JOURNAL P. 487-492 0022-538X/82/110487-06$02.00/0
Copyright01982,American Society forMicrobiology
Vol.44,No.2
Replication
of Mouse
Hepatitis
Virus:
Negative-Stranded
RNA
and
Replicative
Form RNA Are
of
Genome Length
MICHAEL M.C. LAI,* CHRISD. PATTON,ANDSTEPHEN A.STOHLMAN
Departmentof Microbiologyand Departmentof Neurology,* UniversityofSouthernCaliforniaSchool of
Medicine,LosAngeles, California90033
Received 10 May1982/Accepted 16July 1982
Therearesevenvirus-specific mRNA speciesin mousehepatitis virus-infected
cells (Lai etal., J. Virol. 39:823-834, 1981). In this study, weexamined
virus-specific negative-stranded RNA to determine whether there are corresponding
multiplenegative-strandedRNAs. Intracellular RNAfrommousehepatitis
virus-infected cells was separated by agarose gel electrophoresis, transferred to
nitrocellulose membranes, and hybridized to positive-stranded genomic 60S
[32P]RNA. Onlyasingle RNAspecies ofgenomic size wasdetectedunder these
conditions.This RNA was negative stranded. Nonegative-stranded subgenomic
RNA wasdetected. Wealso studieddouble-stranded replicative-formRNAin the
infected cells. Only onereplicative-formof genomic size was detected. When the
double-strandedRNAisolated withoutRNasetreatment wasanalyzed, again only
one RNAspecies of genomic sizewasdetectable.Furthermore,mostof the
virus-specific mRNAs couldbe releasedfrom this RNA species uponheating. These
resultssuggestthatallofthe mousehepatitis virus-specificRNAs aretranscribed
from a singlespeciesofnegative-stranded RNAtemplateofgenomic size.
Mousehepatitis virus (MHV), acoronavirus,
contains a single, positive-stranded genomic
RNAwithamolecularweightof5.4x
106(7,
8).This RNA can be divided into at least seven
genetic regions (5) whichencode three structural
proteins,pp5O,gp23, andgp90/180,andpossibly
severalnonstructuralproteins (11).Theinfected
cells contain six virus-specific subgenomic
mRNAs and a genomic mRNA (2, 5, 9, 13).
These mRNAs have a nested-set structure in
which the sequence of each mRNA is included
withinthe next-larger mRNA. Furthermore, the
sequenceofeach mRNAcorresponds tothe
3'-endofthegenomicRNAand extends into the 5'-endforadistance corresponding tothe size of
eachindividual mRNA species (5).Thus, all of
the mRNAsshare identicalsequences at the
3'-end but presumably contain different 5'-half
sequences. We have also found that all ofthe
mRNAs share at leastfive nucleotidesatthe
5'-endsandthatsomemRNAs contain an
oligonu-cleotide which is not present in the genomic
RNA(6). These datasuggest that the synthesis
of MHV mRNAs involves a complex
mecha-nism, possibly an unusual form of RNA splicing
whichtakesplace in the cytoplasm (6).
Recently, we have detected two RNA
poly-merase activities in MHV-infected cells, one
detected early (1 hpostinfection [p.i.]) and the
other detected late (6 h p.i.). These two
poly-meraseshave different enzymatic requirements
andproperties (1). Furthermore, the RNA
prod-ucts of the early polymerase are of
negative-strand polarity, compared with the genomic
RNA, whereas those of the late polymeraseare
of positive-strand polarity (P. R. Brayton,
M. M. C.Lai, and S. A. Stohlman,unpublished
data). Therefore, MHV RNA replicatesthrough
a negative-stranded RNA intermediate which
serves as the template for the synthesis of
mRNAs. Since multiple subgenomic mRNAs
havebeendetected,it is of interesttodetermine whether the negative-stranded RNA
intermedi-ateisasingle genomicspeciesorincludes
multi-ple subgenomic and genomic species
corre-sponding tothepositive-stranded RNA. In this
study, weexamined thisissuebydirectly
exam-iningthenegative-strandedRNAspeciesand the
double-stranded replicative-form (RF) RNA in
MHV-infectedcells. We found thatonlya
nega-tive-stranded genomicRNAintermediate could
be detected.
