10 1520 0 5 10 15 200 5 10 1520 5 10 1520(mnts
f52s
DNA-DBP-Au
-DNA-DBP -
6
0
a
aM_~~~4
fa0
ts125
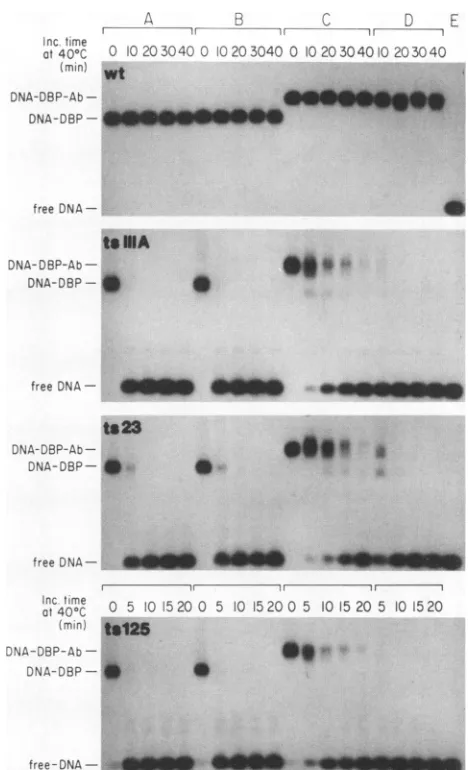
FIG. 2. Gelmobility shift assay showing stabilization of ts DBPs byDNA. The DBPs(200 ng)wereeither incubatedat40°C forthe
indicated periods and mixed with 5 ng of32P-labeled84-mer DNA (A) or mixed with 5 ng of 32P-labeled 84-mer DNA and then incubated at 40°C for the indicated periods (B). Mixtures were
subjected toagel mobility shift assay at4°C. Lanes C contained
32P-labeled84-merDNAonly. wt, wild type; Inc., incubation.
aslongas 80min, indicatingthatbindingto DNArendered
the ssDNA-binding activity of the ts DBPs resistant to
thermalinactivation.
To examinethe effect ofantibody, DBPsalone or mixed
with either unimmunized mouse IgG or purified anti-DBP
antibody were heated at
40°C
forvarious periods and theirDNA-binding activities were examined at 4°C by the gel
mobility shiftassayusing 32P-labeled 84-mer DNA(Fig. 3).
Binding of antibodytothe DBPresultedin afurther shift of
free-DNA-
-
-
-FIG. 3. Gel shift assay showing stabilization of ts DBPs by an anti-DBPantibody (Ab). Four sets of experiments were carried out. DBPs alone were incubated at 40°C for the indicated periods (A). DBPsmixed with unimmunized mouse IgG were heated at 40°C (B). DBPs bound to B6 anti-DBP antibody were heated at 40°C (C). DBPs were incubatedalone at 40°C for the indicated periods and then mixed with B6antibody (D). DNA-binding activity was mea-suredat4°C by the gel mobility shiftassaywith32P-labeled84-mer DNA. Lanes E contained 32P-labeled 84-mer DNA only. Inc., incubation; wt, wild type.
themobility ofthe DNAband. The B6anti-DBP monoclonal
antibody did not interfere with binding of DBP to DNA.
Thus, the epitope recognized by the antibody, which has
beenmapped between amino acids 170and270ofthe DBP
(23), does not appear to be the site that directly interacts
with ssDNA. The ts DBPsbound to the antibody retained
their DNA-binding
activity
at40°C
for significantly longerperiods thandid the unbound DBPs. The
antibody
had noeffectwhen itwas addedtothe ts DBPsafterthey had been
incubatedat40°C for30 min.
EMof DBP-M13 DNA complexes. We
previously
showed that thets DBPs lost theability
to bindtooligonucleotides
after heating at
40°C
but retained somebinding
activity tolarger ssDNA molecules, such as M13 DNA
(33).
Thisobservation led us to
speculate
that the ts DBPs, whenfree DNA
ts
ilA
ts
23
. _
v*a
a
...040000
0
VOL.66, 1992
k
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.318.552.74.459.2] [image:4.612.79.276.74.539.2]J. VIROL.
