Vol. 58, No.2 May
0022-538X/86/050513-09$02.00/0
Copyright© 1986, American Society forMicrobiology
Cloning
and
Characterization
of
oriL2, a
Large
Palindromic
DNA
Replication Origin
of
Herpes
Simplex Virus Type
2
DANIEL LOCKSHONAND DENISE A. GALLOWAY* FredHutchinson Cancer Research Center, Seattle, Washington98104
Received 2 December1985/Accepted7February 1986
Anorigin ofreplication within the long unique sequence of herpes simplex virustype2designated
OriL2
has been identified in aposition homologous to its type 1 counterpart,oriLl, between map coordinates 0.398 and 0.413. The difficulties encountered inpreviousattemptstoclone bothOriL2
andOriLl
in anundeletedform weresurmountedby (i)minimizingthe growth of the host
Escherichia
col,(ii)using a recBCsbcBE.colihost, and (iii)purifyingthefull-length plasmid from deleted forms byusing a novel method which exploits theabilityof apalindrome-containing plasmidtoadoptacruciformconformation,therebydecreasingitssupercoiling.Ina previously developed assay for functional originactivity,
OriL2 was localized to a 241-base-pair ApaI-SstIIfragment. DNAsequenceanalysisrevealed a 136-basepair,almostperfect palindrome. Comparisonwith
OriLl
showed a very high degree of conservation: the two origins differ in only 16 of the 144-base-pairOriLl
palindromic region. Mostsignificantly, thedifferencesbetweenOriLl
and OriL2 mainlyoccur inpairsso as togenerallypreserve thepotentialfor intrastrand basepairing.ThecentralregionofOriL2ishomologouswiththe shorter palindromic structures detected in origins located within the repetitive sequences of the short component ofherpessimplexvirus type 1or2.
Initiation of replication of herpes simplex virus (HSV)
DNA can occur atmultiple, widely separated loci. Although this wassuggested originallyby electron microscopic
visu-alization of the replicatinggenome (9, 14), subsequent
stud-ies which characterized the DNA sequences present in defective HSV type 1 (HSV-1) were more definitive
(re-viewed by Frenkel [8]). Thesedefective HSVs fall intotwo
classes. The class I defective genome consists of multiple tandem reiterations ofa fraction of the full-length genome
andalways containsasegmentof the "c"repeatunitof the Ssection of HSV-1DNA. That the classIdefectivegenome
bearsareplication originwasdirectly shown by
Vlazny
and Frenkel (38), who demonstrated that cotransfection of monomer-length class I defective DNA, generated byre-striction ofthe defective genome, with full-length HSV-1
DNA as helper resulted in the replication ofthe defective
DNA and the regeneration ofits tandemly repeated form.
Origin
activity within the "c" sequence was subsequentlyconfirmed by Mocarski and Roizman (22). Stow(35) delim-ited this so-called
oris
to a 995-base-pair (bp) restriction fragment,andStow and McMonagle(37)subsequently iden-tified amere 90-bp segmentwhich sufficed for origin func-tion.Inthoseexperimentsarecombinantplasmid containingthe
oris
segment was shown tobeamplified after itstrans-fection into cells which were subsequently infected with
HSV-1. Since
oris
resides in a repeat unit, each HSV standard genome contains two copies (or, in the case of HSV-2, four copies [43]) of this replication origin.Class II genomes contain instead a minimum of 3.4
kilobases(kb) from aboutthemiddle oftheULregion of the HSV-1genome (33).Attempts to clone this region have been
frustrated, however,by its markedtendency to suffer
dele-tions when grown in Escherichia coli (33, 41, 42). When a
plasmid containing the deleted form of this region of the genome is transfected into cells along with HSV-1 DNA,
defectivegenomes indeed arisefromthe plasmid, but their
*Correspondingauthor.
deleted region has been restored, presumably by recombi-nation with the helper HSV-1 DNA (33). This suggests that the deleted region contains the origin ofreplication, oriL, of the class II defective genome. GrayandKaerner (10), after mapping the region of an HSV-1 (strain ANG) class II defective DNA which deletes in E. coli, sequenced this region of the noncloned defective DNA. Similarly,Weller et al. (42) mapped the deleting region of the nondefective HSV-1 (strain KOS) genome to a 425-bp BstEII-BamHI
fragment. In addition, Weller et al. were successful in cloning the 2.3-kb BamHI-V fragment, which contains the 425-bp fragment, in a yeast vector in an apparently undeleted
form. They showed this construct, p1499, to exhibitorigin
function since after its transfection into animal cells, infec-tion withHSV-1 caused a dramatic amplification ofplasmid.
Bycontrast,deletedclones would notamplifyinthis assay. Both groups(10,42) found oriL of HSV-1 (hereafter referred to asoriLl) to contain a144-bp (i.e., two arms of 72 bpeach)
perfect palindromic sequence. As wasexpected from previ-ous demonstrations ofthe instability of palindromes in E. coli(2, 6, 12, 17, 21, 29), the sequences deleted from p1499 were always at least partially within its palindromic region (42). Thus, the palindrome contains elements required for oriLl function.
orisn
(23, 37, 40) andsubsequently ori2 aswell (43) werealso shown to contain palindromes. These are only about one-third as long the oriLl palindrome, but the middle
regions of allthree arehighly homologous. Knowledge ofthe sequence of oriL2 would completethispicture. Attempts by
Spaete and Frenkel tocloneoriL2inE. coliin anundeleted
form have been unsuccessful (34). The present report dem-onstratesthe construction of E. coliplasmids containingthe undeleted oriL2 sequence. We have also demonstrated clonedoriL2 to be functional. Finally, we have determined theoriL2 sequence. Asexpected,itishighly homologous to
oriLl,
yet severalimportant differencesareapparent as well.We argue that these differences between the two oriL se-quences further substantiate a physiological role for a cruciformconformation of the palindrome.
513
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS ANDMETHODS
Cloning procedures. AllDNA-modifyingenzymes (restric-tion enzymes, T4 DNA ligase, E. coli DNA polymerase I, Klenowfragment ofDNApolymerase,DNaseI) werefrom
New England Biolabs, Inc. or Bethesda Research
Labora-tories, Inc. and were used according to the supplier's instructions or established procedures (19). The cloning vectors used were Kosl (19), pKC7 (27), and pUC19 (45).
Electrophoresis was typically done through a 1% agarose
(Sigma, medium EEO) gel in submerged "mini"orhorizontal
(10by 17 cm) slabs. Preparative gelelectrophoresis(used to
purify theSall fragment of pDL017, to separate the deleted andfull-lengthforms of pDL701, andoccasionally forDNA
sequencing) employed 1% low-melting-point agarose (Be-thesdaResearchLaboratories,Inc.). Thedesired bandswere cut from the ethidium bromide-stained preparative gel,
melted at68°C, extracted with 1volume ofbuffer-saturated
phenol andthenwith 1volume of chloroform, and run over an Elu-Tip column (Schleicher& Schuell, Inc.). The DNA was theneluted from the column as suggested by the supplier and ethanol precipitated. HSV-2 DNA was prepared as
previously described (39). The cosmid pBglGJO was
constructed by ligation ofthe product of the partial BglII digestion of HSV-2DNAinto the Kosl vector.