MATERIALS ANDMETHODS
Viruses and cells.MHVstrain A59(MHV-A59)was used throughout. In some experiments, strain JHM wasalso used. These two strains have beendescribed previously (8). Virus was grown in DBT cells in Dulbeccominimalessentialmedium(DMEM) supple-mentedwith1% fetal calf serum.
PreparationofIntrcellular virus-specificRNA. Prep-aration ofintracellular[32P]RNAfrom MHV-infected cells has been described (5, 6). Briefly, L-2 cellswere 487
on November 10, 2019 by guest
http://jvi.asm.org/
preincubated with DMEM containing 1/10 the normal concentration of phosphate and 0.5% fetal calf serum for 10 to 12 h before infection. After virus adsorption, the cells were incubated in phosphate-free DMEM containingtwice thenormalconcentrationofvitamins and amino acids, 2 ,ug of actinomycin D per ml, 1% dialyzedfetal calf serum, and 250 ,Ci of32Pi (ICN Pharmaceuticals) per ml. At 7 to 9 h p.i., the cells were chilledonice andlysedwith a buffer containing 10 mM Tris (pH 8.5), 60 mM NaCl, 1 mM EDTA, and 0.5% Triton N-101. Nuclei were removed by centrifugation at1,800xg for 5min. Sodium dodecyl sulfate (SDS) wasadded to the supernatant fluid to a final volume of 1% SDS,and thesupernatantfluid was thenextracted withchloroform-phenol (1:1). The RNA was precip-itatedwith 2 volumes of ethanol.
Forpreparation of unlabeledintracellularRNA, the same protocol wasfollowedexcept that theinfected cellswere grown inunmodifiedDMEM supplemented with 1% fetal calf serum throughout infection.
Preparation ofdouble-sandedRNA. The intracellu-lar [32P]RNAextractedas described above was precip-itated by sedimentation at 15,000 xg for 15min.The RNA was resuspended in NTE buffer (0.1 M NaCl, 0.01 MTris-hydrochloride [pH7.4],0.001 MEDTA). NaClwas added toobtain a finalconcentrationof 2 M, and the solution was left at 0 to4°Cfor 48 h. The RNA was then sedimented at 13,000 rpm for 30min. The supernatantwasdiluted with an equal volume of water and precipitated with 2 volumes of ethanol. For diges-tionwithRNase, the RNA waspelleted at 12,000 rpm for15 minand suspended in low-salt buffer (0.01 M Tris-hydrochloride [pH 7.4], 0.001 M EDTA), and SSC buffer was added to the solution to a final concentrationof 2x SSC (1xSSC is 0.15 MNaCl plus 0.015 Msodium acetate). The RNA was digested with RNase A (20,ug/ml)at37°C for 60 min. The RNA was then extracted with phenol-chloroform (1:1) and pre-cipitatedwith 2volumesof ethanol.
Agarosegelelectrophoresisof RNA.Electrophoretic analysis of RNA was generally performed in 1% agarose gelsmade in REbuffer,pH 8.1(50 mM boric acid,5mMsodiumborate,1 mMEDTA, and 10 mM sodium sulfate (5, 6), except when RNA was to be transferredtonitrocellulose filters (seebelow). Elec-trophoresiswas at90 Vfor5.5 h.After electrophore-sis, the wet gel was wrapped with cellophane and autoradiographed.
Forextraction ofRNAfrom the agarose gel, the agarose fractioncontainingRNAwashomogenizedin abuffer containing0.3 M NaCl, 0.01 M Tris-hydro-chloride [pH7.4],0.001MEDTA,and0.5% SDS and continuously agitatedat roomtemperature for 12 h. After the gel pieces were removed, the RNA was
precipitatedwith 2 volumes of ethanol.
