Vol. 62, No.4 VIROLOGY, Apr. p.
0022-538X/88/041355-09$02.00/0
Copyright © 1988, American Society for
Microbiology
Specificity of Cleavage in
Replicative-Form
DNA
of
Bovine
Herpesvirus
1
WOLFGANG
HAMMERSCHMIDT,lt
HANNS LUDWIG,' AND HANS-JORG BUHK2*Institutfur Virologie derFreien Universitat' andRobertKoch-Institut des Bundesgesundheitsamtes,2 Nordufer20, 1000Berlin
65,
FederalRepublic of
GermanyReceived 13 October 1987/Accepted 30 December 1987
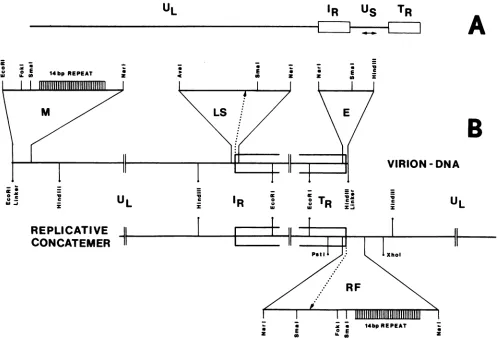
The linear double-stranded DNA genome of herpesvirus as it is present in infectious virions needs to be circularized after infection of host cells and before DNA replication. Replicative-form genomes have to be cleaved into linear unit-length molecules during virion maturation andare most probablythesubstrate for inversion of the shortsegmentrelativetothe longsegmentof thebovine herpesvirus1(BHV-1)genome.Those regions of the BHV-1 genomewhich are functionally involved in these processes havebeen analyzed atthe molecular level by cloning andsequencing the genomic termini, the fusion of both termini from replicative-form molecules, and thejunction betweenthe short and the longgenomesegment. On the basis of the simple
genomearrangementofBHV-1, itwasinferable thatthecleavage ofreplicative-formgenomesbyahypothetical
BHV-1terminase activitymaybespecifiedby asequenceattheleftendof UL
(An
element), which is located proximaltoareiterated , element that makesupthecleavage siteitself. Therelationshipof those elements inBHV-1 and the comparison to similar regions of other herpesviruses indicate consensus sequence elements
whicharefunctionally important forcleavage and isomerization of viral DNA during maturation of virions.
Bovine herpesvirus 1 (BHV-1) causes infections of
eco-nomic importance in cattle. Definedclinical symptoms lead tothedifferentiation of infectious bovine rhinotracheitisand
infectious pustular vulvovaginitis virus. Attempts to define
biological
markers oforgan tropism andvirulencecorrelat-ing withthe twoclinicalBHV-1 subtypes(infectious bovine rhinotracheitis and infectious pustular vulvovaginitis) gave
divergent results. By restriction enzyme analysis, one can
differentiate atleast twogroupsofBHV-1isolates. Whether these groupsdo ordo not correspond to the clinical subdi-vision into BHV-1 subtypes remains unclear (11, 12, 26).
Immunological investigation ofviral proteins and polypep-tide patterns do not elucidate the relatedness of clinical
subtypes ofBHV-1 andserological groups (25, 26). The life cycles of herpesviruses include virus-specified
recombinational events. All herpesviruses studied to date
replicate DNA during the productive phase of their life cycles by pathways which require the specific joining and
eventual severing of the ends ofthe viral DNA found in
virions. Many ofthe herpesviruses also recombineDNA at
internal sites such that their virions contain one DNA
molecule, which is one of either two or four possible isomeric forms. These recombinational events have been
studied mostwithherpes simplex virus type 1
(HSV-1).
Its DNA occurs in four isomeric forms and is therefore morecomplex than that ofBHV-1, which has
only
two isomers.We havecapitalized on the relative simplicity ofBHV-1 to
identify signals within its DNA that specify cleavage of replicating
species
toyield
virion DNA and thatspecify
recombinationatinternal sites.
BHV-1 is a member of the a
subfamily
of the bovineherpesvirus (19). The135-kilobase
(kb)-spanning
genome of BHV-1 is composed ofaunique long
segment UL(100
kb),
aninternal repeatIR(11
kb),
aunique
short segmentUs
(13
kb), and a terminal repeat TR
(Fig.
1A). The repeats are*Corresponding author.
tPresent address: McArdle Laboratory for Cancer Research, University ofWisconsin, Madison,WI 53706.
inversely oriented with respect to each other. The
Us
segment situated in between IR and TR appears in two alternative directions with respect to UL. Thus, DNA iso-lated from virions exhibits two isomeric forms in roughly
equimolaramounts. The twoisomers are designated proto-type (P) and inversiontype (IS) of Us (11, 13, 23). Nothing is known about possible functions of this flip-flop mecha-nism. But it is noteworthy that, in contrast to
Us,
the orientationofUL is fixed. The biologicalimplications ofthisinversionare notknown.
The BHV-1 genome structure represents a D-class herpes-virus(33) andisalsoexemplified inpseudorabies (PsR)virus
(3),varicella-zostervirus(VZV) (6), and equine herpesvirus 1, 3, and 4(5, 16, 39).
Here wereport the resultsofaninvestigationonregions of
the genome which arefunctionally important for viralDNA
replication. Initially after cell infection, the termini ofthe
linear genome are supposed to become ligated to form a circular molecule. Such circularization to replicative-form
genomesallows thecomplete replicationofotherwiselinear
molecules, e.g., by the rolling-circle model leading to the
accumulation of endless concatemers in the cell (18). By cleavageof these concatemers,
complete,
linear virion DNA units are provided forpackagingduring
virus maturation.The less-complicated situation with
regard
toisomeriza-tion in the BHV-1 genome
compared
with that in HSV-1provides a better chance to define
functionally important
sitesforthe
recognition
andformationofgenomic
termini,
as well as for the recombination event which leads to theinversion of
Us.
Thesimplicity
of the BHV-1 genomecompared with the HSV-1genome allowed usto define the
recognitionandcleavage sites ofan endonuclease
perform-ing cleavage of concatemers into virion DNA and
probably
inversionof
Us.
Thefunctional DNA sites of theenzyme(s)
called terminase-recombinase are conserved in
herpesvi-ruses with different genome
organizations.
Animproved
knowledgeabout the mechanismtogenerate
replicative
and matureformsofherpesvirus
genomes couldpossibly provide
tools fortherapy of
herpesvirus
infections. 1355on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS ANDMETHODS
Cells and virus strain. BHV-1 Bi (Schonboken) and the cell-culture-adapted derivativeB4were agenerousgiftfrom
0. C.Straub, Federal Research Centerfor Virus Diseasesof
Animals, Tubingen, Federal Republic of Germany. Virus was propagated in Georgia bovine kidney cells as reported
earlier(13).
Preparation of viral DNA and restriction endonuclease cleavage. DNA was obtained from lysed nucleocapsids,
which were purified from cell culture supernatant as
previ-ously described (13). Viral DNA from infected cells was
isolated 10 h after infection (multiplicity of infection, 0.1). Cellswerescraped off and lysed with sodium dodecylsulfate
andproteinase K. The DNAwasfurtherpurified by equilib-rium density centrifugation in CsCl-ethidium bromide gradi-ents. Restriction endonuclease cleavage and analysis were
doneaspreviously described (13).