The host for all plasmids except those otherwise noted was E. coli HB101 (4). E. coli JC9387 (recB21 recC22
sbcB15;J. R. Gillen,Ph.D. thesis, University of California, Berkeley,1974) wasprovided by G.Smith,andstrain JC811
(recB21 recC22 recF143 sbcB15; 15) was kindly sentby F. Stahl. E. coli cells were transformed by established
proce-dures (19). All E. coli growth, both on plates and in
liquid
culture, used L broth (19). After
transformation,
E. coli growthwasoften keptto aminimumbypermitting
growth ofthetransformed bacteriumonlyuntil it gaveacolonyof less than 0.5 mm in diameter. Most ofthe cells in this colony
were then removed into 1 to 11 ml ofampicillin-containing brothandgrownfor12 to 16h to astationary-phase culture, from which plasmidwasthenpurified.Plasmid wasprepared from either 1, 10, or 1,000 ml ofculture by the alkaline
detergent lysis procedure (19). Thorough
plasmid
purifica-tion was achieved using CsCl-ethidium bromide gradients
(19).
Replicon assays. Theprocedure of Stow andMcMonagle (37)wasclosely followed forrepliconassays except that the amount of cells used per transfection and the amount of
DNA per transfected cell were both scaled down.
Briefly,
35-mm-wide wells(sixwells perplate)ofsubconfluentBHK
cells,grown onDulbecco modifiedEagle medium
containing
5% fetal calf serum, were transfected with
equimolar
amounts ofplasmid (20 to 50 ng of plasmid per well) plusshearedcalfthymusDNA(2.4,ug perwell) ascarrierin 0.2 ml ofcalcium phosphate precipitate. Cells were refed 1 h aftertransfection and then shocked with dimethylsulfoxide 4hafter the DNA was added. At 6 hposttransfection, cells were either mock infected or infected with HSV-2 (strain 333) in Dulbecco modified Eagle medium containing 2% serum at amultiplicity of infection between 5 and 20. After incubation at37°Cfor 15h,all thecellsoftheinfected wells
exhibited cytopathic effects. The medium was then
aspi-rated, 0.5 ml of 0.6% sodium
dodecyl
sulfate-10 mM EDTA-10 mM Tris hydrochloride (pH 7.5)-0.5 mg of pronase per ml was added to the wells, and incubation was continued at 37°C for 4 h. The lysates were transferred to tubes and the wellswere rinsed with 0.5 ml of 10 mMTrishydrochloride(pH
8.0)-i
mMEDTA.NaClwasaddedto0.2M, and phenol and chloroform extractions were then
per-formed. The nucleic acid was ethanol precipitated and
adjusted to a concentration of 140 ,ug/ml, as determined
spectrophotometrically.
The "minifold" apparatus (Schleicher & Schuell, Inc.)
wasusedas atemplate for spottingDNAontonitrocellulose (BA85; Schleicher &
Schuell,
Inc.). A 50-,ul volume ofnormalizedlysate or control samplewascombined with 50
[L of 0.4 N NaOH ina96-well serocultureplate,and 10
RI
ofthiswas removed into90 [L of0.2 N NaOH for succesive 10-fold dilutions. Beforethe denatured samplewas
applied
tothe
nitrocellulose,
itwas neutralized with0.35 mlof 0.75MTris
hydrochloride
(pH7.5)-i
MNaCl. Thefilterwasthengently
rinsed, baked,
and probed with nick-translated pUC19, using established procedures (19). WhenpUC19-hybridizable
material was analyzed by gelelectrophoresis,
between 1 and 6
pLg
oflysate
DNAwasdigested
(or not) and electrophoresed through 1%agarose,afterwhichthegel
wasblottedbythe Southern method(32) ontonitrocellulose and
thefilterwasprobed asabove.
DNA sequence determination. The chemical
degradation
method of Maxam and Gilbert (20) and the method of
dideoxynucleotide
extension ofsynthetic
oligonucleotide
primers by the large fragment ofDNA polymerase using a
closed circular DNA
template (5, 30)
were both used todetermine DNA sequence. The sequenceofboth strandsof
the
326-bp
segmentspanning
theStuI-SstII sites(Fig. 1)
wasdetermined,
and anydiscrepancies
wereconvincingly
re-solved
by
additionalsequencing
runs. Thefollowing
twofragments were analyzed by the chemical method:
(i)
pDL601, linearized at the KpnI site in the
polylinker
se-quence of its vector, was 3'-end labeledusing
terminal tranferase(New
England
NuclearCorp.)
andcordycepin
[a-32P]triphosphate
(New
England
NuclearCorp.),
and thelabeled insert was then
sequenced
afterbeing
releasedby
HindlIl
digestion
andpurified
on apreparative gel;
(ii)
after3'
end-labeling
of the SstIIfragments
ofpDL601
using
terminal transferase and
cordycepin triphosphate,
theStul-SstII
fragment
was madeby
BamHIdigestion
and thenpurified
on apreparative
agarosegel.
Thefollowing
threedeterminations
using
theprimer
extension method were done:(i)
pDL601washybridized
withd(GTAAAACGACG-GCCAGT) (New England
Biolabs, Inc.);
(ii) 701A,
thefull-length
subpopulation
ofpDL701 (see
Fig. 3),
washy-bridizedwith the "reverse
sequencing primer"
d(AACAG
CTATGACCATG) (New England
Biolabs);
and(iii) 701B,
the deleted
subpopulation
ofpDL701 (see
Fig. 3),
washybridized
with the same reverseprimer.
RESULTS
Cloning of
OrlL2
in an undeleted form. Incloning
oriL2,
difficultiescomparable
tothoseseen whenoriLl waspropa-gatedinE.coli(33, 41, 42)were
anticipated. However,
in the presentstudy, problems onlyaroseafter extensivesubclon-ing.Figure1 illustrates theseriesof
subcloning
stepswhichpurified
oriL2 from thesurrounding
HSV-2 sequence.Be-cause the HSV-1 and HSV-2 genomes are known to be
colinear,
oriL2wasanticipated
tolie within the 0fragment
ofBglII-digested
HSV-2.Digestion
of cosmidpBglGJO
withBglII
andligation
of theproduct
intotheBglII
siteofpKC7
gave
pBglO.
Comigration
of the authentic4,650-bp BglII
0fragment
ofHSV-2(Fig. 2,
lane2)
with this insert(Fig.
2,
lane3,lower
band)
indicates thatnodetectabledeletionhad occurred. Infact, however,
afterpurification
of the whole insert frompBglO
anddigestion
withSalI(Fig. 2,
lane5),
asmallamountof deleted
fragment
was seeninadditiontotheon November 10, 2019 by guest
http://jvi.asm.org/
D RP G J 0 C N I H M L K
119 II
"I
IConstruct
nRnl C1
Insert size(kb)
Tendency vector to delete 34 KosI N.D.
PvuI[ _
tUl
HindM SalI StuI Sst1 Pst I SalBgIIE
I
S5I
SalIKpnISolI 'ApaI
pBgl 0 4.65 pKC7 verylow
pDL017 2.28 pKC7 verylow
pDL506 1.68 pUCl9 low
pDL601 1.40 pUC19 low
pDL701 0.33 pUC19 moderote pDL411 0.25 pUC19 N.D.