484 TSUJI AND KITCHINGMAN
v~~ ~ ~ ~7~ ~te2F 1' 4: ' ~ -< , 4.:. ;
4: F -.s
Fy,ff 4
A * S 4:4 . F' ,* R
;S¢j,
S.' o!.1' * CA4F 't
're*
BS*' 44 v Z ts
<d
'SC se.;'<...7.g~';',S-;G,"''...
p; Fb / ;' - 2Wsrik
°
'p0t' - ;'S#Jt W -LW'SS.>'w_,~
*4.~~~~~~~~~~
S~~~~
ttF "ar. i 't '44 ; ' ' '
44 "A~~~~~~~~~~~~~~~~~~~~~~~~~~~~A
St.,.
FIG. 4. Ad5tsl25 DBP-M13 DNAcomplexes observed by EM.(A) M13 DNAonly. (B) A4-,pgsample ofunheatedAd5tsl25DBP plus 0.1 ,ugof M13DNA. (C) A4-,ug sample ofAd5tsl25 DBPheated at40°Cfor30minand then mixed with0.1 pugofM13DNA.
on November 10, 2019 by guest
http://jvi.asm.org/
TEMPERATURE-SENSITIVE ADENOVIRUS DNA-BINDING PROTEINS 485
wt tsIllA ts23 ts125
Inc. time Zn(-) Zn(+) Zn (-) Zn (+) Zn (-) Zn(+) Zn
(-H
Zn (+)at40CC 0 5 10 2030 10 20 40 60 0 5 lO 20
30'0
10 20 30 40 0 5 10 20 30 0 lO 20 30 40 0 5 1020 30 0 10 20 30 40(min)
DBP-DNA-
**'ows
s.free DNA- _rn
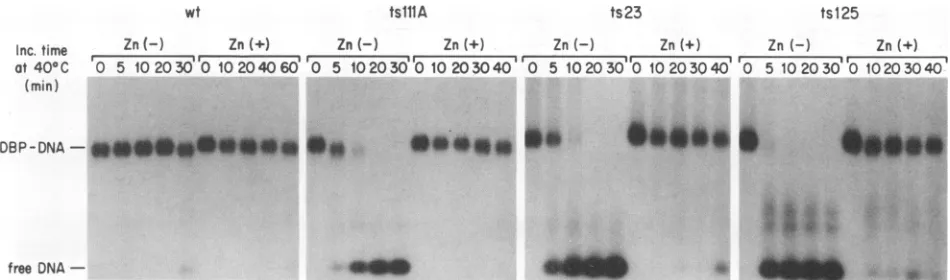
FIG. 5. Stabilizing effect of zinc on the DNA-binding activity of ts DBPs. DBPs (250 ng) were heated at40°Cfor various periods in 10 mM Tris-HCl(pH
8.0)-i
mM EDTA-2 mM 2-ME-20 mM NaCl-200 ,ug of bovine serum albumin per ml-10% glycerol or in the same buffer containing 2 mM zinc acetate, mixed with 5 ng of32P-labeled 84-mer DNA, and electrophoresed at 4°C through a 1% agarose gel. Inc., incubation; wt, wild type.heatedto the nonpermissive temperature, bind to ssDNA in
anaberrant fashion.
To detect any alterations in the DNA-binding mode,
complexeswereformed betweenthe DBPs (either unheated
orheatedat40°C for30min) and M13 ssDNA at a DBP/DNA
(wt/wt) ratio of 40 and examined by EM. The complexes
were spread either from 50% formamide onto water or from 0.25 Mammoniumacetate onto 0.025 M ammonium acetate.