RNA blottingand fflterhybridization. Intracellular RNA was analyzed by a modification of the filter hybridizationmethod of Thomas (15). Briefly, intra-cellular virus-specific RNA was denatured in 1 M
glyoxal-50%odimethylsulfoxide(DMSO)-10mM sodi-umphosphate buffer (pH 6.8)at50°Cfor1h(10).The RNA was then electrophoresed ina1% agarosegel made in 10 mMphosphate buffer(pH6.8) at90 Vfor 5.5 h.Afterelectrophoresis,the RNAwastransferred to nitrocellulose membrane filters by the procedure described by Thomas (15). Hybridization was per-formed in10mlofbuffercontaining50%oformamide,
5x SSC, 50 mM sodium phosphate (pH 6.8),250,ugof sonicated, denatured salmon sperm DNA per ml, 0.02%each bovineserumalbumin, Ficoll, and poly-vinylpyrrolidone,250 ,ugof rRNA perml,10% dextran sulfate, and 5 x 106 cpm of 60S MHV [32P]RNA (specific activity, S x 106 to 10 x 106 cpm/,ug). Hybridizationwasperformedat42°Cfor 3days. The RNAblots were washed with fourchanges of room-temperature 2x SSC containing 0.1% SDS and then with two changes of50°C 0.1x SSCcontaining 0.1% SDS. They were thenexposed to Kodak XRfilm at -70°C with anintensifyingscreen.
RESULTS
Characterization of negative-stranded RNA in
MHV-infected cells. We first characterized the
negative-stranded RNA in MHV-infected cells
by a direct approach. The total cytoplasmic
RNA of MHV-A59-infected L-2 cells was
ex-tracted at 7to 9 hp.i., when the virus-specific
RNA synthesis was at its maximum level (5).
Thetotal RNAwasthendenatured with
DMSO-glyoxalby the procedure of McMasterand
Car-michael(10)andseparated byelectrophoresison a1%agarosegel. Aparallelcontrol experiment with MHV-A59-specific intracellular [32P]RNA analyzed under identical conditions revealed
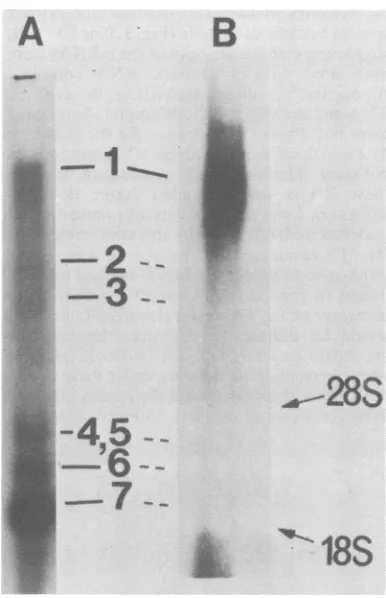
that there were atleast six virus-specific RNA
species (Fig. 1, lane A). These species have been
extensively studied and represent
positive-strandedgenomicandsubgenomic virus-specific
mRNAs (5, 6). The unlabeled RNA was
trans-ferred from the agarose gel to a nitrocellulose
membrane and hybridizedto60S[32P]RNA
ex-tracted from the purified MHV-A59 virion (7).
The viral genomic RNA contains
only
positive-stranded RNA (7, 8); therefore, this 2P-labeled
probewould detectonly negative-strandedRNA
species. Onlyone RNA band, correspondingin
size tothe viriongenomic RNA, was detected
(Fig. 1, lane B). Therewas occasionally some
faint radioactivity in the lower part of the gel;
however, it was not reproducible and did not representdiscrete RNAbands.