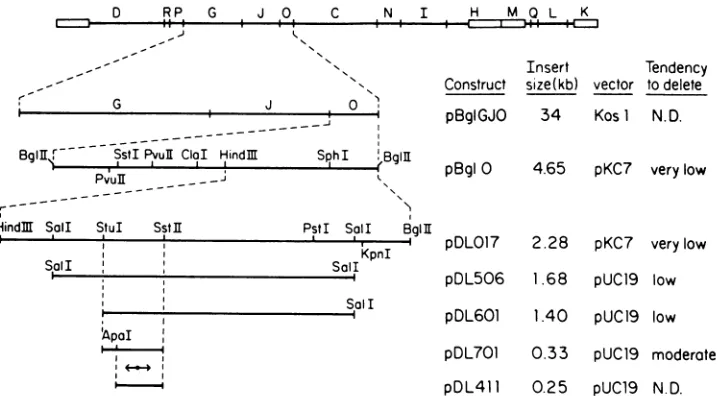
FIG. 1. Restrictionenzymecleavagemapofthe regionsurrounding oriL2. ThetoplineportraystheBglII fragments of HSV-2 (7),and the
repeatedsections of the L and S regionsareindicatedby boxes. The lines belowportraythesuccessive subcloneswhich containoriL2.The
tendency for the subclonestodelete thepalindrome, aqualitativeassessmentdescribedin the text, issummarizedintherightward column. ClaI,PvuII, SstI, KpnI, StuI,Sall, HindIII,andSphIcleave the entireBglII0fragment onlywhereindicated.PstIcleavesthepDLO17 insert
only where indicated, but has additional sites in therestofBglII 0.SstIl cleavesthepDLO17insertatleast fourtimes,butonlytheonesite whichwasmappedis shown. TheClaI site is cleavable in pBglOgrownindam, butnotdam',E.coli. The ApaI sitewasdeduced from DNA
sequence.The156-bppalindromic regionisrepresented byadouble-headedarrow.
full-length, 1,680-bp internal Sall fragment. The unique Hindlll sitewithin theBglII 0fragmentwas usedtobisect the insertto give pDLO17 (containing the rightwardhalfof the BglII 0fragment) and pDLO18 (containing its leftward
half).Inanassaydescribedbelow, bothpBglOandpDLO17 contained a functional HSV DNA replication origin,
whereaspDLO18 didnot.TheSalI fragmentwithin the insert ofpDL017wasthenpurifiedandligatedintotheSalIsiteof pUC19, ahigh-copy-number vector,to givepDL506.
Unlike pBglO and pDL017, it was obvious that pDL506 suffered deletions. Some E. coli transformants from this construction yielded homogeneous populations ofplasmid from which between 60 and 150 bp of insert was deleted,
whileother transformants containedamixtureof deleted and
apparently full-length molecules. As lanes 7 and 8 ofFig. 2 show,thefraction of deleted material could be kept accept-ably small (<10%) by limiting the extent of E. coli growth before plasmid preparation. By comparing restriction pat-ternsofplasmid from fully deletedtransformants with those ofpDL506, the deletionswerealwaysfoundtomapbetween the unique StuI site and the SstII site portrayed in Fig. 1. Thus, the next subcloningstepinvolvedtheexcision of the smallerSalI-StuIsegmentfrom theinsert by digestionwith StuI andXbaI, the latterof which cleaves within thevector polylinker. Repairof theends by DNApolymerase (Klenow fragment) treatment and recircularization by DNA ligase produced pDL601.
pDL601 suffered deletiontoabout the sameextentasdid
pDL506, and again, the fraction of deleted material in its preparations could be lowered sufficiently (toless than 10% of thepopulation) by minimization of E. coli growth (Fig. 2, lane9). Thefragmentof inserton therightward side of the
StuI-SstIIsegmentwasthenexcisedby digestionwith SstII
and PstI, the latter of which cuts in the vector polylinker rightward of the Sall site, to produce pDL701. This
con-struct, however,showed an evenhigher tendency todelete. Even when its E. coli H13101 host was grown as little as
possible, approximately half of the population of plasmid
molecules was deleted (Fig. 2, lane 10). Since DNA
se-quence cannot be determined from such a heterogeneous
population, the deletion problem now had to be solved in otherways.
Theeliminationof deleted moleculeswasachievedbytwo different methods. Initially, the closedcircular deleted and full-length populations were purified from eachother on a
preparative agarosegel. The unfractionated plasmid prepa-ration ("701"; Fig. 3, top) contained, in additionto nicked circles, both a highly supercoiled fraction (comprised ofa
series oftopological isomers which, because of their high supercoiling, were notresolvableon thisgel)and afraction
containing the less highly supercoiled topological isomers. Thesetwofractionswereseparated by cuttingapreparative
gel throughwhichalargeamountof 701waselectrophoresed
togive701A and 701B (Fig.3, top). Releaseof the insertby BamHIandHindIII digestion (Fig. 3, bottom)showed701to containtwospecies ofinsert,whereas701A, the lesshighly supercoiled fraction,containedonlythelarger insert (shown laterto be undeleted) and 701B containedonlythe smaller, deleted form.The nicked forms whicharose during
fraction-ation also illustrated that 701A is largerthan701B (Fig. 3, top).
By changing the E. colihost strain, thedeletionproblem
was subsequently more easily solved. A recBC sbcC host
genotype has been shown to enhance palindrome stability (16, 25, 44). Another studyhas demonstratedthat, in addi-tion,aninactiverecFgenewillallowmaintainence ofalarge
palindrome (3). Whenpurified701A(Fig. 3, top) was
intro-duced into E. coliJC9387(recBC sbcB),mosttransformants, aftergrowthto at least the same extentas the E. colifrom whichthe701 preparationinFig. 3waspurified,containeda
high fraction (>90%) offull-length plasmid (the remaining transformants bore exclusively deleted material). In the
same experiment, transformants of E. coli JC8111 (recBC recF sbcB) had either too low a proportion offull-length
material (never more than 60%) or only deleted material (data not shown). Thus the palindromic sequence in oriL2
G J 0
58,
L =
IJLYI%vv%
- -j
Bgl]jr---
---.StIPvull
ClaI HindM SphI eBglllon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.129.489.72.270.2]
C---650 4370- 2320--2030'
HE
-
-t-.i 5-:
I-O. C) f
I-i
0 00
0n (i!
_i -j
-L... C-2
2-CIL C.) :L
1350
i 1080
-870 --600 -275
2 3 4
6
7
69
lullFIG. 2. Restriction enzyme analysis of oriL2 subclones. Lane 2, BglII-digested HSV-2 DNA. Lane 3, Intact insert ofpBglO,released byBglII digestion. Lane 4, Intact pBglO, after SalI digestion, gave the 1,680-bp Sallfragment of the insert in addition to two vector-insert junction fragments and a vector-derived fragment. After purification of the insert from pBglO, digestion with Sall (lane 5) gave the internal 1,680-bp SalI fragment and, in addition, a small amount of deleted SalI fragment migrating slightly faster than the full-lengthform.SalIdigestion ofpDL017(lane 6)again revealed the SalIfragment, but the small amount of deleted Sall fragmentwas obscured, asin lane 4, by the vector-derived band. Lanes 7 and 8, Two different preparations of pDL506 were digested with Sallto -release the full insert. The plasmid in lane 7 was prepared from extensively grown E. coli, whereas in lane 8, host growthwaskept toaminimum. Lane 9, Release of the complete insert of pDL601 (from minimally grown E. coli) by restriction with BamHI and HindIII, two enzymes whose sites reside in thevectorsurrounding thisinsert, providing the 1.4-kb insert andasmallamountof deleted material as well. Lane 10, Digestion of pDL701 (prepared from minimally grown HB101 host) withBamHI andHindIII gave the full-length 335-bp insert andacomparable amount of190-bp deleted insert. Fragment size markers areHindIII-digested X phage DNA (lane 1) andHaeIII-digested4X174RFform(lane 11).
appears to be more stable in strain JC9387 than in strain JC8111.
The DNA sequence analysis during the course of the
subcloning revealed a unique
ApaI
site within the 326-bp insert ofpDL701 and to theleftofthepalindromicsequence.The finalsubcloningstep wasthus the excision of this85-bp leftwardfragment by digestion withApal
andKpnl
(another polylinker site) to produce pDL411. In this construction, strain JC9387 served as the immediate host of theligation
product. Again, >90% of the plasmid preparation was full
length (data notshown).