Typicalobservations are presented in Fig. 4. The M13 DNA
moleculeswere readily visualized as open circles (Fig. 4A),
whereas theAd5tsl25DBP-M13 DNA complexes were very
faint andhadslightlydistorted structures (Fig.4B).The poor
contrastmay bethe resultof interference by the DBP with the cytochrome c coating of DNA. Structures similar to
those in Fig.4B were alsoobserved withthe other unheated
ts DBPs and the wild-type protein. Interesting
structures-large, marblelike aggregates-were seen in the complexes
made with theAd5tsl25DBPheated to 40°C and M13 DNA
(Fig.4C). SimilarDNA-proteinaggregates wereformedwith
the heatedAd2+NDlts23 andAd2tslllADBPs but not with
the heatedwild-typeprotein (data not shown). Few globular structures wereobservedwhen theDBPs were heated alone,
so it islikely that theaggregates wereformed followingthe
binding oftsDBPs to DNA. Toexaminethepossibility that
the structures
observed
resulted from the spreadingcondi-tions,wealso spreadthesamples byusing buffers containing
ammoniumacetate instead of formamide. Underthese
con-ditions,M13 DNAs were collapsed and visualizedastightly
wound structures, whereas thecomplexes formed between
the unheated or heated ts DBPs and M13 DNA showed
structures
resembling
those presented inFig.
4B andC,
respectively (datanotshown). Thus,the structuresresulted
from theexperimentaltreatmentandnottheconditions used
for
spreading.
Effects of metal ions on theDNA-binding activity of DBP.
Mutations in Ad2+NDlts23 and Ad2tslllA are located
withinaputative zinc finger subdomain (37),andtheir effects
mayreflect thefunction of this subdomain. Previous results
(27, 33) and those presented above have shown that the
Ad2+NDlts23 mutation, atthe
permissive
temperature,re-sults in some decrease in
DNA-binding affinity
withoutchanging the overall
protein
structure,indicating
that theputative
zincfinger
isdirectly
involvedin DNAbinding.
Toinvestigate this possibility, the
wild-type
and mutantDBPswere heated at40°C inthe presence ofzinc ionsorEDTA
and their DNA-binding activities were tested by a gel
mo-bilityshift assayusingthe32P-labeled 84-mer oligonucleotide
(Fig. 5). Surprisingly, in the presence of zinc, all of the
mutantproteins retained DNA-bindingactivity at 40°C over
extended periods whereasall ofthe ts DBPs were quickly
inactivated at 40°C in the presence of EDTA. Addition of
zinc to the ts DBPs that had already been heated at 40°C,
however, didnot restoreDNA-bindingactivity. The intrinsic
zinc ion appeared to be bound very tightly in the wild-type
and mutantproteins, because EDTA up to 100 mM had no
effect on the DNA-binding activity ofany ofthe DBPs at
30°C(Fig. 5).
To test whether the stabilizing effect is specific for zinc,
similar experiments were done by usingthe closely related
metalions
Cd2",
Hg2+, andCo2"
and unrelated ions such asMn2"
andMg2"
(Fig. 6).Noneoftheseions protectedthe tsDBPs frominactivationat40°C. Infact, both the wild-type
and mutant proteins lost DNA-binding activity after brief
incubation withCdCl2 at30°CorHgCl2at0°C (Fig.7). The
results imply that cadmium and mercury, owing to their
atomic similarity to zinc, readily substitute for zinc in the
protein, but that these atoms may betoo largeto formthe
exact "finger" structurethatmaybe essential for
function-ing ofthe DBP. The Hg ion also has the ability to modify
sulfhydryl groups in the protein, which may contribute to
inactivation ofthe DBP.
DISCUSSION
This work demonstratesthat thefunctionalchangesin the
Ad2+NDlts23, Ad2tslllA, and Ad5tsl25 ts DBPs at the
nonpermissive temperature can be ascribed to structural
alterations in the
proteins.
The structural alteration wassufficienttochange the
accessibility
ofproteolytic
enzymestotheircleavage sites. However, theactualchanges appear
to be minimal-the unheated and heated
proteins
shared many bands whentheyweredigestedwithrelativelyspecific
proteases
(trypsin,
chymotrypsin,
andthermolysin),
and such bands were not obtained with thetotally
denatured proteins.Our EM studies suggest that the mutationscause
changes
notonly in proteinstructure butalso in the self-association
ofthe DBP. The DBP self-associates intwodifferent
situa-tions: oligomer formation in solution and
cooperative
bind-ingtoDNA.Studiesinotherlaboratories
(29)
have indicated"i"m
AMIL.- 1:0
VOL.66, 1992
AL,mLA&A&
IW 0
16
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.71.546.73.213.2]486 TSUJI AND KITCHINGMAN
CdCI2
000
15°0
3000
Inc. time r -
r---C---
00r-(men.)