Derivation and construction of recombinantplasmids. Clon-ing of terminal DNA fragments of BHV-1 was performed
aftermodifying the genomic termini. Whole-virion DNAwas
treated with a large fragment of DNA polymerase I in the
presence of the four deoxynucleoside triphosphates to
gen-erateblunt ends.32P-phosphorylated dodecamer EcoRI link-ers (for the left-hand terminus) or decamerHindlIl linkers (for the right-hand terminus) (New England BioLabs, Inc.)
wereligated with T4 DNA ligasetothe virion DNA. Linker molecules and virion DNAwerecleavedwithacombination
oftworestrictionenzymes(HindIll andEcoRI). Cut linkers and cleaved virion DNAfragments were separatedon
ana-lytical agarose gels. The gels were dried and exposed to X-ray films. Autoradiographs allowed the identification of theterminal fragments in the EcoRI-HindIII double digests of virion DNA. Genomic terminal fragments were isolated from preparative gels and ligated to an EcoRI-HindIII-cut
pUC8vector.Thisprocedureensuredcloningof the terminal fragments in defined orientations. Competent JM83 cells (24)
weretransformed (14), and recombinant clones were
identi-fied by colony hybridization, restriction enzyme mapping, and Southern blot hybridization. The series of plasmid clones bearing the left terminus was calledM (13), and the series bearing the right terminus wascalled E.
Clones of the BHV-1 fragment carrying the internal junc-tion betweenUL andIRwereobtained after isolation of the
EcoRI-HindIII fragment spanning the region from the right endofULtoIR (Fig. 1). The plasmid clonesweredesignated
LS. Cloning of the BHV-1 fragment carrying the fusion of left andright genomic termini appearing in replicative-form molecules was achieved after cleavage of total DNA of
virus-infected cells with PstI and XhoI, separation of the resulting fragments on apreparative agarose gel,elution of
thefusionfragment, and ligationtotheappropriately cleaved (PstI and Sall) pUC8 vector. Recombinant plasmid clones designated RF harboring the fusion of the left and right genomic termini were identified by colony hybridization.
DNAsequencing. DNAwas sequenced by the method of Maxam and Gilbert (21, 22) after end labeling the cloned fragments with polynucleotide kinase (Stehelin AG) and [.y-32P]ATP (Amersham Corp.). Left-terminal DNA frag-ments were sequenced by the chain terminator method according to Sanger et al. (34) as described earlier (13).
Reaction products were separated in 20, 8, and 6% poly-acrylamide-urea gels whichwereboundtoglass plates, dried afterelectrophoresis, and autoradiographed withXAR-5film (Eastman Kodak Co.).
Blothybridization. Restriction fragmentsweretransferred
(35) after electrophoresis on agarose gels to GeneScreen membranes (New England Nuclear Corp.). In vitro labeling of recombinant plasmid DNA by nick translation (31) in the presence of [a-32P]dCTPor [aL-35S]dCTP and hybridization
tothe immobilized DNAfragments were done asdescribed previously (13).
RESULTS
Cloning and characterization of thegenomic termini. We modified theterminal fragments of BHV-1 DNA such that the left terminuswastaggedwithsyntheticDNAcontaining
anEcoRIrecognitionsite and therightterminuswastagged
with asyntheticDNAcontainingaHindIIIrecognitionsite. Cleavage of the modified virion DNAs both internally and within the syntheticDNAs yieldedterminal fragments with
characteristic orientations (Fig. 1). Itwas possible toligate
suchfragmentsindefined orientations into theappropriately cleaved vector, pUC8, with a reasonable efficiency. As
described earlier (13), recombinant M-seriesplasmidclones
bearingthe left-endHindIIIfragmentof the genome included a setofclonescontaining fragments rangingin size from 2.4 to 2.8 kb. In contrast, the recombinant E-series plasmid
clones containing the right-end EcoRI fragment ofTR had
inserts of only one size, 8.2 kb. Physical maps of the
genomic terminiwere deduced andare presentedin part in
Fig. 1.
Cloningandcharacterization of the internaljunction
frag-ment of UL-IR. TR and IR are two large (11-kb) inverted repeats bracketing
Us
(Fig. 1). To locate precisely thejunctionbetweenULand'RStheEcoRI-HindIII LSfragment
(9.9 kb) carrying the junction was cloned in pUC8. We
generatedaphysical mapofthe LSfragmentandcompared
it to the physical map ofthe right genomic terminus. This comparisonidentifiedanAvaI-NarIfragment (210basepairs [bp])thatcontained boththeleft end of'R which is identical to the rightend ofTR, and the right end of UL. Thus, this AvaI-NarI fragment harbors the internaljunction between ULand
IR-Identification, cloning, and characterization of the fusion fragment. The free termini as present in virion DNA of herpesviruses disappear after infection of cells. This appar-entloss has been described for D-type herpesviruses, such
asPsRvirus(2, 3) andVZV(7),aswellasforherpesviruses
with different genomic arrangements, such as HSV-1 (30). Left andright genomic termini became fusedin head-to-tail
arrangements, whichare considered to bereplicative
inter-mediates ofthe viral genome. BHV-1virion DNAand total DNAofBHV-1-infected cellswere cleavedwith restriction enzymes(e.g., HindIII). The cleavage products were sepa-rated onanalytical agarosegels, transferredto nylon
mem-branes, and probed by hybridization with DNA sequences
specific for the left genomic end of BHV-1. Virion DNA exhibitedonly onesignal ontheautoradiograph,
represent-ing the left-terminal DNA fragment (Fig. 2). In contrast, cellular DNApreparationsshowed twosignals afterHindlIl digestion,onesignalcorresponding to the left-terminal frag-mentand the second, weaker signal representing two DNA
fragments of approximately 15 to 17 kb (the two fragments are not separated in Fig. 2). The size of these newly identified fragments is explainable by fusion of the left-terminal fragment (2.4 to 2.8 kb) to the right-terminal frag-ment(12.5 or 14.5 kb), as would be expected in replicative-form molecules. Similar analysis of DNA cleaved by other
restriction enzymes gave cleavage patterns which were consistent with the presence of a fusion fragment in
on November 10, 2019 by guest
http://jvi.asm.org/
REPLICATIVE-FORM DNA OF BHV-1 1357
UL
IR
US
TR
0
A
I
B
VIRION
-DNA
6.
O,.
wlJ -i
U
IR
0 oR
zUL
REPLICATIVE
CONCATEMER
-. a *X l4bp REPEAT
a 0E 0EE
I Iz
FIG. 1. Genome arrangement ofBHV-1 and cloning strategy for the left-hand (M-series plasmids) and right-hand(E-series plasmids) genomic termini, theinternaljunction UL-IR(LS-series plasmids), and the fusion of the right and left genomic termini (RF-series plasmids). (A) The genome arrangement of BHV-1consists of the unique long segment UL, aninternal repeatIRS the unique short segment
Us,
and a terminal repeatTR.IR and TR are inverted relative to each other, bracketingUs. Usitself occurs in two differentorientations relative to UL, IRS andTR, giving rise to two equimolar genomic isomers (indicated by a double-headed arrow). The orientation ofUL is fixed relative toIR andTR.(B)Relevantrestriction enzyme sites used for generating clones of the two termini and theUL-IRjunction as they occur in the virion DNAmolecule (M, LS, and E series, top line), as well as of the fusion fragment of the left and right genomic termini in the replicative-form molecule(AFseries, bottom line),aresketched. The localizationof the 14-bp repeat array at the left genomic terminus is shown. The detailed restriction enzyme maps of the different regions are based onthenucleotide sequences (Fig. 4 to 6).cellular viral DNA. Furthermore, these Southern blots did
not reveal any additional signals even when overexposed
(datanotshown).Theobservationsdemonstrate that UL, in
contrast to
Us,
doesnotinvert relativeto IRin virion DNA. Inversion OfUJL
with respect to IR would create a newHindIII left-hand DNAfragment differing in size from the
cloned left-terminal
fragment.