Analysis of origin function in vivo. As was shownfirstwith simian virus 40 (24) and later with HSV-1 (35, 37), cloned
viral origins can drive the replication of their bacterial vectors when thetrans-acting viral factors are present in the
transfected cell. Using this method, analysis of a series of
BAL 31 exonuclease-generated deletants of cloned HSV-1 enabled Stow and McMonagle (37) to delimit sequences necessary forthefunction of
orisl.
We have employedthisrepliconassay to show that cloned oriL2 is functional.Inthe
simplest application ofthisprocedure(Fig. 4),HSV-2 infec-tionwasshown toamplifysometransfectedplasmidsbut not
others. Total DNA from BHK cells, transfected with a
plasmidandsubsequentlyinfected withHSV-2,wasspotted
onto a nitrocellulose filter and probed for
plasmnid
vectorsequence. Therewas anenormousincreasein dotintensity,
uponinfection with HSV-2, of DNA fromcells transfected
with eitherpBglO, pDLO17, pDL601, pDL701, or pDL411
(Fig. 4).Incontrast, amuchsmallerincreaseindot
intensity
was observed when cells were transfected with pDLO18 (containingthe leftward half of BgiII 0) or pUC19. Thus,oriL2isprobably contained within the former plasmids, but notthelatter.
Thevector-hybridizable materialinthese lysateswasalso
tested formethylation of theN6 position of adenine in the sequenceGATC. Thispropertypermitsdistinctionbetween
DNA replicated in E. coli, which modifies this sequence using dam methylase, from that synthesized inaeucaryotic cell, which is apparently incapable of methylating (or demethylating) the N6positionof adenine(28).Digestion of
lysates with either MbcI, which cleaves only at
unmethyl-atedGATC,orwith DpnI,which cleaves thesame sequence only when methylated, serves to testthis property. Figure
SA examines lysates from pDL411-transfected cultures.
Lane 1 demonstrates the vector-hybridizable material from thepDL411-transfected andHSV-infected culturesto be of high molecular weight. This was expected, since earlier reports of replication of cloned
oris1
(35) and ofdefectiveHSV-1 (38) have shown the producttobeatandemrepeatof high molecular weight. Digestion with MboI (Fig. 5, lane 2) completely converted this slow-migrating material into frag-ments which displayed the same mobility as the digestion prQducts of authentic pDL411 (Fig. 5, lane 12). However,
digestion withDpnI had noapparenteffect (Fig. 5, lane 3). DpnIwasindeedactive, since DpnI digestion after addition of E. coli-derived pUC19 to the lysate (Fig. 5, lane 5)
produced bands characteristic of DpnI-digested pUC19(lane 10). Lane 6 of Fig. 5 contains undigested lysate from a culture which was not infected with HSV-2 after pDL411
transfection. As demonstrated by the dots of Fig. 4, far less vector-hybridizable material was apparent than in the HSV-2-infected counterpart (Fig. 5, lane 1). The mobility of the uninfected material corresponded to that of authentic pDL411 (lane 11), and none ofthe high-molecular-weight
material of lane 1 was seen. As expected, MboI digestion
(Fig. 5, lane 7) could not alter its mobility, whereasDpnI
Uncut
Born HI
HindITT
N: ie:c %irc'es
*SJpercoIle,d
1-3C;7l
S1ltP
FIG. 3. Separation of the full-length and deleted forms of pDL701. The upper panel contains undigested samples, and the lower shows samples digestedwithbothBamHIandHindlll, two enzymes whichcut oneitherside oftheinsert, therebyreleasingit. Lane701, Plasmid pDL701 preparedfrom minimallygrownstrain HB101host. The resultof excision ofalltopoisomers(i.e., exclud-ingthenickedcircles) except thehighly supercoiled forms froma
preparativegel of701is shown in lane 701A. Materialinlane 701B waspurifiedfrom ahighlysupercoiledbandof thesamepreparative gel.
.: .." , F, ., ,.;: .1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.98.266.69.257.2] [image:4.612.375.498.496.632.2]digestion (lane 8) converted it to fragments which corre-sponded in mobility to authentic DpnI-digested pDL411 (lane 12). Thus, Fig. SA demonstrates that the large amount of vector-hybridizable material which appears upon infec-tion of a pDL411-transfected culture with HSV-2 is indeed synthesized aftertransfection since (i) it has the character-istic high molecular weight and (ii) it is unmethylated on the N6position of adenine in the sequence GATC.
Can HSV-2 infection induce any replication of transfected DNA whichlacks HSV origin sequence? Thiswas apossible interpretation of the10-fold increase in dot intensity of either pDL018- orpUC19-transfected cultures. The methyl group assay was used to test for small amounts of replication of these transfected plasmids (Fig. 5B). Unlike the slow-migrating material seen with pDL411 transfection (Fig. SA),
transfection with pDLO18 (Fig. SB, lane 14) or with pUC19 (lane 23), followed by HSV-2 infection, gave
vector-hybridizable material which comigrated with input DNA. Moreover, this material was DpnI sensitive(Fig.5B,lane 15,
pDL018; lane 24, pUC19) and MboI insensitive (lane 17,
pDL018,lane25,pUC19).Thus,neitherpDL018 nor pUC19 wascapable ofreplication. Origin function istherefore quite specific: only when the transfected DNA bears a certain sequence is it capable of replicating after the trans-acting
factors are provided by HSV-2. The increase in
vector-hybridizable material upon HSV-2 infection of cells transfected withplasmids whichlackfunctional HSV origins
couldperhaps be attributed to an increasedpermeabilityof
cellsto residualcalcium phosphate-precipitated DNA.
SequenceofOrHL2. WhenpDL601 and 701A, the material
purified from pDL701 growth in HB101 (Fig. 3), were
subjectedtoboththedideoxy triphosphatechain termination
(5, 30) and the chemical modification (20) methods, the sequence ofthe 326-bp StuI-SstII segment was determined
(Fig. 6). The replicon experiment described aboveconfines
thesequenceimportant fororiL2functionto bases 86 to 326.
Thetwo armsofthepalindrome, which extend frombase 122
pBqlO p3iO, pl_018 pDL601 pX720 4'
+
10-fold
dilutiorns
1.0-foG!d
didutiors
...12 3 4 5
[image:5.612.319.557.74.275.2]pUCi CT L
FIG. 4. Hybridization, to vector sequence, of lysates ofcells transfected with oriL2plasmidsfollowedbyHSV-2ormock
infec-tion. Two wells of BHKcells were transfected withthe indicated
plasmids combined with carrier DNAor with carrier DNA alone
(CTRL). Afterdimethyl sulfoxide shock, HSV-2 infection (+) or
mockinfection (-), andovernightincubationat37°C,cells from 16 wellswerelysed, and DNAwasprepared. Tenfold dilutionsof the
lysateswere then spotted onto nitrocellulose. Inaddition, 10-fold dilutions ofvarious controlswerespotted.Column1, 2ngofpUC19 plus undiluted CTRL, + lysate; column 2, 2 ng ofpUC19 plus undilutedCTRL,-lysate; column3, 2ngof pUC19 only;column 4,
0.2,ugof HSV-2DNA;column5,2ngofpKC7.Thefilterwasthen
probedwithnick-translated pUC19.