0 6012018010
2040 60 5 10 15 20DBP-PDN-D-NSA-S
free DNA
Cd2+
Cd2
F ----I
_ cs
In
U-- =
Hg
2+U-)
iSw CD to
3. 4 s
co2+
N3 U-)I
_ rl CNJ
= CM _ : cn en c
a
[email protected]~ 4 e4i,4
HgCI2
6
~000
1500
3000
Inc.
time
,
C 3(min.)
0 5
10
20
40 5 10 20 40
5
10 15
DBP-DNA-S*
free DNA -
=U
_
I___
FIG. 6. Effects of various metalionsontheDNA-binding
activ-ity of DBPs. DBPs (200 ng)wereincubatedat40°Cfor 30mininthe
presence 1 mM EDTA alone or with 2 mM zinc acetate, CdCI2, HgC12, CoS04,MnCl2,orMgCJ2. Theywerethen mixed with 4ngof
32P-labeled 84-mer DNA andelectrophoresed througha1%agarose
gelat4°C. wt,wildtype.
that the wild-type DBP probably exists as a trimer in solution, although itcanbefound in various otheroligomeric forms, depending on the concentrations ofthe protein and
the salt in solution. Cooperative DNA binding is a charac-teristic commonto many prokaryotic and eukaryotic DBPs
that are involved in DNA replication, including the adeno-virus DBP. Complexes formed between thets DBPs heated at 40°C and M13 DNA contained large, marblelike
aggre-gates.Theseaggregatesmayresult fromanaberrationinthe DBP-DBPinteraction involved in cooperative DNA binding because they appeared to be formed in association with bindingto DNA.
The in vitro studies described here and those previously published (33) demonstrate that the structural and functional changes in thetsDBPsatthenonpermissivetemperatureare irreversible. Thisfinding is incontrast tothose of previous in vivo studies (11, 35), in which viral DNA synthesis in Ad5tsl25-infected cells was blocked at the nonpermissive temperature but this effectwas readily reversed by a shift downtothepermissivetemperature,evenin thepresenceof cycloheximide. Ifaggregates such as those seen here are formed in vivo, theremust bea mechanism for disaggrega-tion ofthe DBP in vivo but notin vitro.
Are the mutations that cause the temperature sensitivity phenotype located in functional or structural amino acid residues?Functional mutations havebeen defined as
muta-tion ofanamino acid residue in the protein causingachange in function without altering the overall protein structure.
Structural mutations are defined as those that affect the stability, and hence function, ofaprotein (36). Our findings
FIG. 7. Inactivation ofwild-typeDBPbycadmiumandmercury.
Wild-type DBP (250 ng)wasincubated for variousperiodsat0, 15,
or30°C in the presence of 1 mMEDTA, 2 mM 2-ME, andeither CdCl2orHgCl2at2 mM. Afterincubation,EDTA anddithiothreitol
wereaddedtothesamplesat4°Ctoafinal concentration of 2 mM
each andDNA-bindingactivitywastestedbythegel mobilityshift
assaywith5ngof32P-labeled84-mer. Inc., incubation.
do notfitprecisely with this categorization. The mutations do not lead to global conformational changes but to more
subtle alterationsresultingin changesin the protease
cleav-age patterns. Previous in vivo studies suggested that the Ad5tsl25 DBP might be degraded at the nonpermissive temperature (11, 34), but our in vitro and EM studies indicate that loss of the epitope recognized by the antibody used for immunoprecipitation in the studies ofGinsberg et al. (11) is a more likely explanation. This loss could be causedby the small conformational changeorby aggregation
of the DBP. The Ad2+NDlts23 DBP is apparently stablein vivo, and this appears to be a functional mutation on the basis of thecriteria cited above.However,wedonotbelieve that the mutations are in amino acid residues directly in-volvedin DNAbinding but think, instead, that both the zinc finger region and the region defined by the Ad5tsl25 muta-tionareimportant for the conformation of the DNA-binding pocket. In support of this possibility, studies by Vos etal. (38) demonstrated that the DBP produced by an in vitro translation systemwasfunctional only when itwas synthe-sized in the presenceof zinc.