Thisobservationsuggeststhat there isonlya single negative-stranded RNA species in the
MHV-infectedcells and that it has
approximate-ly thesamesizeasthatof thegenomic RNA.We
couldnotruleoutthepossibilitythat therewere
very small amounts of negative-stranded
sub-genomic RNA; however, this possibility seems
veryunlikely, sincethe amountof smaller
posi-tive-stranded subgenomic mRNAs far exceeded
that of genomic RNA (Fig. 1, lane A) (4, 9).
Therefore, our data suggest that all the
subge-nomic and genomic mRNAs are transcribed
from a negative-stranded genomic RNA
tem-plate. Since the RNA band detected by this
procedurewasbroad (Fig.1, lane B), wecould not ruleout the presence of several species of
negative-strandedRNAwithverysimilar
molec-ularweights.Alternatively,theheterogeneity of
on November 10, 2019 by guest
http://jvi.asm.org/
MOUSE HEPATITIS VIRUS REPLICATION 489
A
B
--2
--3
--3
-45
--
-'6---7
--FIG. 1. Electrophoretic analysis o
positive- and negative-strandedRNAs cellular[32P]RNA from MHV-infecte' denatured and separatedby electropt
agarose gel. After electrophoresis, tl exposed to Kodakfilm directly. Lan intracellular RNAwasdenatured with
(10) and separated by electrophoresis lane A. After electrophoresis, the R ferredtoanitrocellulose filter andhyl genomic [32P]RNA by publishedproce filterwas washed, dried, andexpose
withanintensifyingscreen.
solublefractionwassmallsingle-stranded RNA
which could not be precipitated with high salt
concentrations. Similar observations have been
madefor the RF RNA of influenza virus(3).The
solublefractionwasdigested with RNase A(20
,ug/ml)
in 0.3 MNaCltoremovecontaminating
single-stranded RNA speciessowecouldstudy
the RF RNA. The resulting RNase-resistant
RNAwasthenanalyzedbyelectrophoresison a
1%agarosegel. This RNAwasasinglespecies
with an electrophoretic mobility only slightly
slowerthan that of the genomic RNA (Fig. 2,
lane A). This RNAwasextractedfrom thegel,
denatured with DMSO-glyoxal (10), and
ana-lyzed by agarose gel electrophoresis. A single
genomic RNA specieswasdetected(Fig. 2, lane
B). The electrophoretic mobility of this RNA
was slightly faster than or sometimes
indistin-ooc guishable from that of the double-stranded RF
_-LO RNA (data not shown). It was unclear which
secondarystructure rendered theseRNAs
elec-trophoretically similar. Atthe bottomof thegel,
there was some faint dispersed radioactivity
which might represent the remaining portions,
after RNase A digestion, of the various RNA
species bound to the genomic RNA (compare
with Fig. 3, lane C; see below). This result
showed thatthedouble-strandedRFRNAin the MHV-infected cells consisted of a single
genomic species. Although it was still possible
f MHV-specific thatsomedouble-stranded subgenomic RFRNA
celnls
wAas
heatmight
have aconfiguration
which rendered itioresis on a1% inseparable from the genomic
RF
RNA underhe wet gel was our electrophoretic condition, this possibility e B, Unlabeled seems quite unlikelyin view of the fact thatthe
glyoxal-DMSO size of the smallest RNA is only-1 thatof the asdescribed for genomic RNA (5, 9, 13). This result, together LNA was trans- with the result obtainedfrom the filter
hybridiza-bridized toviral tionanalysis of negative-stranded RNA,
strong-tuo
Koda)
filmly
suggeststhat thetemplate
RNAfor all of theMHV mRNAs is a
single
negative-stranded
ge-nomic RNA species.
Characterizationofapartfally double-stranded
the RNAscouldhave beenanartifact of
extrac-tion,assimilarheterogeneitywasnoticed for the
positive-stranded genomic RNA(Fig. 1, lane A).
Characterization ofdouble-stranded RF RNA.