A
1 2 3 45 6 7 8 90 1 I2 13
qm a
I
..xIt
4 .~~~~~~~4
_ _ do @4
I
B
14 15 16 17181,920 21 22 23 2425
*~~~~~~~~~.E
7')
FIG. 5. Digestion of lysates with MboI and DpnI. pDL411-transfected lysates (A) and pDL018 and pUC19 lysates (B) were analyzed. (A) Lanes 1 through 5 contain lysate from pDL411-transfected andHSV-2-infectedcells.Lane 1, Undigested; lanes 2 and 4,MboI-digestedlysatealone (lane 2) orlysate plus 0.2 ng of pUC19 (lane 4); lane 3 and 5, DpnI-digested lysate alone (lane 3)or lysate plus 0.2 ng ofpUC19 (lane 5). Lanes 6 through 8 contain lysatefrompDL411-transfected and mock-infected cells. A ninefold longer autoradiographic exposure of these three lanes is shown. Lane 6,Undigested; lane 7,MboIdigested; lane 8,DpnI digested. Lanes 9through 13 contain various markers. Lane 9, pUC19 (0.2 ng), undigested; lane 10, pUC19 (0.2 ng), DpnI digested; lane 11, pDL411 (0.3 ng), undigested; lane 12, pDL411 (0.3 ng) DpnI digested; lane 13, X phage DNA, HindIII digested and end-labeled with 32p. (B) Lanes 14 through 17 contain lysate from pDL018-transfected and HSV-2-infected cells.Lane14,Undigested;lane15, DpnIdigested; lane 16, lysate combined with 0.2 ng ofpUC19 and thendigestedwithDpnI; lane 17, MboIdigested. Lanes 18 and 19 contain pDL018-transfected, mock-infected lysate and are undi-gested (lane 18)orDpnIdigested (lane 19).Lanes 20through 22 bear various markers. Lane 20, pDLO18, undigested; lane 21, DpnI-digestedpDLO18;lane22,DpnI-digestedpUC19.Lanes 23through 25containpUC19-transfectedandHSV-2-infectedlysate.Lane23, Undigested; lane 24,DpnIdigested; lane 25,MboI digested.
to base 257, are denoted in Fig. 6 by brackets. Note the
absence of any intervening nonpalindromic sequence. The
ATrichness ofthe oriLl palindrome has been noted previ-ously (42)and isreadilyapparent fororiL2 as well. The37%
A+T content ofthe entire 326-bp segment is only slightly higher than the 31% A+T content of the entire HSV-2 genome. The 136 palindromic base pairs are 51% A+T,
while the 190 bp of flanking nonpalindromic sequence is
merely 28%A+T.The middle 96bp ofthepalindromehas an A+T content of 64%. Thus, as far as base composition is concerned, oriL2, likeitscounterpart inHSV-1,iscomposed of GC-rich domains which flank the AT-rich palindrome. ThishighAT contentis concentrated in the centralregion of thepalindrome.
The deleted subpopulation ofpDL701 grown in HB101, 701B(Fig. 3),wasalsosequenced usingthedideoxy triphos-phatechain termination method. It was found to beidentical
in sequence to 701A except for the deletion of bases 94
through 241 of Fig. 6. This directly demonstrates, as had been shown previously in cloning oriLl (42), that deletion occursatleast partiallywithin thepalindrome.
4- -r- -
-0 0
-AW&WOMMAL. A..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.84.277.460.604.2]CCTGCGGATG GCGGCCCAGA GCTGGGTATG CTCGGCCGGG GCGGCCGGTA TATGTACGGC 60 GTGCTGGGAG GGGCGGCGTC GGGCCCCGCC CACGGTCCGC CACGCCCCGC GCGTCATCGG 120 C[AGGGGGCGT GGTCGCCCTT CTAAAAAAAG TGAGAACGCG AAGCGTTCGC ACTTTGTCCT 180 AATAGTATH ATATTATTAG GACAAAGTGC GAACGCTTCG CGTTCTCACT TTTTTTAGAA 240 GGGCGGCCAC GCCCCCTJTTG ACGTCACGCT CACCCGGGCG GCCGGCCGCC CATAAGCGCG 300
GCCTGCCGGG CCGATAAAAA GAAACC 326
FIG. 6. DNA sequenceoftheinsertofpDL701.Onestrand,reading 5' to 3',of theHSV-2sequence contained inpDL701isshown. Each 68-bparmof the palindrome is enclosed inbrackets.ApaIcleaves between bases 85 and 86.
DISCUSSION
Cloning of oriL2. A 241-bp fragment of HSV-2 which
containsoriL2has beencloned.Weconcludethat nodeletion has occurred during the course of this construction (aside
from that inasmallfraction of the plasmid population)since
(i) the cloned oriL2 is functional in vivo and (ii) when the
clonedoriL2 sequenceiscompared to the sequenceof
oriLl
derived from viral (uncloned)DNA(10,42), thesesequences are highly homologous and colinear, with no indication of
sequence present inoriLl thatismissingin oriL2 (Fig. 7).
Deletion did pose certain problems, butthese were sur-mounted by minimizingthegrowthof the host, bypurifying deletedfrom undeleted plasmid, and by using abetterhost
strain. The problem is most simply solved by limiting the
growth ofthe E. colitransformant. If this is unsuccessful,
the use ofadifferenthost strain is necessary. Since oriL2 is
more stable in strain JC9387 (recBC sbcB) than in strain JC8111 (recBC recF sbcB), whereas the large palindrome from minute virus of mice is more stable in the latter (3),
both strains should be tested whencloning palindromes.
Thepurification of undeletedpalindrome-containing
plas-mid reported here could be used when the simpler
ap-proaches prove unsuccessful. This procedure (Fig. 3)
ex-ploits the ability of palindromes to adopt the cruciform
structure(21, 26). The numberofsupercoils is determinedin part by the number of primary helical turns in a closed
circular molecule of DNA. The extrusion of palindromic
sequenceintoacruciformconformation depletesamolecule
of
primary
helical turns. Thenegative supercoiling
oftheplasmid thereby decreases byanamountwhichdepends on
the size ofthe palindrome. In the case oforiL2, about 13
supercoils are removed by cruciform formation (136 bp of palindrome + 10.4bpperhelicalturn). Theaverage number
of supercoils in noncruciform pDL701 is about 18 (0.096 helicalturnsperbp x 3,100 bp x 0.06supercoilsperhelical
turn; 1). Crucifortn formation, which
presumably
occursonly during the course of plasmid purification, therefore causesasubstantialreductioninsupercoiling. Since
electro-phoretic mobilitydepends on the degree of supercoiling, the
plasmid which is capable of a decrease in supercoiling by virtue ofcruciform formation is separable from that which
lacks the palindrome.
Palindromic
sequences are known tobepoorlypropagatedinE.coli(2, 6, 12, 17, 21, 29). oriLof HSVwasfirst reported
to deletein E. coli by Spaete andFrenkel (33), who
exam-ined HSV-1. Weller. et al. (42) were subsequently able to
cloneoriL, butonly ina yeast vectorsincedeletionororiLl inE.coli provedto betoo severe.SinceoriLl
cloned
in yeast was functional in a replicon assay similar to the onede-scribed here, it appears that at least part of the plasmid preparation used was undeleted (42). Previous attempts to
maintain
oriL
of HSV-2 in E. coli havebeen unsuccessful. Spaeteand Frenkel(34) indicatethat theSallfragment
(Fig.1,this paper) suffers "small"deletions. Haywardand Reyes (13) had earlier commented on such deletion in HSV-2, although they claimed thatthe ClaI site, which is 1,200 bp
away from the palindrome (Fig. 1, this paper), is always
within the 120 bp of deleted sequence. Since we find no evidenceof deletionaround theClaI site, andsincetheClaI siteis cleavablein plasmid fromdam butnot
dam'
E. coli(data not shown), it islikelythat theearlierstudy(14) didnot detect aClaIsite in theclonedDNAdue todammethylation
rather than to deletion. The present study, in successfully cloning full-length oriL2 in E. coli, allowsthepreparation of relatively largequantities ofanoriLclonecompared with the
small amountsobtainable fromayeast vector. Theability of
the present study, but not previous ones, to obtain full-length oriL in E. coli can be most likely attributable to
technical differences andperhapsto aninherentdifference in thebehavior oforiLl andoriL2 in E. coli, although the
high
degree of similarity between the oriLl and oriL2 sequences argues againstthe latterinterpretation.