Functionalinactivation of thetsDBPs invitro by. heating canbeprevented by prior association with either ssDNAor antibody. Stabilization of the DBP by bindingtossDNAis in agreement with the results ofin vivoexperiments showing thatoneround of DNAreplication canoccurfollowing shift
up to the nonpermissive temperature in Ad5tsl25-infected cells(11, 34). As the DBPsarereleasedfrom the ssDNAby
OOmM EDTA
(300C
30min)
=-< N
*-="cn _)c
3 .tb sn
-No metcl Ions
l =
ca
cwvx
u!2-1n-Mg
2_ Nq N <U)_- n) U)
Mn2+
U) U) U)
<z r) U')
= C\M CM
- vn _)
*s_* * __ * *
Zn2+
Zn2
1 to
N1
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.67.299.75.340.2] [image:7.612.337.540.76.358.2]TEMPERATURE-SENSITIVE ADENOVIRUS DNA-BINDING PROTEINS 487
displacement by the elongating second strand, the molecules
becomefunctionally inactivated.
Thepresenceofapotential metal-binding site inthe DBP
was originally suggested by comparative sequence analysis
(37) of the DBPs from distantly related human adenovirus
serotypes. A consensus sequence of the region is
-His-X-Cys-(X)3-Glu-Gly-(X)3-Cys-X-His-, which is similarbut not
identicaltothe typical zinc finger motif(4, 7)found inmany
sequence-specific double-stranded DBPs. Both the Ad2+
NDlts23 and Ad2tslllA mutations are located around the
middle of this sequence, and theamino acid residues
substi-tutedbythese mutationsarehighly conserved among seven
different adenovirus serotypes (37). A key question is
whether this zinc fingerlikeregion is directlyinvolved in the
interaction with DNA or serves as a structural core for
protein folding. Although our results do not clearly
distin-guish betweenthese twopossibilities,we favorthelatter, in
view ofstudies by Vos et al. (38), whichdemonstrated that
the DBP produced by an in vitro translation system was
functional only when it was synthesized in the presence of
zinc.
In summary, we have demonstrated that the
Ad2+ND1
ts23, Ad2tslllA, and Ad5tsl25 ts mutants have an altered
protein structure at the nonpermissive temperature. The
structural alteration is apparently linked to the functional change of these ts DBPs in DNA binding and in DNA
replication. The ts DBPs bound to ssDNA or antibody are
more resistantto inactivation at the nonpermissive
temper-ature than are the free proteins. We have also presented
evidence suggesting potential significance of zinc and the
putative metal-binding subdomain for the structure and/or
function ofthe DBP.
ACKNOWLEDGMENTS
Wethank J.Sublett and P. Stow forhelp with tissueculture, G. Murti for performing the EM studies, and Christy Wright for editorial assistance.
This workwassupported bytheNational Institutes of Health and
theAmerican Lebanese andSyrian Associated Charities. REFERENCES
1. Adam, S. A., and G. Dreyfuss. 1987. Adenovirus proteins associated with mRNAand hnRNA in infectedHeLacells. J. Virol. 61:3276-3283.
2. Ariga, H.,H. Klein, A. J. Levine, and M.S. Horwitz. 1980. A
cleavage product of the adenovirus DNA binding protein is active in DNAreplicationinvitro. Virology101:307-310. 3. Babich, A., and J. R. Nevins. 1981. The stability of early
adenovirusmRNAiscontrolledbytheviral 72 kd
DNA-binding
protein. Cell 26:371-379.
4. Berg, J.M. 1990.Zincfingersand othermetal-bindingdomains. J. Biol. Chem.265:6513-6516.
5. Cleghon, V. G., and D. F. Klessig. 1986. Association of the adenovirusDNA-bindingproteinwithRNAboth invitro and in vivo. Proc. Natl.Acad. Sci. USA 83:8947-8951.