In addition, were UL toin-vert, the inversion would create a newdiscerniblejunction
with IR. In contrast to other D-type herpesviruses (6), this putative second junction with 'R was never observed for
BHV-1.
Nucleotide sequence determination and sequence analysis.
Both strands ofthe inserts in the recombinantDNA clones
harboring the left (M clones) and right (E clones) genomic termini, the junction fragment UL-IR (LS clone), and the
fusion fragment (RF clones) (Fig. 1) were sequenced. The
nucleotide sequence derived fromsix
plasmid
clonesofthe Mseriesis shown inFig.3. Asreportedearlier,theterminal NarIfragment (Fig. 1)includesa14-bptandem repeat array,giving rise to size
heterogeneity
of thisfragment
due tovariations in the copy number(8to 38) ofthe 14-bprepeats (13). The 235-bp-long DNA sequence distal to the repeat
arraywasidenticalinfive ofsixclonessequenced. The sixth
clone, M297,showed anadditional base pairattheposition
at which the linker had been ligated to the left genomic terminus and a few differences within the following 25-bp stretch compared with the five other M-series plasniid clones. The reason for these sequence differences is not known.
Relevant DNA sequences ofthe right terminus are
dis-played in Fig. 4. A total of six individually cloned right terminiwere sequenced. Comparison ofthe nucleotide se-quence from the right and left termini showed that the
termini do not exhibit sequence
redundancy. Aligning
the sequencesofthe cloned termini and the sequence spanning thefusion ofUL andTR (Fig. 5; a total of six clones weresequenced) revealed that a single base pair at the fusion
pointwas missingfor all cloned andsequenced fragmentsof the genomic termini, except for the already mentioned
plasmid clone M297. This finding indicates that because of thecloning procedure (seeMaterials and
Methods),
asingle-baseextensionatthe 3' endofthe
genomic
terminiwaslost,
presumablyas aresultofthe3'-to-5'exonuclease
activity
of DNA polymerase usedpreviously
to blunt the ends of the VOL.62, 1988on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.59.557.73.411.2]GGCGCCGGACCCAGGGCGGAGCCCAGAGCGGGCCCGGGCCCGGGCCCGCCGCGCCGAAAT CCGCGGCCTGGGTCCCGCCTCGGGTCTCGCCCGGGCCCGGGCCCGGGCGGCGCGGCTTTA
Marla
70 90 a
TTCCGCCCCCCCCCAAAAACACCCCCCCGGGGTTGCAGGGCCCCCG GCGGCGCGGCGCG AAGGCGGGGGGGGGTTTTTGTGGGGGGGCCCCAACGTCCCGGGGGCGCGCCGCGCCGCGC
L VSmall
GAGGG-3' CTCC--5'
FIG. 4. Nucleotidesequenceofthenaturalright genomic termini ofBHV-1representedby plasmidclone E16. Thesequenceisgiven starting from the most distalNarl siteextending tothe right end. The aelementand its relative orientation isindicated, the localiza-tionoftheyelementisshown, and restrictionenzymesitesgiven in Fig. 1 areindicated.
FIG. 2. Detectionof the fusion of left andright genomictermini of BHV-1 by Southern blotting. Virion DNA (extracted from purified capsids [lane 1]) and whole-cellDNAof GBK cellsinfected with different plaque-purified BHV-1 stocks (lanes 2 and 3) were
digested with HindIII, separated on agarose gels, blotted (35) to
nylonmembranes, andhybridized toplasmidcloneM23. The
auto-radiograph shows signals which refer to the left-terminal HindIlI fragment (M). Theirsize heterogeneity is due to a differentcopy number of the 14-bprepeatsinplaque-purifiedvirusstocks(13). In restricted cellular DNA, additionalfragmnentscanbedetected which correspond to the HindIII fusion fragment of the left and right genomic termini (RF).
M297
-~ An 50
5'-GGTGCAGGGCCCCCGCGCGGCGCGGCGCGGAGAAAAAAAAAATTTTCTCTGCGCGGCGCGT 3'-CCACGTCCCGGGGGCGCGCCGCGCCGCGCCTCTTTTTTTTTTAAAAGAGACGCGCCGCGCA
M23
______________An_50
5'--GGCCCAGCCCCCGCGCGGGGGGCGCGGAGAAAAAAAAAATTTTCTCTGCGCGGCGCGT
3'-CCCGGGTCGGGGGCGCGCCCCCCCGCGCCTCTTTTTTTTTTAAAAGAGACGCGCCGCGCA
Fo0k
130 150 170
ACACTGCCAAGATCACCGAGCCTGTGCGCGGCCATCTTGCTTCCAAACTCATTAGCATAC
TGTGACGGTTCTAGTGGCTCGGACACGCGCCGGTAGAACGAAGGTTTGAGTAATCGTATG
SauSA
190 210 230
CCCGCCCACTATTCCATTCTCATTTGCATACCCACCATCGCACATGCCGCCATATT ...
GGGCGGGTGATAAGGTAAGAGTAAACGTATGGGTGGTAGCGTGTACGGCGGTATAA...
630 650
GCTCCTCCTCAAAACACTACCGCGGGCGTCCG CGAGGAGGAGTTTTGTGATGGCGCCCGCAGGC
virion DNA molecules. If thesingle-base extension hadbeen at the 5' end, then our cloning procedures would have
maintained this nucleotide (see Materials and Methods). A similar artifact has been reportedin the cloning of HSV-1 (29), VZV (6), and human cytomegalovirus (HCMV) (40) termini. Since the missing base pair is retained in fusion fragments, we conclude that for the right terminus the 3'
single-base extension is a guanosine, whereas for the left
terminus the 3' base extension is a cytosine, which was
taken into account in Fig. 3 and 4. As a result of the
sequence of the fusionfragment, the genomic termini fused
via the complementary 3' base extensions.