To confirm the interpretation derived from the
analysis of negative-stranded RNA, we studied
the double-stranded RF RNA involved in the
synthesis ofMHV mRNA.Virus-specific
intra-cellular [32P]RNA was precipitated with 2 M
NaCl at0to4°C for48h. The double-stranded
RNA in the soluble fraction was found to be
enriched(Table 1). However, 24% of this
frac-tion at most was RNase resistant. This value
was consistently obtained in repeated
experi-ments. The reason for the failure to obtain a
higherpercentageof double-strandedRNAwas
notclear. Presumably,therestof the 2 M
NaCl-TABLE 1. RNaseresistance of various forms of MHVintracellular RNA'
Sourceof RNA resistance% RNase
2MNaCl(soluble) 24
2MNaCl(precipitate) 2
RI RNA(Fig. 2, lane A) 100
RI RNA(Fig. 3, lane B) 61
RNAno. 6(Fig. 3, lane C) 9 RNAno. 7(Fig. 3, lane C) 9
a RNA was dissolved in 0.5 ml ofasolution contain-ing0.3 MNaCl, 0.03 M sodium acetate, 0.01 M Tris-hydrochloride (pH 7.4), 1 mM EDTA, and 20 Fg of RNase Aper ml and incubatedat37°Cfor 60 mnu. The amountof RNase-resistant RNAwas determined by precipitation with trichloroacetic acid.
VOL. 44,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.491.47.240.75.374.2] [image:3.491.250.447.524.629.2]the majority of the MHV-A59-specific mRNA
species became detectable (Fig.3, lane C) (5, 6),
suggesting that the majorityof the mRNAswere
associated with a genomic RNA complex. Among the heat-dissociated RNAs,thegenomic RNA and the subgenomic RNAs no.4and no. 5 were not distinct. Thereasonfor the failure to
detect distinct bands of these RNAspecieswas
notclear. The mostlikely explanation wasthat
these RNAs were degraded during isolation.
RNAsno.1andno.4 wereusuallymoredifficult
todetect undermostoftheanalyticalconditions
(5). This resultsuggeststhatatleastsomeof the
virus-specific mRNAs in MHV-infected cellsare
bound to genomic RNA complex which might
represent anRI.Themolecularweight ofthis RI
would be difficult to determine because the
secondarystructureof the RNA wouldinterfere
with electrophoretic mobility under these
condi-tions. Thisobservation and the results obtained
from the study ofnegative-stranded RNA
sug-A
B
C
FIG. 2. Electrophoretic analysis of double-strand-ed RF RNA. Lane A, Double-stranded RF RNA (arrow) isolated from MHV-infected cells and then digestedwith RNasewasanalyzed byelectrophoresis on1%agarosegelasdescribedin thelegendtoFig.1. LaneB, TheRNA band in lane Awaseluted from the gel,denatured withglyoxal-DMSO, andanalyzed by
agarose gel electrophoresis (arrow) as described for
laneA.
RI. To better understand the RNA replicative intermediate (RI) involved in the synthesis of MHVRNA,wefurtheranalyzed the RNA in the
2 MNaCl-soluble fraction without RNase treat-ment.Anysingle-stranded RNA partially bound
tothe negative-stranded RNA could be
identi-fiedelectrophoretically. This RNA fractionwas
analyzed by agarose gel electrophoresis. There
wasonlyoneRNAspecies detected underthese conditions, and it was in the region of the gel
correspondingtothegenomic RNA (Fig. 3,lane B). This RNA hadamigrationratesimilartothat of RNase-treated RF RNA (Fig. 3, lane A). Again, itwasunclear whichsecondary structure rendered these RNAspecieselectrophoretically indistinguishable. No discrete subgenomicRNA
was detected even when no RNase digestion was performed. When this RNA species was
heatdenaturedandanalyzed byelectrophoresis,
[image:4.491.86.210.79.378.2]-*7
FIG. 3. Electrophoretic analysis ofpartially dou-ble-stranded RI RNA. RNA obtained fromthe 2 M NaCI-soluble fraction without RNase digestion was
analyzed by electrophoresis on a 1% agarose gel. Double-stranded RF RNA (as described for Fig. 2, laneA)wasusedas acontrol(lane A).LaneB,Native RI RNA. LaneC,RI RNAheatedat100°Cfor 1min beforeelectrophoresis.