What might account for the instability of
palindromic
DNA inE.coli?Onefact whichmayprove to be relevant to this problem is the marked decrease in stability of the smaller subclones of oriL2. ThiswasalsoseenwithoriLl(42).S2 Si
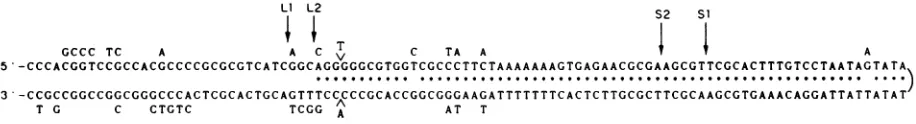
GCCC TC A A C T C TA A A
5 -CCCACGGTCCGCCACGCCCCGCGCGTCATCGGCAGGGGGCGTGGTCGCCCTTCTAAAAAAAGTGAGAACGCGAAGCGTTCGCACTTTGTCCTAATAGTATA
3 -CCGCCGGCCGGCGGGCCCACTCGCACTGCAGTTTCCCCCGCACCGGCGGGAAGATTTTTTTCACTCTTGCGCTTCGCAAGCGTGAAACAGGATTATTATAT
T G C CTGTC TCGG AA AT T
FIG. 7. Palindromic regionof oriL2 and itscomparisonwithoriLl. Onestrandof the oriL2 sequence(bases89through290 ofFig. 6) is portrayed inasnakelikefashiontoillustratethepotential for intrastrandbasepairing. The A and Tontherightwardendsofthetwolinesare
adjacentinitssequence. Eachasteriskdenotes apotential intrastrandbasepair of oriL2.The basesoforiLl(42) whicharedifferentfromoriL2
areindentifiedbyletters above orbelowthe lines oforiL2 sequence. Forinstance, oriLl containsanAinsteadof G in the fifthpositionfrom therightendofthe topline.The caratsindicatethat the A and T are inserted inoriLl relativetooriL2. Theextentof thepalindromes of
oris1,
oriS2, oriL2, and oriLl are indicatedbyarrows labeledSi,S2, L2,and Li,respectively.on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.86.544.608.670.2]HSV-2
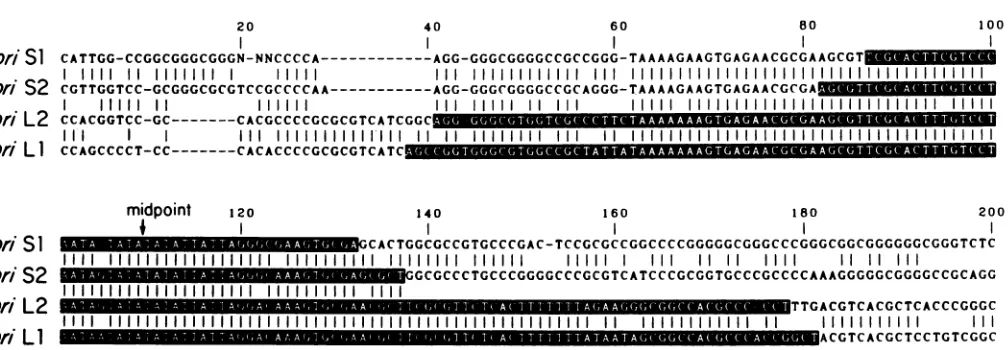
20 40 60 80 O1
Orn SI CATTGG-CCGGCGGGCGGGN-NNCCCCA---AGG-GGGCGGGGCCGCCGGG-TAAAAGAAGTGAGAACGCGAAGCGT
ori S2
CGTTGGTCC-GCGGGCGT
CTCCGCCCCAA---AGG-GGGCGGGGCCGCAGGG-TAAAAGAAGTGAGAACGCGAI I I11 IIII II II IIH il II
ori
L2
CCACGGTCC-GC .CACGCCCCGCGCGTCATCGGC..ugu--orn LI CCAGCCCCT-CC.---CACACCCCGCGCGTCATC
0
midpoint 120 140 160 180 200
on, SI CACTGGCGCCGTGCCCGAC-TCCGCGCCGGCCCCGGGGGCGGGCCCGGGCGGCGGGGGGCGGGTCTC
11111111111111111111111 11111111111 11111 1 111 11i11 111 11 11111
ori S2 GCGCCCTGCCCGGGGCCCGCGTCATCCCGCGGTGCCCGCCCCAAAGGGGGCGGGGCCGCAGG
or0 L2 TTGACGTCACGCTCACCCGGGC
ori LI
1W
ACGTCACGCTCCTGTCGGCFIG. 8. FourHSV replicationorigins. Sequencesarealignedsothat thecentersof thepalindromes (notedas"midpoint") coincide. The palindromic regions areindicated with white lettering. To allow the optimal alignment ofthe sequences, gaps (indicated atdashes) are
inserted. Thus,for example, G (base 60)and T(base 61)areadjacent intheoris1 sequence.Verticallines indicate that correspondingbases inadjacent sequencesarethesame. The
oris1
sequence is takenfrom reference23(bases 2345through2526; N indicates bases whichwereundetermined); theoriS2sequence is taken from reference43(bases711through896), theoriL2 sequenceshown is bases 90through 279 of Fig. 6,and the oriLl sequence is taken from reference42 (bases147through 338).
In the present study, pBglO and pDL017 were initially thoughttoberefractorytodeletion. In retrospect, however, it appears that a small fraction of pBglO prepared from extensively grown E. coli is deleted (Fig. 2,lane5).pDL506 andpDL601 haveahighertendencytodelete, and extensive E. coligrowth had tobe avoided togive a sufficiently high fraction of full-length material. Further subcloning to give pDL701 caused an even higher deletion rate. Presumably
pDL411 tends to delete to a similar, or perhaps greater extent than does pDL701, although this was not tested.
Unfortunately,nosimplewaytoquantitatethisphenomenon is apparent. Theincrease indeletion tendency withsmaller
insert size is not due simply to a change in plasmid size. When the Sall fragment of pDLO17 was ligated into pKC7 (5.8 kb) instead ofpUC19 (2.7
kb),
theproducthad asgreat atendencytodeleteasdidpDL506 (datanotshown). Thus,it appears that some feature of the surrounding HSV se-quence has a stabilizing effect on palindromes in E. coli. Examination of these flanking sequences would be most
interesting if other natural eucaryotic palindromes could be shown to actsimilarly inthisregard.
Features of theOrlL2 sequence. The most salient featureof oriL2-indeed, the feature which prevented it from being easily cloned-is its 68-bppalindrome. Thisis illustrated in
Fig. 7 by noting with asterisks the 66 potentialintrastrand base pairs. Note that aside from two mismatches, the
palindromeisperfect, i.e.,therearenobases whichserveto
interruptthe register ofthetwo arms.