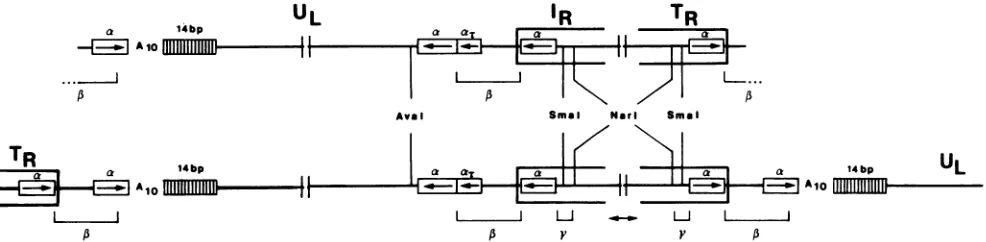
By comparing the nucleotide sequenceatthefusion ofthe termini with the sequence atthejunction UL-IR, it became evident that the fusion andjunction sequences were
identi-cal, considering their inverse orientations (Fig. 5 and 6). Additionally,a 18-bpsequenceelement, consisting of C and Gonly, is reiterated fourfoldinthegenome,always located
close to the fusion and junction. This 18-bp reiterations, referred to hereafter as a elements, display remarkable
features. (i) The a sequence 5'-CCGCGCGGCGCG
GCGCGG-3' differs from a palindromic structure only at positions 7 (G) and 12 (G). Thesetwodifferences define an
orientation for the a elements. (ii) Single a elements are
located close to the left terminus andtothe right terminus (Fig. 3 through 6). Both of thesea elements have the same
orietitation. (iii) The a element located atthe left terminus
has a variant sequence, as determined from sequencing
differentplasmid clones. Transversionsappearatposition 9 (CtoG) andatposition11(CtoG), resulting inanincreased
divergence from a palindromic structure. However, these transversionsarelocated in themiddle of the palindrome and
RF2
30
Marl
90
50
a.. TR UL
CCCCCCCCAAAAACACCCCCCCGGGGTTGCA0GGGGCCCGCGCG0CGCGGCGCGGA0G_C
GGGGGGGGTTTTTGTGGGGGGGCCCCAACGTCCCCCGGGCGCGCCGCGCCGCGCCTCC
670
CTCTCACTAGCTTCGGCGCC GAGAGTGATCGAAGCCGCG
[image:4.612.124.239.75.286.2]Mar
FIG. 3. Nucleotide sequence of plasmidclones M23 and M297
covering the left-hand genomic terminus. Sequences are given
starting from the left genomicterminustothemostdistalNarIsite. Severalrestriction sitesare indicated for comparison with Fig. 1. The3'single-base extensionatthe natural left end ofthegenomeis
deducedfromthe fusion of bothgenomic termini. Thesequencesof
the functionally important a elements are boxed, positions of
variable nucleotidesaremarked by dots underneath the sequence,
thearrowgives the relative orientation of the a elements, the An
stretchismarked, and the 14-bprepeatarrayis shown inbrackets.
.F. *ma
150 170
I GCCCAGCCCCCGCGCGGGGGGCGCCGAGAAAAAAAAAATTTTCTCCGCGCGGCGCGTGCA CGGGTCGGGGGCGCGCCCCCCGCGCCTCTTTTTTTTTTAAAAGAGGCGCGCCGCGCACGT
190 210
TTGCGGCGGGCGGGGGCGGGGTGGGGGATG
AACGCCGCCCGCCCCCGCCCCACCCCCTAC
Fok
FIG. 5. Nucleotide sequence of the fusion of left and right genomic terminioccurring inreplicative-formDNArepresentedby
plasmid clone RF2. The fusion between TR and the left end ofthe UL segmentisindicated, showing the cleavage byaterminase activity.
Thecleavage site ispartof the P element, encompassed bytwo a
elementswhicharearranged in the same relative orientation. The
consensussequence-yof theright-hand termini of different
herpes-virusgenomesisunderlined.Restriction sites refertoFig. 1.
1 2 3
-4---- R F
CTCCTCCTCCCI
GAGGAGGAGGG
20 v
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.327.555.76.141.2] [image:4.612.72.297.443.637.2] [image:4.612.327.557.563.654.2]LS37
10 30 50
5'-CTCGGGGTGACGCGCCGGGAGACGGCCGCGAGGGCGGGGGCGGGGCGGGGGCAGGGCGC
3'-GAGCCCCACTGCGCGGCCCTCTGCCGGCGCTCCCGCCCCCCGCCCCGCCCCCGTCCCGCG
BHV-1
70 90
CGGGCGGGCGGCGAAAAGGCCCGCACCCCCCGGCCGCCCCAAGGCGAAACCCCCCACGCA GCCCGCCCGCCGCTTTTCCGGGCGTGGGGGGCCGGCGGCGTTCCGCTTTGGGGGGTGCGT
130 a. __ UL IR
AGGCGCGACGCCCGCGCCGCGCCGCGCGGCCCCCCCCGCGGTGCC-CC TCCGCGCTGCGGGCGCGGCGCGGCGCGCCCGGGGGGGCGCGCCCCCGACCCGCGAGGC
a 210
CGCCGCGCCCOCCGCGGGCCCTGCAACCCCGGG
GCCCGCGGCGCGCCCCCGGGACGTTCGGGCCC
FIG. 6. Nucleotide sequence ofthejunction between the right endofUL and IR. Thederivation of thisDNA sequence is shown in Fig. 1 with corresponding restriction enzyme sites. The junction point is deduced by similarity with thefusion sequence shown in Fig. 5. The junction is located withina1Belement, which is inverted relativetothefusion fragment(1B').The1 elementissurrounded by two complete (ax') and one partially duplicated
(cx',)
ax elements. Note that incontrast tothe fusion fragment, anAn stretch is not presentin thissequence (Fig.5).could form a loop in a possible hairpin structure. (iv) Since the a element at the right terminus is part of TR and is oriented rightwards by definition (Fig. 4), the analogous a element in 'R should be inversely oriented. This interpreta-tion was confirmed by sequencing thejunction (Fig. 6). (v) At the junction between UL and
'RI
the right end of ULcontainsacompleteaelement, whichisdirectly preceded by
atruncated(14-bp)aelement(at). UL is therefore bracketed
by twoinversely oriented a elements. This arrangement is
also found in the region
IR-Us-TR.
These findings arede-picted in Fig. 7.
Alignment ofthenucleotide sequenceoftheTR-ULfusion
fragment and the related IR-UL junction fragment in an
inverse orientation (Fig. 5, 6, and 8) leads to two
conclu-sions. (i)Thenucleotide sequences at the fusion point of TR andUL and at thejunction of UL and'R areidentical. The sequence identity between the parts of TR and IR was
expected. However,thehomologyextends across thefusion
andjunction and includes astretchof28bp atboth endsof
UL. We havedesignatedthis 28-bpregion, whichcovers 14
bp of the a-element sequence, a 1 element (Fig. 7). (ii) Cleavage of replicative-form DNA generatingfree genomic termini of virionDNA occurs within the ,B element and leads to3'
single-base
extensions(Fig. 5). Thus,the sketch(Fig. 7) ofthe BHV-1 genome can be completed and described byTR AIA
L&A
LeftEndUL
FUSION
IR
1}--"Jl
- - JUNCTIONPA
vzv
LeftEndUL
FUSION
RlghtEndUL JUNCTION
A10
FIG. 8. Schematic comparison of the fusion and junction of BHV-1 contrast to VZV. The segments (UL,TR, andIR) make up similar nucleotide sequence elements (a elements and
A.
stretch)in bothvirusgenomes. Thecleavage ofthose genomesin their repli-cative form is indicated by vertical arrows. In contrast to VZV, cleavageoccursinthe BHV-1 genomeonlyin thefusionfragment of TR and UL. In the VZV genome, putative cleavage sites are encompassed by identical sequenceelements(IR, TR, ana-similar element[boxed], and theAnstretch[A1o]). In contrast, theputative cleavage site in the fusion of replicative-form DNA in BHV-1 is composed ofaunique arrangement of sequence elementscompared with the internal junctionIR-UL-thecombinationof reiterated and nonreiterated elements
(,B'
stands forinversely oriented 1 element):(24.5 bp)
P-UL-P'
(28bp) IRUS-TRP
(3.5 bp) The 1 elements makeup thecleavage site for the enzyme(s) performing the cleavage event in the concatemeric mole-cules to generate mature, virion DNA. We propose to call thisenzymatic activityBHV-1 terminase. The concatemeric arrangement of BHV-1 genome units in a head-to-tail man-nerresults in theappearance of two13elementsin onevirion genome unit, which can be depicted as:,B
UL
13'IR-US-TRP-UL-1'IR-Us-TRI3-UL
ak 1ibp
-1
A 10uJ_
...