A
B
_ i
_b
.. _
.
-_ _:
g
w
:^... ..
.:
:.. :: ss s
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.491.303.401.307.599.2]MOUSE
gest that this RI consists of various mRNA
species associatedwith anegative-stranded ge-nomic RNA.
To further understand the structure of the
various RNA species detected in the
MHV-infected cells, we determined the RNase
resist-anceof each RNA species after the species was
elutedfromtheagarose gel. The
double-strand-ed RF RNA wascompletely resistant to RNase
digestion (Table 1). In contrast, RNA isolated
directlyfromthe 2 M NaCl-solublefractionwas
only 61% resistant to RNase. This result
con-firmed thehypothesis that the latter form
repre-sented an RNA RI which contained
single-stranded RNA tails. In contrast, the RNA
species releasedfrom the RIby heat treatment
were sensitive to RNase digestion, consistent
with the idea that they represented
single-stranded RNAspecies.
DISCUSSION
From thestudy of thevirus-specific
negative-stranded RNA and double-negative-stranded RF RNA in
MHV-infected cells, we concluded that there
was only one negative-stranded genomic RNA
speciesand thatthis species servedasthe
tem-plate for the synthesis of multiple
positive-stranded mRNAs. Therefore, thereplication of
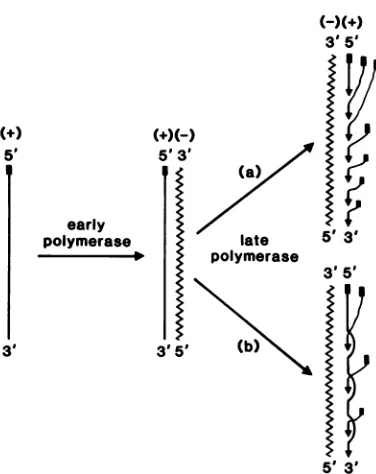
MHV RNA canbe summarized asin
Fig.
4. Inthis scheme, the positive-stranded genomic
RNAfromincomingvirus is firsttranscribedby
the early RNA polymerase (1) into a
negative-stranded genomic RNA. This process
presum-ably involvesfaithfulcopying ofallof the
geno-mic sequences, except for the
3'-polyadenylic
acid sequences. However, no details of the
structureofthe negative-stranded RNA,except
foritssize, arecurrentlyknown. Thenextstep inreplication isthe synthesis ofmultiple
geno-mic and subgenomic mRNAs. This step
appar-ently involves transcription
by
the late RNApolymerase (1) ofthefull-length
negative-strand-ed RNA. This isaverycomplexstep, and very
few details about itarecurrentlyknown.
Tran-scriptional mapping data have shown that the
UV targetsizeofeach mRNA is thesame asits
physical size (4),suggestingthat each mRNAis
transcribed individually at its own initiation
pointrather that beingderived fromagenomic
RNA. Furthermore, as we have shown previ-ously,thesynthesisofsubgenomicmRNAsmay
involve some forms ofRNAprocessing(6). One
of these processes could be involvement ofRNA
splicing (6; unpublished data). Itisnotclear how
and whenRNAprocessingoccursduringmRNA
synthesis.
The second unanswered question is whether
all of the mRNAsaresynthesizedfrom thesame
negative-strandedRNAmolecule. Thefindingin
(i.)