Figure7also illustratesthedifferences between oriL2 and
oriLl,
notedaslettersabove and belowtheportrayal oforiL2sequence. The 16differences whichoccurwithin the 144-bp
palindromic region (rightward ofthearrowlabeled
Li
inFig. 7)areoftwotypes: those in oriL2which eliminate potentialintrastrand basepairs, and those whichoccurin pairs so as topreservethe potentialof intrastrand basepairing. Ten of the16differences fall into this latter group of paired changes, as far higher fraction than should occur if there were no selection pressure to maintain the relationshipbetween the twoarmsofthepalindrome. Fortwochangestoconstitutea
pairwisechange, theymust notonly occuratcorresponding
positions onthetwo arms, butthey must also be alteredto the correct base as well. Four of the pairs of changes are
simply thesubstitution of Gand CforAand T or viceversa. The fifth pair of changes, the insertion ofTand Ain oriLl
relativetooriL2,is
particularly striking.
Thus,despitethe 16differences in the oriLl and oriL2palindrome, the twodiffer by only sixin thenumber ofpotential intrastrand basepairs.
At least two functions ofpalindromes can be envisioned
(21). It is possible that the twofold rotational symmetry inherent in a DNA palindrome is important only for its interaction with a dimeric protein complex which also
ex-hibits twofoldsymmetry. Eachhalf ofthedimerwould thus
recognize one arm of the palindrome. To explain the high degree of
perfection
ofthepalindrome, theprotein complexwould have to be atleast as long as thepalindrome, binding
to it veryintimately soas torecognize all the bases over its
entire length. Ifthis were notthe case, there would be no
basistoexplaintheconservation of
palindromic perfection.
Considering
the extreme structural demands which thismodel would placeon suchahypothetical protein complex,
we consideritunlikely.
Inthe other model, the palindrome exists because of its ability toadoptacruciform conformation suchthatthetwo arms of the palindrome base pair with each other in an
intrastrand fashion. This is suggested merely by the
exist-enceofthese large perfect palindromes. Thecomparison of
thetwooriL sequencesevenfurthersuggeststhis model:the
differences, occurring mainlyin pairs, serveto maintainthe
potential for intrastrand base pairing. We thus consider it
likelythat the oriLpalindromebecomesacruciformduringat least partofthe virallifecycle. Cruciform formation iswell knowntobe influencedbyDNAtorsional strain(11, 21,26),
and thisiscertainlya
plausible
mechanism forahypotheticaloriLstructural transition. It should benoted, however, that there is no evidence for such strain in HSV-1 in vivo as determined by the rate oftrimethylpsoralen photobinding (31).This doesnoteliminate thepossibleexistenceofasmall class ofreplicating viral genomes which have unrestrained supercoils, ashas beenseen in simian virus 40 (18).
Itisunlikelythat the basepairingin thecruciformcould be maintained all the way to thecenter of thepalindrome, yet thereisno nonpalindromic sequencebetween itstwo arms.
Interestingly, this central region is not simply palindromic
but is, moreover, an alternatingAT sequence. Perhaps the VOL.S58,1986
I
I
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.57.561.64.239.2]unique sequence in this region serves a function different from that of the rest of the palindrome. Its AT composition makesiteasily meltable, and the alternating sequencecould
be necessary to allowthis region to melt as easily aspossible
(11) to allow the initiation of cruciform formation or of
replication itself. Stow (36), in fact, has shown the center of
the
oris1
palindrome to be essential fororigin function.Figure 8 compares the four HSV ori sequences. The
relationship between oriLl and oriL2 has been discussed above. Whittonand Clements (43) havepreviously discussed
the relationships between
oris1
andoriS2.
As reiterated in Fig. 8, these two sequences arevirtually identical. Welleretal. (42) have discussed the relationship between oriLl and
orisl,
and these are notdirectly comparedinFig.8. Becauseofthe close similarity between oriLl andoriL2and between
oris1
andoriS2,
general comparisons between oriL andoris
can be made. As noted previously (42), the sequences
leftward ofthe centersofthe oriLand
oris
palindromesarehighly homologous, whereas rightward of this center, the
homology extends only for about 25 bases. Thus, the
oris
palindrome is quite similartothat oforiL, but isonly about
25 bases long.
Acomparison of
oris
andoriL
suggeststhat notall of themoreextensive palindrome oforiLisrequired for replication. After all, since
oris1
is functional with merely a 23-base palindrome(37), and 47 or soadditionalpalindromicbases in oriL should be nonessential. Furthermore, since Stow andMcMonaglehave shown that aplasmid (Sl1 in reference 37)
which contains 298 bp of HSV-1 sequencerightward of base
51 (Fig. 8) is afunctional
oris1
replicon, whereas aplasmid (S13 in reference 37) whoseHSV-1 insert extendsrightward from base 84(Fig. 8) isnonfunctional, a sequenceessentialfor
oris5
function resideswithin the sequencehomologoustothe 25ththrough the 57th base oftheleftwardarmoftheoriL palindrome (bases51 through 83 of
Fig.
8). In otherwords,
asequencewhich isessential fororis
function lies outside of its palindrome, while itshomolog in oriLiscontained withinthe oriL palindrome. If
oris
and oriLfunctionsimilarly,
anassumptionatthis
point
basedsolely
ontheirhomology,
thisagain suggests that the outer
region
ofthepalindrome
isrequired forsomefunction otherthan
origin activity.
A role for the oriL palindrome in the control ofreplication
ortranscription thus becomes a
tempting possibility.
If this viewwere correct, the outerregion ofthe oriL2palindrome
should be superfluous with respectto
origin
functionin therepliconassay. We arecurrently
examining
thesignificance
ofvariouselementswithin theoriL2regionfororiginactivity.ACKNOWLEDGMENTS
We thank Jim Wallace and Margaret Swain for help with the computerassisted analysis of the DNA sequence, Toni Higgs for typing, and Paul Su for artwork.
This study was supported by Public Health Service grants CA35568 andCA26001from the NationalCancer Institute.
LITERATURE CITED
1. Bauer, W. R. 1978. Structure andreactions ofclosedduplex DNA. Annu. Rev. Biophys. Bioeng. 7:287-313.
2. Behnke, D., H. Malke, M. Hartmann, and F. Walter. 1979. Post-transformational rearrangement of an in vitro recon-structed group-A streptococcal erythromycin resistance plas-mid. Plasmid2:605-616.
3. Boissy, R., and C. R. Astell. 1985. An Escherichia coli recBCsbcBrecFhostpermitsthedeletion-resistantpropagation of plasmid clones containing the 5' terminal palindrome of minute virus of mice. Gene35:179-185.
4. Bolivar, F., and K. Backman. 1979. Plasmids of E. coli as
cloningvectors.MethodsEnzymol.68:245-280.
5. Chen,E.Y.,and P. H.Seeburg. 1985.Supercoilsequencing:a
fast and simple method for sequencing plasmid DNA. DNA 4:165-170.
6. Collins, J., G. Volckaert, and P. Nevers. 1982. Precise and nearly-preciseexcision of the symmetricalinverted repeats of Tn5;commonfeatures ofrecA-independentdeletioneventsin Escherichiacoli.Gene 19:139-146.
7. Cortini, R., and N. M. Wilkie. 1978. Physical maps for HSV type 2 DNA with restriction endonucleases. J. Gen. Virol. 39:259-280.
8. Frenkel,N.1980.Defectiveinterfering herpesviruses,p.91-120. InA. J.Nahmias,W.R.Dowdle,andR. F.Schinazi(ed.),The humanherpesviruses. Elsevier, NewYork.