A
TR 14bp
/3r
a
UL
-4 -I
4'-IR
TR
I
IX HI
-Ava I SmaI Narl Smai
IL L _l-11 I
A Y Y A
FIG. 7. SkeletonsketchofBHV-1virionDNAmolecule (top line)andofapartof the
replicative-form
DNAoccurringinconcatemers(bottomline).The sketch isnot toscale,and thepairsofvertical linesinterruptingeach molecule indicate wherelargeareasof the DNAare
notdepictedfor convenience ofpresentation. See Fig. 1 fora less-detailed sketch. The virion DNA molecule is builtof UL,
IR'
andTRharboring morphologically importantarrays (14-bprepeats and An stretch [Alo])and elements (a, l,and y), as shown in Results. In the replicative-formDNA, thevirionDNAunitsarelinkedtoeachother,formingacomplete 13 elementatthe fusions. Asequence-identical1 elementoccursin thejunctionUL-IR.The virion DNAmolecule showstwoparts ofasinglecleaved 1 elementatthe termini. The localization of restriction enzyme sitesarealsogiveninFig. 1,4,and 6.
4bp
UL
I
AloVOL. 1988
TR
i - Aioon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.319.560.67.267.2] [image:5.612.64.297.77.173.2] [image:5.612.68.561.540.661.2]I,
AAAAACA mTTrGT
TTTAGACCA AAATCTGGT AAAATAA TTTTATT TTTAAG AAATTC AAACACA TTTGTGT AAAACACA TTTTGTGT AGGAAAGA TCCTTTCT ACTGATTTT TGACTAAAA TAAAAACA ATTTTTGT ATCAAAATA TAGTTTTAT
CCCCCCCGGGG GGGGGGGCCCC
CGCCCCCC GCGGGGGG
CCCCCCCCCGG GGGGGGGGGCC
CCCCGCCCCC GGGGCGGGGG
CCCCCCGGGGG
GGGGGGCCCCC
Ub
CCCCCCGGGGG GGGGGGCCCCC
Ub
CCCCCGGGGGGC GGGGGCCCCCCG TR
CCCCCCGGGGG GGGGGGCCCCC
CCCCCCCGCCCC GGGGGGGCGGGG
CCCCCCCGGGGGGCCCGCGCGC GGGGGGGCCCCCCGGGCGCGCG
TTGCAGGGCCCCCGCGCGGCGCGGCGCGGAGGG AACGTCCCGGGGGCGCGCCGCGCCGCGCCTCC
TAAGCCCACGCCCCCACTCTGCGCCCGACCCC TTTCTGGGCGCCCGGCGGACCCCGGGAGAGG
TTTTTTCGCGGCCCCCGGGGAGCGGGGGGTGGC
AGCGCGCGCCGGGAAATTTCGCGCCGCCGC -5'
DR
TCGCGCGCGGCCCTTTAAAGGCGGGCGGCGGGC-3' AGCGCGCGCCGGGAAATTTCCGCCCGCCGCCC -5'
DR
TTCGGGGGGTGTTGGAGAGGGC-3' ?
AAGCCCCCCACAACCTCTCCCG-5'
ATAACTTGGTAGGCAAAGAGGGGGAAGGG-3'
TATTGAACCATCCGTTTCTCCCCCTTCCC-5' TCGAGGACCCACCACGCGGCCCGGAATGGA-3' AGCTCCTGGGTGGTGCGCCGGGCCTTACC -5'
ACTCAGACGGCCGGGGGG-3' TGAGTCTGCCGGCCCCCC-5 ?
GGGX-3'
ccc -5'
5,-GGCCAGCCCGCGGGGGGWCGAGCa 3' -CCCGGGTCGGGGGCGCGCCCCCCCGCGCCTC
5'- GGCTAGGCTCTCTCTCGGGCGCGGGCCCGTG 3'-CCCGATCCGAGAGAGAGCCCGCGCCCGGGCAC 5|-AGGCCAGCCCTCTCGCGGCCCCC!TCG-AGAGAG
-CTCCGGTCGGGQGAGCGCCGGGGGAGCTCTCTC
3
5'-GGCCCAGCTCTCCCCCGAGCGCGGATCTCTG
3'-CCGGGTCGAGAGGGGGCTCGCGCCTAGAGAC
0-.-* _.
5'-AGCCCGGGCCCCCCGCGGGCGGGGCGGCGCGC
3 -GTCGGGCCCGGGGGGCGCCCGCCCCGCCGCGCG
DR Uc
5'-AGCCCGGGCCCCCCGCGGGCGCGCGCGCGCGC
3'-GTCGGGCCCGGGGGGCGCCCGCGCGCGCGCGCG
DR UC
5'-ATGGGGGGAGCATGGGCCGCCGCGCATTCCTGG 3 -TACCCCCCTCGTACCCGGCGGCGCGTAAGGACC
TR
An
AAMAMAAAM
AAAAAAM
TrTTTI1r=
AAA&AAAAA
AAAAAAAMA TTTTTTTTT
A.AAAA
Mkkm
AAAAMAAA
Trmm AAAAA
TTTTTT AAAAAA TT~TTT
TTTTCTCTG...
AAAAGAGAC.. .
GGC...
AAAAAGCCG...
GCGACCCCA...
CGCTGGGGT....
TTTCCCGCC...
AAAGGGCGG.. .
GGCGGGCGG...
CCGCCCGCC...
GGCGGGCGG.. .
CCGCCCGCC...
GTGGAGGGG...
CACCTCCCC.. .
BHV-1
EHV-1
vzv
PsR
HSV-1 (F)
HSV-1(KOS)
EBV
5 -GGGCCCCAAGCAGTGGGCGCTGGGCAGCTGG AAAAA TGTCCTGGG...
3'-CCCGGGGTTCGTCACCCGCGACCCGTCGACC TT ACAGGACCC... HVS
5 -TGTCGGGCGTCCACCTAGATGGGTGCGCGCCCG GGAGGCGGC..
3'-TACAGCCCGCAGGTGGATCTACCCACGCGCGGGC CCrCCGCCT HCMV
5,- XGCXXXGXXCXCXCGCXGXXGXG
[image:6.612.97.510.66.690.2]3 -XXCGXXXCXXGXGXGCGXCXXCXC
FIG. 9. Nucleotidesequencecomparison. The followingarecompared: the right(a)andleft (b) naturalgenomic termini of BHV-1 (this
study), equine herpesvirus 1(EHV-1) (Chowdhury and Hammerschmidt, in preparation), PsR (15), VZV (6), HSV-1 F (29), HSV-1 KOS (43), Epstein-barr virus (EBV) (20), herpesvirus saimiri (HVS) (1), HCMV(41), and mousecytomegalovirus (MCMV) (41). Differentsequence
elements thathave beendescribedaredepicted (a, P,-y, and An),andthenatural ends oftheherpesvirusgenomes areindicated by giving
...ccccccccC ...GGGGGGGG
....ccccccccC
... GGGGGGGGG
.CCCCCCCCCG ...GGGGGGC
.cccccccccc
.GGGGGGGGGG
...CCCCCCCCG
...GGGGGGGGC
...cccccccc
...GGGGGGGG
...cccccc
...GGGGGG
....ccccc
...GGGGG
...CCCCCCCGC ...GGGGGGGCG
...ccccccc
...GGGGGGG
A...
a
B-3'
-s1 BHV-1
-sI EHV-1
3-3'
-5, vzv
G-3'
-5@ PsR
HSV-1 (F)
HSV-1 (KOS)
EBV
HVS
HCMV
MCMV
A3
b
on November 10, 2019 by guest
http://jvi.asm.org/
REPLICATIVE-FORM DNA OF BHV-1 1361 DuringBHV-1 DNAmaturation, theterminasecleavesonly
the ,B elementat thefusion TR-UL but not the I element at thejunctionIR-UL. Cleavage at that second 3 element would lead to virion DNA molecules carrying UL inversely ori-ented withrespect to 'R. Suchinversions were not detected in BHV-1 virions. However, both
P
elements do show anidentical potential cleavage site for the terminase. This
finding indicatesthat thespecificity ofthe BHV-1 terminase
reaction is mediated not only by the ,3 element but also by sequencesoutsideofthe
P
elementanduniqueto thefusion TR-UL. Since TR and IR consist of sequences which arereiterated,thespecific recognition site for BHV-1 terminase islocatedwithinunique sequences at the left end of UL.