5,
U
early polymerase
5'3'
.10
I
3959 3'
3p5'
5' 3' 35 53
5' 3' FIG. 4. Proposed scheme for MHV RNA replica-tion.This scheme assumes that nascent RNAstrands areimmediately displaced byother newlysynthesized strands, so that there are multiple single-stranded RNA tails in the RI. The shorter positive-stranded RNAtails complementary to the 5'-end of the nega-tive-strandedRNAtemplatein the RI representnewly synthesizedsubgenomic mRNAs. The solid rectangles atthe 5'-ends of the positive-stranded RNAs represent the presumptive leader sequences (6; unpublished data). The number of RNA strands in the RI is arbitrarily assigned. No evidence has been obtained that the nascent positive strands contain the leader sequences. (a) Semiconservative mechanism for mRNAsynthesis; (b)conservative mechanism.
this study that no double-stranded subgenomic
RF RNA was detected after RNase digestion
suggests that the entire negative-strandedRNA
template is protected by binding to mRNAs.
Thus, any singlemolecule of negative-stranded
template RNA is probably used for the synthesis
of more than one mRNA species and also of
genomicRNA(Fig.4,model a). It is notknown,
however, whetherallof themRNAs are
synthe-sized on the same RNAtemplate
simultaneous-ly. Analternative possibility is that the
MHV-specific negative-stranded RNA is always
present as adouble-stranded RNA. In that case,
mRNAS would be synthesized by a
conserva-tive mechanism (Fig. 4,model b) as opposed to a
semiconservative one (model a). At the present time, our data did not enable us to determine
whether the negative-stranded MHV RNA was
everpresent as freemolecules. Our data also did
notenable us to rule out thepossibility that there
were very small amounts ofnegative-stranded
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.491.259.447.73.310.2]subgenomic RNA. However, if this was the
case, the rates of synthesis of the smaller
mRNAs must have been very fast, since the
amount of smaller subgenomic RNAs far
ex-ceeded thatofgenomicRNA.
This replication scheme raises an interesting
question: how is the synthesis of various
mRNAs regulated? It hasbeen shown that the
smaller mRNAs (no. 7 and no. 6) arefar more
abundant than the rest of the mRNAs (4, 5, 9).
Furthermore, the relative rate of synthesis of
most of mRNAs is constant throughout viral
replication (9).Therefore,there mustbe a
mech-anism which regulates the frequency of tran-scription of each mRNA species on the
negative-stranded RNA template.This remains one of the
outstanding unansweredquestions regarding the
replication scheme of MHV RNA.
It should be noted that ourfindingof a single
negative-stranded genomic RNA is
compati-blewithpreviously reported UV transcriptional
mapping data (4). In that study, UV light was
administered toMHV-infected cells at6 h p.i.,
at which time synthesis of negative-stranded
RNA should have already been completed.
Therefore, the dataobtained in that study
per-tain only to the synthesis of positive-stranded
RNA. Our present findings also suggest that if
RNA splicing or another modification was
in-volved in MHV RNA synthesis, it most likely
took placeduring thesynthesis ofmRNA, rather
than during the synthesis of negative-stranded
RNA. Therefore, late RNA polymerase, rather than early RNA polymerase (1), may possess
properties which areresponsible for such RNA
processing.
Our present findings thus distinguish the
mechanism of RNA synthesis for coronavirus
from that for other positive-stranded-RNA
vi-ruses, e.g., togavirus and picornavirus. In the
case oftogaviruses, several genomic and
sub-genomicRF RNAshavebeen detected(12, 14).
There is asubgenomic RF RNAcorresponding
tothesubgenomic mRNAofalphaviruses.
Fur-thermore, the RI involved in the synthesis of
alphavirus subgenomic mRNAcontainsa
nega-tive-stranded genomic template which is
sensi-tive to RNase atthe site of initiation of
subge-nomic mRNA(12). Thisappeared not to be the
case with coronaviruses, which use a unique
mechanism ofRNA synthesis. Further
under-standing of coronavirus RNAsynthesiswill
re-quirethe study ofthe sequences ofthe mRNAs
and the detailedstructureofthe RI. Such
experi-ments are in progress.