6. Ensinger, M. J., and H. S. Ginsberg. 1972. Selection and preliminary characterization oftemperature-sensitive mutants
of type 5adenovirus. J. Virol. 10:328-339.
7. Evans,R.M.,andS. M. Hollenberg.1988. Zincfingers: gilt by association. Cell52:1-3.
8. Fowlkes, D. M., S. T. Lord, T. Linne, U. Pettersson, and L. Philipson. 1979. Interaction between the adenovirus DNA-bindingprotein anddouble-stranded DNA. J. Mol. Biol. 132:
163-180.
9. Friefeld, B.R.,M. D.Krevolin,and M.S. Horwitz. 1983.Effects of the adenovirusHStsl25 and H5ts107DNA
binding
proteins
onDNAreplication invitro.Virology 124:380-389.
10. Friefeld,B.R.,J.H.Lichy,J.Field,R. M.Gronostajski, R. A. Guggenheimer, M. D. Krevolin, K. Nagata, J. Hurwitz, and
M. S. Horwitz. 1983. Invitroreplication ofadenovirus DNA.
Curr.Top. Microbiol. Immunol. 110:221-255.
11. Ginsberg,H.S.,U.Lundholm,and T. Linne. 1977.Adenovirus
DNA-binding proteinin cellsinfected with wild-type 5
adeno-virus and two DNA-minus, temperature-sensitive mutants, H5ts125 andH5tsl49.J. Virol. 23:142-151.
12. Kitchingman,G. R.1985.
Sequence
of theDNA-binding protein
ofahumansubgroup Eadenovirus(type 4):
comparisons
withsubgroup A
(type
12), subgroup B (type 7), andsubgroup
C(type 5).Virology 146:90-101.
13. Klein, H., W. Maltzman, and A. J. Levine. 1979.
Structure-functionrelationshipsof the adenovirus
DNA-binding protein.
J.Biol. Chem. 254:11051-11060.
14. Klessig, D. F., and T. Grodzicker. 1979. Mutations thatallow humanAd2andAdStoexpresslate genes in
monkey
cellsmapin the viral gene
encoding
the 72K DNAbinding protein.
Cell 17:957-%6.15. KruiJer, W., J. C. Nicolas, F. M. A. van Schaik, and J. S.
Sussenbach. 1983. Structure and function of DNA
binding
proteins from revertants of adenovirus type 5mutants with a
temperature-sensitive
DNAreplication. Virology
124:425-433. 16.Kruijer,
W.,F. M. A. vanSchaik,andJ. S. Sussenbach. 1981.Structure and
organization
of the genecoding
for the DNAbinding protein
of adenovirus type 5. Nucleic Acids Res. 9:4439-4457.17.
Kruier,
W.,F. M. A.vanSchaik,andJ. S. Sussenbach. 1982. Nucleotide sequence of the geneencoding
adenovirus type 2DNA
binding
protein.
Nucleic AcidsRes. 10:4493-4500. 18. Laemmli,U. K.1970.Cleavageof structuralproteins during
theassembly
of the head ofbacteriophage
T4. Nature (London) 227:680-685.19. Levinson,A.D.,E. H.Postel,and A.J.Levine.1977.Invivoand
in vitro
phosphorylation
of the adenovirustype 5single-strand-specific DNA-binding protein. Virology
79:144-159.20. Lindenbaum,J. O.,J. Field,andJ.Hurwitz. 1986.The
adeno-virus DNA binding proteinand adenovirus DNA
polymerase
interact to
catalyze
elongation
ofprimed
DNAtemplates.
J.Biol. Chem. 261:10218-10227.
21. Morin, N., C. Delsert, and D. F.
Klessig.
1989. Nuclearlocal-ization of the adenovirus
DNA-binding
protein: requirement
fortwo
signals
andcomplementation during
viral infection. Mol. Cell. Biol. 9:4372-4380.22. Neale, G. A. M., and G. R.
Kitchingman.
1989. Biochemicalanalysis
of adenovirustype 5DNA-binding
protein
mutants.J.Biol. Chem. 264:3153-3159.