Apoly(A)stretch (An)is located proximalto theleft-enda element (Fig. 3 and 8). In contrast, such an
An
stretch ismissing at the right end of UL, and instead there is a truncated (at) plus a complete a element. Consequently, it seemslikely that
An
is atleastpart of therecognition site for the BHV-1terminase which providesthespecificity required to recognizethe fusion ofTR-UL-DISCUSSION
Togainfurther informationabout theinvolvement of the
genomic termini of herpesviruses in DNA replication and
virus maturationand toidentify structuresthat are function-ally essential for these processes, we chose to study the
relatively simple arranged herpesvirus genome of BHV-1.
Apparently two major characteristics contribute to the ge-nome structure. (i) First, precise cleavage of concatemers leads to virion DNA units generating unique sequences at
both termini. The cleavagehasto beperformedat adistinct site by a endonuclease activity, a terminase. One has to
postulate thatrecognition ofthecleavage site is based on a
nucleotide sequence. The two
p
elements in a concatemeric BHV-1 genome unit couldtheoretically serve as aputativecleavage site to generate virionDNA units. The fact that a
ULsegmentofthe BHV-1genomeisfixed inits orientation withrespect to IR consequently means that only thepotential
cleavage
site in thep
elementattheleftendofULis cleavedbytheputativeterminase, whereas the
p
elementattherightendofUL isnot. Thus, therecognition site can notcoincide
withthecleavage siteoftheterminase. (ii)Second, inversion
of
Us
generates two isomeric molecules of BHV-1. Virusisolated from a single plaque in cell culture gave rise to progeny consisting ofan equal proportion ofboth isomers (datanotshown). Provided that onevirion has thecapacity
toformaplaque,then one has to conclude that isomerization is anearlyeventin asingle cycle ofvirusmultiplicationora
rather
frequent
one. Genome isomerizationrequires
are-combinationevent
involving cleavage
andrejoining
ofDNAmolecules. The similaritiesbetween the UL-IRjunction and
the TR-UL fusion in BHV-1 are consistent with the notion that both inversion of
Us
and cleavage ofconcatemers are events that areprobably
executedby
a similar oridenticalenzyme(s) withdifferent cofactors.
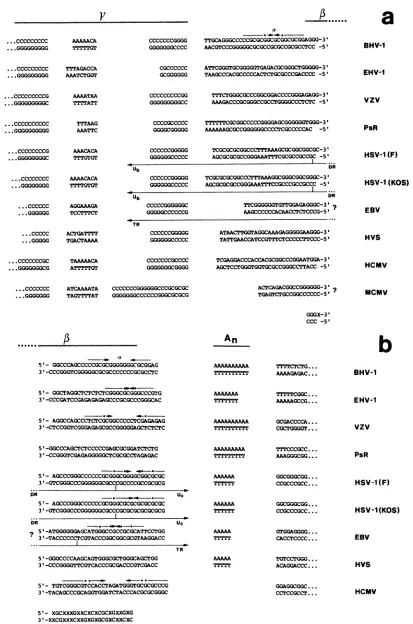
To define the recognition site of the terminase, we com-paredthe terminal nucleotide sequences of other herpesvirus genomeswith BHV-1. As we have already emphasized (13), BHV-1 exhibits the sequence 5'-GAGAAAAAAAAAA-3' from position 29 to 41 (Fig. 3), a sequence which is also presentin the VZV genome within the terminal repeat of its
ULsegment29nucleotides away from the genomic terminus (6). This is the only extended sequence homology shared at the UL termini between BHV-1 and VZV. We propose that thisconserved
An
stretchis part of the recognition site of the BHV-1terminase (see Results). To gain further information about the conservation of thisAn
sequence element, we compared available terminal sequences of different herpes-virusgenomes, including the termini of equine herpesvirus 1, a class D-herpesvirus genome (S. I. Chowdhury and W. Hammerschmidt, manuscript in preparation) (Fig. 9).Itis remarkable that except for HCMV, all herpesviruses bear a more or less extended
An
element 31 to 42 bp proximal to their termini, if sequences are aligned accord-ingly. Inaddition, most herpesvirus termini possess a palin-dromic structure located distal to theAn
stretch, similar to the a element of BHV-1. There is no obvious sequence identity in the palindromic sequences of comparable a elements in otherherpesvirusesdespitethe highGC content. Inno case has aperfectpalindromebeenfound;the stem of thepossible hairpin structures consists of 6 bp disruptedby one ormoremismatches. An aelement can be detected even in HCMV lacking theAn
stretch, whereas herpesvirus saimiri and PsR virus whichpossess anAn
sequence element exhibit a lessconvincing aelement. Aligningthenucleotidesequence of theright terminusof BHV-1 and other available terminal sequences (Fig. 9a) confirmed that the conserved sequence first reported by Tamashiro et al. (40, 41) also occurs in BHV-1. In accordance with our nomenclature
designating a and
p
elements, we propose to call this conserved areafromposition67 to 93 (Fig. 4and9a) in the BHV-1genome the -y element. Itis obvious that in contrast to the a element present at the right-hand terminus inBHV-1, there is noequivalenta elementat the right termi-nus ofother known herpesviruses. Similar sequence
com-parisons werepublishedrecently (9, 10, 41).
InHSV-1,the
An
stretchispartoftheUc
sequence (Fig. 9b)and the yelementis part of the Ub sequence(Fig.9a).Uc
andUbareunique nucleotide stretches in the "a" sequence
ofHSV separated bya repeat arrayconsisting of tandemly reiterated strain-dependent sequences (8, 27). This array of
tandemly reiterated sequences is structurally similar to the
14-bp repeats of BHV-1 (13). Distal to the Ub and
Uc
stretches,twodirect repeat sequences(DR)(Fig. 9aandb)
arearranged inthe sameorientation. Thelocation of reiter-ated "a" sequencesofHSV-1 can be
depicted
asanb-UL-b'a
mc'-Us-ca
(n and m may varyfrom 1 to10).TheULand
Us
components invert relative to each other,generating
fourequimolar
isomeric forms of the HSV-1 genome
(for
areview,
seethe strandorientations (5'or3'). Aquestion markpointstounknownendstructures. ForEpstein-Barrvirus,the terminal sequencewithin TR isinferredfromsequence homology. Itisnoteworthythatthis sequencealignmentand thedepictedsequenceelementsdonotallowto postulateany distinctlength-measuringmechanismperforminganaccuratecleavageofthePelements. To addressthis
question,
thebottom line showsa,-elementconsensusdeduced from thissequencealignmentmainlymakingup sequences of theleftgenomic
endsofdifferent herpesvirusgenomes.Thisconsensussequencecould contributetothe baseprecisecleavageeventwithin thepelementof theseherpesvirus
genomes. Thefollowingscoresshowanextended structuralsimilaritytotheproposed p-elementconsensussequences(number of nucleotides
outof15): BHV-1, 14;EHV-1, 13;VZV,11;PsR, 12;HSV-1(F), 14;HSV-1(KOS), 15;
Epstein-Barr
virus, 11;herpesvirus
saimiri, 10; and HCMV, 10. Arrowsindicatepalindromicsequenceelements;dotspointtomismatches within thepalindrome. Ub,
Uc,
DR,and TRreferto definedsequenceelementsinHSV-1 (29, 43)andEpstein-Barrvirus(20).VOL.