ACKNOWLEDGMENTS
We thank Todd Lasman forexcellenttechnical assistance and Raymond Mitchell for assistancein manuscript prepara-tion.
This work was supported in part by grant PCM4507 from the National Science Foundation,grantRG1449-A-1from the National Multiple Sclerosis Society, andPublic Health Ser-vice grants NS 15079, AI 19244, and CA 16113 from the NationalInstitutes of Health.
LITERATURECITED
1.Brayton, P. R., M. M. C. Lai, C. D. Patton, and S. A. Stohlman.1982.CharacterizationoftwoRNApolymerase activities induced by mouse hepatitis virus. J. Virol. 42:847-853.
2. Cheley,S., V. L. Morris, M. J.Cupples,and R.Anderson. 1981. RNA and peptidehomology amongmurine cdrona-viruses.Virology 115:310-321.
3. Content, J., and P. H. Duesberg. 1971. Base sequence differencesamong theribonucleic acidsof influenza virus. J.Mol.Biol.62:273-285.
4. Jacobs, L., W.J.M.Spasan,M.C. Horzinek, and B. A. M. van derZeIjst.1981.Synthesis of subgenomicmRNA'sof mousehepatitis virusisinitiated independently: evidence fromUVtranscriptionmapping. J. Virol.39:401-406. 5. Lai, M. M. C., P. R. Brayton, R. C. Armen, C. D. Patton,
C.Pugh,andS. A.Stobhnan.1981.Mousehepatitisvirus A59: mRNA structure and genetic localization of the sequencedivergence fromahepatotropic strainMHV-3. J. Virol.39:823-834.
6. Lai, M. M. C., C. D. Patton, and S. A.Stohlman. 1982. Further characterization ofmRNA's ofmousehepatitis virus:presenceof common5'-end nucleotides. J. Virol. 41:557-565.
7. Lai, M. M. C., and S. A.Stobhlman.1978. RNA of mouse hepatitisvirus. J. Virol. 26:236-242.
8. Lai, M. M. C., and S. A. Stoblman. 1981.Comparative analysis ofRNA genomesofmousehepatitis viruses.J. Virol. 38:661-670.
9. Leibowltz, J.L., K. C.Wilbelmsen,andC. W. Bond. 1981. The virus specific intracellular species oftwo murine coronaviruses: MHV-A59 and MHV-JHM. Virology 114:39-51.
10. McMaster, G. K., andCarmichael,G.G.1977.Analysis ofsingle-anddouble-stranded nucleic acidson polyacryl-amideand agarose gels by using glyoxal and acridine orange. Proc. Natl. Acad.Sci. U.S.A. 74:4835-4838. 11. Siddell,S., H. Wege, A.Barthel,andV. terMeulen.1981.
Coronavirus JHM:intracellularproteinsynthesis.J. Gen. Virol. 53:145-155.
12. Simmons,D.T., and J. H. Straus. 1972.Replication of Sindbisvirus. II. Multiple forms of double-stranded RNA isolated from infected cells. J. Mol. Biol.71:615-631. 13. Spaan,W.J.M.,P.J. M.Rotter,M.C.Horznek,and
B. A. M.van derZeUst.1981.Isolationandidentification of virus-specific mRNAs in cells infected with mouse
hepatitis (MHV-A59).Virology 108:424-434.
14. Strauss,3.H., andE.G. Straws.1977.Togaviruses,p. 111-166. InD. P.Nayak (ed.),The molecularbiologyof animalviruses, vol.1MarcelDekker, Inc.,New York. 15. Thomas,P.s. 1980.Hybridizationof denatured RNAand
small DNAfragmentstransferredtonitrocellulose. Proc. Natl.Acad. Sci.U.S.A. 77:5201-5205.
on November 10, 2019 by guest
http://jvi.asm.org/