23. Neale, G. A. M., and G. R.
Kitchingman.
1990. Conservedregion 3 of the adenovirus type 5
DNA-binding protein
isimportant for interaction with
single-stranded
DNA. J. Virol.64:630-638.
24.
Nevins,
J. R., and J. J. Winkler. 1980.Regulation
ofearly
adenovirus
transcription:
aprotein product
ofearly region
2specifically
repressesregion
4transcription.
Proc. Natl.Acad. Sci. USA 77:1893-1897.25. Nicolas, J. C., P. Sarnow, M. Girard, andA.J. Levine. 1983.
Hostrange
temperature-conditional
mutants in the adenovirusDNA
binding
protein
aredefective in theassembly
of infectious virus.Virology
126:228-239.26. Nicolas, J. C., F. Suarez, A.J. Levine,and M. Girard. 1981.
Temperature-independent
revertantsof adenovirusHSts125andHSts1O7
mutantsinthe DNAbinding
protein:
isolationofa newclassofhost rangetemperatureconditionalrevertants.
Virology
108:521-524.
27. Prelich,G.,and B. W.Stillman. 1986. Functional
characteriza-tionof thermolabile
DNA-binding proteins
thataffectadenovi-rusDNA
replication.
J. Virol. 57:883-892.28. Reich, N. C., P. Sarnow, E.
Duprey,
and A. J. Levine. 1983.Monoclonal antibodies which
recognize
native anddenatured forms ofthe adenovirusDNA-binding
protein. Virology
128: 480-484.29. Schechter, N. M., W. Davies, and C. W. Anderson. 1980.
Adenovirus coded
deoxyribonucleic
acidbinding
protein.
Iso-lation,
physical
properties,
andeffects ofproteolytic
digestion.
Biochemistry
19:2802-2810. VOL.66, 1992on November 10, 2019 by guest
http://jvi.asm.org/
30. Seiberg, M., Y.Aloni, and A. J. Levine. 1989.The adenovirus
type 2 DNA-binding protein interacts with the major late
promoterattenuated RNA. J. Virol.63:1134-1141.
31. Stillman, B. W., E. White, andT. Grodzicker. 1984. Indepen-dentmutations in Ad2tslll causedegradationofcellularDNA
and defective viralDNAreplication. J.Virol. 50:598-605.
32. Stuiver, M. H., and P. C. van der Vliet. 1990. Adenovirus
DNA-binding protein formsamultimericprotein complex with
double-strandedDNAand enhancesbindingofnuclearfactorI.
J. Virol. 65:379-386.
33. Tsuji, M., P. C. van der Vliet, and G. R. Kitchingman. 1991.
Temperature-sensitive mutants of adenovirus single-stranded
DNA-binding protein: inability to support DNA replication is associated withanalteredDNA-binding activity oftheprotein.
J. Biol. Chem.266:16178-16187.
34. van der Vliet, P. C., A. J. Levine, M. J. Ensinger,and H. S.
Ginsberg. 1975.ThermolabileDNA-binding proteinsfrom cells
infected with a temperature-sensitive mutant of adenovirus defective in viral DNAsynthesis. J. Virol. 15:348-354.
35. vanderVliet,P.C.,andJ.S. Sussenbach.1975.Anadenovirus
type 5 gene function required for initiation of viral DNA replication. Virology67:415-426.
36. Vershon,A.K.,J.U.Bowie, T. M. Karplus, and R. T. Sauer. 1986.Isolation andanalysis ofarcrepressormutants: evidence for an unusual mechanism of DNA binding. Proteins Struct. Funct. Genet. 1:302-311.
37. Vos, H.L.,F.M. vanderLee,A. M. C. B.Reemst,A. E.van
Loon,andJ.S.Sussenbach. 1988. Thegenesencoding the DNA binding protein and the 23Kproteaseofadenovirustypes40 and
41.Virology 163:1-10.
38. Vos,H.L.,F.M.vanderLee, andJ.S.Sussenbach. 1988. The
binding of in vitro synthesized adenovirusDNAbindingprotein
tosingle-stranded DNA is stimulated by zinc ions. FEBS Lett. 239:251-254.