62,
1988on November 10, 2019 by guest
http://jvi.asm.org/
reference 32). Cleavage during DNA maturation occurs in DR, which is part of element "a" (Fig. 9aand b). Asaresult of the variable orientations of UL and
U.
components, cleavage of the concatemers can occur ateach"a"sequence in any DR. This is in contrast to the less-complicated genome organization of BHV-1, in which cleavage for virus maturation occurs only inacertain c element. Itisof interest that the cleavage of HSV-1 is notmediated by DR itself but by other internal elements of "a" (42). Thesesequence-unspecific cleavages require a complete
Uc
sequence, whereas Ub can be deleted. Additional data support this observation. In amplicon constructs (36, 37, 43), intact or deleted "a" sequences of HSV-1 canbetested forcleavage before packaging into defective interfering particles. Deisset al. (9) have shown with such amplicon constructs that deletions inUc
spanning theAn
stretch inactivated the cleavage function, whereas Ub could be omitted withoutinfluencing the cleavage and packaging efficiency. These
findings with HSV-1 support ourhypothesis that the cleav-age site bearing
P
element does not mediate cleavagespec-ificity, whereas the
An
stretch does.In BHV-1, it is likely that at the UL-IR junction an
An
stretch as part of the recognition site of the terminase has been replaced by an a element during evolution (Fig. 6). This replacement eliminates not only the
An
stretch in the UL-IRjunction but also a potential secondary hairpin structure of the single a element. Because of the partial duplication
a+at,
the loop is either larger or dislocated with respect to theputative cleavage site. The functional importance of thisregion is underscored by a comparison of the nucleotide sequencesofthejunctionUL-IRof BHV-1 and VZV (Fig. 8).
Although bothgenomes displayin general the same arrange-ment
of
UL,
IR'
US,
andTR,
theUL
segment
of VZV is bracketed by two inverted repeats TRL and IRL spanning 88.5bpatthe left andright ends of UL. Thus, theAn
stretch, as well as the a element of VZV, is duplicated in inverseorientationatthejunction UL-IR. Consequently, two identi-cal putative recognition sites and two cleavage sites occur, which should give rise to inversion of UL relative to IRS
providedthat both cleavage sites are equivalent. VZV shows this predicted inversion of UL although the two isomers do not represent equimolar proportions (6).
It is noteworthy that two reports emphasize the universal
function oftheterminase at the cleavage-recognition sites of
herpesviruses. With amplicon vectors containing a HSV-1
origin of replicationand "a" sequences of two herpesviruses onthesamemolecule,Spaete and Mocarski (38) showed that theHCMV"a" sequence is cleaved if HSV-1 helper virus is present. Although HSV-1 and HCMV share no detectable sequencehomology (17), the "a" sequences of HCMV were
recognizedby the terminase of HSV-1. By applying the same
approach, similar findings were reported with the "a" se-quenceof HSV-1 and a related herpes simplex virus type 2 strain as a helper to enable cleavage-packaging of tandemly arranged multimers of HSV-1 sequences in defective parti-cles (37).
Site-specific inversion in the HSV-1 genome requires the "a" sequence (27, 28) and involves both cleavage and
joining of the inverting sequences (4). In HSV-1, unique sequences bracketed by two "a" sequences invert,
contrib-uting to the generation of four equimolar genomic arrange-ments in herpes simplex virus. In contrast, the BHV-1 genome contains only two genomic isomers. It is obvious that the inverting short segment of BHV-1 in virion DNA
molecules isbracketed by two inversely orientedaelements and the adjacent sequences of the -y elements (Fig. 7). In
contrast, the noninvertable UL segment isbracketed bytwo at elements only. This observation leads to the
assumption
that the two yelements maycontribute signal(s) required for
the inversion of
Us
ofBHV-1.Itis not yet clear whatrolethe "a" sequences perform in
the inversionofULand
Us
of HSV-1. Invivorecombinants of HSV-1 carrying an additional complete or deleted "a" sequence in UL give rise to different results. Whereas Varmuza and Smiley (42) reported low levels of inversion inthese mutants, Mocarski and Roizman (27, 29) have de-scribed similar mutantswhich are active in inversion.
Addi-tionally, deletions of the internal DRs of "a" reduce inver-sion significantly.
Our hypothesis that the
An
sequences ofherpesvirus
genomes areessential for the maturation ofreplicative-form
DNAand theformation ofthegenomic isomerscanbe tested by the introduction of mutations into a and,B elements and
into the
An
stretch of BHV-1. The useof BHV-1, which has alesscomplicated genome structure thanHSV-1, is likelyto aid in the detailed understanding of these recombinational events common to many herpesviruses.ACKNOWLEDGMENTS
B. Sugden, V. Baichwal,and J. Knutsonare gratefully acknowl-edged for stimulating discussions and critically reading the manu-script. We thank Kristen Luick for manuscript preparation and Terry Stewartfor photographic assistance.
This workwassupportedby grants BCT 0363 (H.-J.B.) and PTB 8352/BCT311A (H.L.) from theBundesministerium fur Forschung undTechnologie.
LITERATURE CITED
1. Bankier, A. T., W. Dietrich, R. Baer, B. G. Barrell, F. Colbere-Garapin, B. Fleckenstein, and W. Bodemer. 1985. Terminal repetitive sequences in herpesvirus saimiri virion DNA. J. Virol. 55:133-139.
2. Ben-Porat, T., A. S. Kaplan, B. Stehn, and A. S. Rubenstein. 1976. Concatemeric forms of intracellular herpesvirus DNA. Virology69:547-560.
3. Ben-Porat, T., F. J. Rixon, and M. L. Blankenship. 1979. Analysis of thestructureofthe genome of pseudorabies virus. Virology 95:285-294.
4. Chou, J., and B. Roizman. 1985. Isomerization of herpes sim-plexvirus 1genome: identificationof thecis-acting and recom-binationsiteswithinthedomain ofthe asequence. Cell 41:803-811.
5. Chowdhury, S. I., G. Kubin, and H. Ludwig. 1986. Equine herpesvirus type 1 (EHV-1) induced abortions and paralysis in a Lipizzaner stud: a contribution to the classification of equine herpesviruses. Arch. Virol. 90:273-288.
6. Davison, A. J. 1984. Structure of the genome termini of vari-cella-zoster virus. J. Gen. Virol. 65:1969-1977.
7. Davison, A. J., and J. E. Scott. 1986. The complete sequence of varicella-zostervirus. J. Gen. Virol. 67:1759-1816.
8. Davison, A. J., and N. M. Wilkie. 1981. Nucleotide sequences of the joint between the L and S segments of herpes simplex virus types 1 and 2. J. Gen. Virol. 55:315-331.
9. Deiss, L. P., J. Chou, and N. Frenkel. 1986. Functional domains within the a sequence involved in the cleavage-packaging of herpes simplex virus DNA. J. Virol.59:605-618.