9. Friedmann, A., J. Shlomai, and Y. Becker. 1977. Electron microscopy of herpes simplex virus DNA molecules isolated from infected cellsby centrifugationinCsCldensitygradients. J.Gen. Virol. 34:507-522.
10. Gray,C. P. and H.C.Kaerner. 1984. Sequenceof theputative origin ofreplication in the ULregionofherpes simplex virus type 1 ANGDNA.J. Gen. Virol. 65:2109-2119.
11. Greaves,D.R.,R. K.Patient,and D. M.J. Lilley. 1985. Facile cruciform formation byan (A-T)34 sequence from aXenopus globingene.J. Mol. Biol. 185:461-478.
12. Hagan,C.E.,and G.J.Warren. 1983.Viabilityofpalindromic
DNA is restored by deletions occurring at low but variable frequency inplasmidsof Escherichiacoli. Gene 24:317-326. 13. Hayward,G.S.,andG. R.Reyes. 1983.Biochemical aspects of
transformation by herpes simplex viruses. Adv. Viral. Oncol. 3:271-306.
14. Hirsch, I., G.Cabral,M.Patterson,and N. Biswal.1977.Studies
on the intracellular replicating DNA of herpes simplexvirus type 1.Virology81:48-61.
15. Horii, Z.-I.,and A.J.Clark. 1973.Geneticanalysisof theRecF pathwaytogeneticrecombination inE. coliK12: isolation and characterization ofmutants.J. Mol. Biol. 80:327-344. 16. Leach,D. R. F., and F. W. Stahl. 1983. Viability of Aphages
carryingaperfectpalindromein the absence ofrecombination nucleases. Nature(London)305:448-451.
17. Lilley,D. M.J.1981.Invivo consequences ofplasmid topology.
Nature(London)292:380-382.
18. Luchnik, A. N., V. V. Bakayev, I. B. Zbarsky, and G.P. Georgiev.1982.Elastic torsional strain in DNA withinafraction ofSV40 mini-chromosomes: relationtotranscriptionallyactive chromatin. EMBOJ. 1:1353-1358.
19. Maniatis, T., E. F. Fritsch,andJ. Sambrook. 1982. Molecular cloning:alaboratorymanual. ColdSpringHarborLaboratory,
ColdSpring Harbor, N.Y.
20. Maxam,A.M.,and W. Gilbert.1980. Sequencingend-labelled DNAwithbasespecificchemicalcleavage. MethodsEnzymol.
65:499-560.
21. Mizuuchi, K., M. Mizuuchi, and M. Gellert. 1982. Cruciform
structuresinpalindromicDNAarefavoredbyDNA
supercoil-ing.J. Mol. Biol. 156:229-243.
22. Mocarski,E.S.,andB. Roizman.1982.Herpesvirus-dependent amplification and inversion of cell-associated viral
thymidine
kinase gene flankedbyviralAsequencesand linkedtoanorigin
of viral DNA replication. Proc. Natl. Acad. Sci. USA 79:5626-5630.23. Murchie, M.-J., and D. J. McGeoch. 1982. DNA sequence analysisofanimmediate-earlygeneregionoftheherpes
simplex
virus type1 genome.J.Gen. Virol.62:1-15.24. Myers,R. M.,andR.Tjian.1980.Constructionand
analysis
of simianvirus 40 originsdefectiveintumorantigenbindingand DNAreplication.Proc. Natl. Acad. Sci. USA77:6491-6495. 25. Nader,W.F.,T. D.Edlind,A.Huettermann,andH. W.Sauer.1985.CloningofPhysarumactin sequences inan exonuclease-deficient bacterial host. Proc. Natl. Acad. Sci. USA 82: 2698-2702.
26. Panayotatos,N., and R.D. Wells. 1981.Cruciformstructuresin supercoiledDNA.Nature(London)289:466-470.
27. Rao, R. N.,and S. G. Rogers. 1979. PlasmidpKC7: a vector
on November 10, 2019 by guest
http://jvi.asm.org/
containing ten restriction endonuclease sites suitable for cloning DNA segments. Gene. 7:79-82.
28. Razin, A., H. Cedar, andA.D. Riggs (ed.).1984. DNA methyla-tion:biochemistry and biological significance, p. 1-10. Springer-Verlag, Inc., New York.
29. Sadler, J. R., J. L. Betz, and M.Tecklenburg. 1978. Cloning of chemically synthesized lactose operators. II. EcoRI-linked op-erators.Gene 3:211-232.
30. Sanger, F., S. Nicklen,and A.R.Coulson. 1977.DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
31. Sinden, R. R., D. E. Pettijohn, and B. Francke. 1982. Organiza-tionof herpes simplex virus type 1 deoxyribonucleic acid during replication probed in living cells with 4,5',8-trimethylpsoralen. Biochemistry21:4484-d490.
32. Southern, E. 1979.Gelelectrophoresis of restriction fragments. Methods Enzymol.68:152-176.
33. Spaete, R. R., andN.Frenkel. 1982. The herpessimplex virus amplicon: anew eucrayoticdefective-virus cloning-amplifying vector.Cell 30:295-304.
34. Spaete, R.R.,andN. Frenkel. 1985. The herpessimplex virus amplicon: analyses of cis-acting replication functions. Proc. Natl.Acad. Sci. USA82:694-698.
35. Stow,N. D. 1982. Localization ofanorigin ofDNAreplication within theTRs/IRsrepeated region of the herpes simplex virus type1genome. EMBO J. 1:863-867.
36. Stow, N.D.1985.Mutagenesis ofaherpessimplex virus origin of DNAreplication and its effect on viral interference. J. Gen. Virol.66:31-42.
37. Stow, N. D., andE. C. McMonagle. 1983. Characterization of
theTRs/IRsoriginof DNA replication of herpes simplex virus type 1.Virology 130:427-438.
38. Vlazny, D. A., and N. Frenkel. 1981. Replication of herpes simplex virus DNA: localization of replication recognition sig-nals within defective virus genomes. Proc. Natl. Acad. Sci. USA 78:742-746.
39. Walboomers,J. M. M.,andJ.T.Schegget. 1976. A new method for the isolation of HSV-2DNA. Virology 74:256-258. 40. Watson,R.J.,andG.F.VandeWoude. 1982. DNAsequenceof
animmediate-early gene (IE mRNA-5) of herpes simplex virus type I.Nucleic Acids Res. 10:979-991.
41. Weller, S. K., K. J. Lee, D. J. Sabourin, and P. A. Schaffer. 1983. Geneticanalysis oftemperature-sensitive mutantswhich define the gene for the major herpes simplex virus type 1 DNA-binding protein. J. Virol.45:354-366.
42. Weller, S. K.,A.Spadaro, J.E.Schaffer,A. W.Murray,A. M. Maxam, and P.A. Schaffer. 1985. Cloning, sequencing, and functionalanalysis of oriL, a herpes simplex virus type1origin ofDNAsynthesis. Mol. Cell. Biol. 5:930-942.
43. Whitton,J. L.,andJ.B.Clements. 1984.Replication origins and asequence involved in coordinate induction of the immediate early genefamilyareconserved inanintergenic region of herpes simplex virus. Nucleic AcidsRes. 12:2061-2079.
44. Wyman,A.R.,L. B.Wolfe,and D.Botstein.1985. Propagation ofsome human DNA sequences in bacteriophage vectors re-quires mutant Escherichia coli hosts. Proc. Natl. Acad. Sci. USA 82:2880-2884.
45. Yanisch-Perron, C., J. Vieira, andJ. Messing. 1985. Improved M13 phage cloning vectors and host strains: nucleotide se-quences oftheM13mpl8andpUC19vectors.Gene 33:103-119. 58,