10. Deiss, L. P., and N. Frenkel. 1986. Herpes simplex virus amplicon: cleavage of concatemeric DNA is linked to packaging and involves amplification of the terminally reiterated a se-quence. J. Virol. 57:933-941.
11. Engels, M., C. Guiliani, P. Wild, T. M. Beck, E. Loepfe, and R. Wyler. 1986/1987. The genome of bovine herpesvirus 1 (BHV-1) strains exhibiting a neuropathogenic potential compared to known BHV-1 strains by restriction site mapping and cross-hybridization. Virus Res. 6:57-73.
12. Gregersen, J.-P., G. Pauli, and H. Ludwig. 1985. Bovine
on November 10, 2019 by guest
http://jvi.asm.org/
REPLICATIVE-FORM DNA OF BHV-1 1363 pesvirus 1. Differentiation of IBR- and IPV viruses and
identi-fication and functional role of their major immunogenic compo-nents. Arch. Virol. 84:91-103.
13. Hammerschmidt, W.,H. Ludwig, and H.-J. Buhk. 1986. Short repeats cause heterogeneity at genomic terminus of bovine herpesvirus 1. J. Virol. 58:43-49.
14. Hanahan, D. 1983. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 166:557-580.
15. Harper, L., J. Demarchi,and T.Ben-Porat. 1986. Sequenceof the genome ends and of the junction between the ends in concatemeric DNA in pseudorabies virus. J. Virol. 60:1183-1185.
16. Henry, B. E., R. A.Robinson, S. A. Dauenhauer, S. S. Atherton, G.S.Hayward, andD.J. O'Callaghan. 1981. Structure of the genomeofequine herpesvirus type 1. Virology 115:97-114. 17. Huang, E.-S., and J. S. Pagano. 1974. Humancytomegalovirus.
II. Lack of relatedness to DNA of herpes simplex I and lI, Epstein-Barr virus, and nonhuman strains of cytomegalovirus. J. Virol. 13:642-645.
18. Jacob, R. J., L. S.Morse, and B. Roizman. 1979. Anatomy of herpes simplex virus DNA. XII. Accumulation of head-to-tail concatemers in nuclei of infected cells and their role in the generationof the four isomeric arrangements of viral DNA. J. Virol. 29:448-457.
19. Ludwig, H. 1983. Bovine herpesviruses, p. 135-214. In B. Roizman (ed.), The herpesviruses, vol. 2. Plenum Publishing Corp., NewYork.
20. Matsuo, T.,M.Heller, L.Petti,E.O'Shiro,andE. Kieff. 1984. Persistence of the entire Epstein-Barr virus genome integrated into human lymphocyteDNA. Science 226:1322-1325. 21. Maxam, A. M., and W. Gilbert. 1977. A new method for
sequencing DNA. Proc. Natl. Acad. Sci. USA 74:560-564. 22. Maxam,A. M., andW. Gilbert. 1980. Sequencingend-labeled
DNAwithbase-specificchemicalcleavages.MethodsEnzymol. 65:499-560.
23. Mayfield, J. E.,P.J. Good,H.J.VanOort,A.R.Campbell,and D.E. Reed. 1983. Cloning and cleavage sitemappingof DNA from bovineherpesvirus1(Cooperstrain).J.Virol.47:259-264. 24. Messing, J., R. Crea, and P. H. Seeburg. 1981. A system for
shotgunDNAsequencing. Nucleic Acids Res. 9:309-322. 25. Metzler, A. E., H. Matile, U. Gassmann, M. Engels, and R.
Wyler. 1985. European isolates of bovine herpesvirus 1: a
comparison of restriction endonucleasesites, polypeptides and reactivity with monoclonal antibodies. Arch. Virol. 85:57-69. 26. Misra, V.,L. A.Babiuk,andC. L.Q.Darcel. 1983.Analysisof
bovineherpes virus-type 1isolatesbyrestriction endonuclease fingerprinting. Arch. Virol. 76:341-354.
27. Mocarski,E. S.,and B. Roizman. 1981. Site-specificinversion sequence of the herpes simplex virus genome: domain and structural features. Proc.Natl. Acad. Sci. USA 78:7047-7051. 28. Mocarski, E.S.,and B. Roizman. 1982. Herpesvirus-dependent
amplification and inversion ofcell-associated viral thymidine
kinase gene flanked by viral a sequencesand linked to an origin of viral DNA replication. Proc. Natl. Acad. Sci. USA 79:5626-5630.
29. Mocarski, E. S., and B.Roizman. 1982. Structure and role of the herpessimplex virus DNA termini in inversion, circularization andgeneration of virion DNA. Cell 31:89-97.
30. Poffenberger, K. L., and B. Roizman. 1985. A noninverting genome ofaviable herpes simplex virus 1: presence of head-to-taillinkages in packagedgenomes and requirements for circu-larization after infection. J. Virol.53:587-595.
31. Rigby,P.W. J., M.Dieckmann, C. Rhodes, and P. Berg. 1977. Labelling deoxyribonucleic acid to highspecific activity in vitro bynick translationwith DNA polymeraseI. J. Mol. Biol. 113: 237-251.
32. Roizman, B. 1979. The structure and isomerization of herpes simplex virus genomes. Cell 16:481-494.
33. Roizman, B. 1982. Thefamily Herpesviridae. General descrip-tion, taxonomy, and classificadescrip-tion, p. 1-23. In B. Roizman (ed.), Theherpesviruses, vol.1.PlenumPublishing Corp., New York. 34. Sanger, F., A. R.Coulson, B. G. Barrell, A. J. H.Smith, and B. A. Roe.1980.Cloninginsingle-stranded bacteriophage as an aidtorapid DNAsequencing. J. Mol. Biol. 143:161-178. 35. Southern, E. M. 1975. Detection of specific sequences among
DNAfragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517.
36. Spaete, R.R., and N.Frenkel.1982. Theherpes simplex virus amplicon: a new eucaryotic defective-virus cloning-amplifying vector. Cell 30:295-304.
37. Spaete,R.R., and N. Frenkel. 1985. Theherpessimplex virus amplicon: analysis of cis-acting replication functions. Proc. Natl. Acad. Sci. USA82:694-698.
38. Spaete,R. R.,and E.S. Mocarski. 1985. The asequenceofthe cytomegalovirus genome functionsas acleavage/packaging sig-nal for herpes simplex virus defective genomes. J. Virol. 54: 817-824.
39. Sullivan, D. C., S. S. Atherton, J. Staczek, and D.J. O'Cal-laghan. 1984. Structure of the genome ofequine herpesvirus type3. Virology 132:352-367.
40. Tamashiro, J. C.,D.Filpula,T.Friedmann,and D. H.Spector. 1984. Structure of the heterogeneous L-S junction region of human cytomegalovirus strainAD169 DNA. J. Virol. 52:541-548.
41. Tamashiro, J. C., and D. H.Spector. 1986. Terminal structure
andheterogeneity in humancytomegalovirus strainAD169. J. Virol. 59:591-604.
42. Varmuza,S.L.,andJ.R.Smiley. 1985.Signalsforsite-specific cleavage of HSV DNA: maturation involves two separate cleavageevents atsites distaltotherecognitionsequences.Cell 41:793-802.
43. Vlazny, D. A., and N. Frenkel. 1981. Replication of herpes simplex virus DNA:localization ofreplication recognition sig-nals within defective virus genomes. Proc. Natl. Acad. Sci. USA 78:742-746.