0022-538X/07/$08.00⫹0 doi:10.1128/JVI.01215-07
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Mutations in Envelope gp120 Can Impact Proteolytic Processing of the
gp160 Precursor and Thereby Affect Neutralization Sensitivity of
Human Immunodeficiency Virus Type 1 Pseudoviruses
䌤
Wendy M. Blay,
1Theresa Kasprzyk,
3Lynda Misher,
1Barbra A. Richardson,
4and Nancy L. Haigwood
1,2,4*
Viral Vaccines Program, Seattle Biomedical Research Institute, Seattle, Washington 98109,1and Departments of Pathobiology,2
Microbiology,3and Biostatistics,4University of Washington, Seattle, Washington 98195
Received 3 June 2007/Accepted 31 August 2007
The design of an efficient human immunodeficiency virus (HIV) immunogen able to generate broad neutralizing antibodies (NAbs) remains an elusive goal. As more data emerge, it is becoming apparent that one important aspect of such an immunogen will be the proper representation of the envelope protein (Env) as it exists on native virions. Important questions that are yet to be fully addressed include what factors dictate Env processing, how different Env forms are represented on the virion, and ultimately how these issues influence the development and efficacy of NAbs. Recent data have begun to illuminate the extent to which changes in gp41 can impact the overall structure and neutralizing sensitivity of Env. Here, we present evidence to suggest that minor mutations in gp120 can significantly impact Env processing. We analyzed the gp120 sequences of 20 env variants that evolved in multiple macaques over 8 months of infection with simian/human immunodeficiency virus 89.6P. Variant gp120 sequences were subcloned into gp160 expression plasmids with identical cleavage motifs and gp41 sequences. Cells cotransfected with these plasmids and ⌬env genomes were able to produce competent virus. The resulting pseudoviruses incorporated high levels of Env onto virions that exhibited a range of degrees of virion-associated Env cleavage (15 to 40%). Higher levels of cleavage correlated with increased infectivity and increased resistance to macaque plasma, HIV immunoglobulin, soluble CD4, and human monoclonal antibodies 4E10, 2F5, and b12. Based on these data, we discuss a model whereby changes in gp120 of 89.6P impact Env processing and thereby mediate escape from a range of neutralizing agents.
The envelope protein (Env) is the only viral protein on the surface of human immunodeficiency virus type 1 (HIV-1) and, as such, is the sole target of neutralizing antibodies (NAbs). The mature Env glycoprotein comprises a 120-kDa surface unit (gp120) noncovalently bound to a 41-kDa transmembrane unit (gp41). The HIV Env precursor is synthesized as a gp160 polyprotein, which is cotranslationally modified by the addition of high-mannose carbohydrates in the endoplasmic reticulum. Trimerization also occurs in the endoplasmic reticulum, after which the precursor is cleaved by furin or a related cellular protease, presumably in the trans-Golgi network (56). Furin cleaves gp160 primarily at the motif R-x-R/K-R (24) but can also cleave a secondary site, K-x-K-R-R, located 8 amino acids N-terminal to the first (32). The cleaved trimer undergoes further modifications of the N-linked carbohydrates through-out the Golgi network until the mature Env spike is trans-ported to the cell membrane for incorporation into the assem-bling virus. Several lines of evidence suggest that the Env spikes interact with the structural matrix (MA) protein via the cytoplasmic tail of gp41 (15, 17, 26, 57, 84). However, this interaction does not appear to be a requisite for Env incorpo-ration, as cytoplasmic-tail-truncated mutants can still be
incor-porated onto the virion (26, 27, 57). Likewise, both cleaved and uncleaved Env spikes can become part of the budding virion (35, 36, 38, 55, 58). Much of the envelope present on a virion may exist in a nonfunctional form, as it is often suggested that the high mutation rate of HIV results in a majority of defective virions. One study showed that defective virions can make up 50 to 90% of the total viral population, depending on the strain (8).
HIV interacts with the target cell by first binding to the CD4 receptor via a recessed pocket on gp120 (44). This triggers a large conformational change which creates the coreceptor binding site from the gp120 bridging sheet and variable loops 1, 2, and 3 (13, 44). Upon the binding of coreceptors such as CCR5 and CXCR4, another conformational change displaces gp120 to expose the fusion peptide of gp41, ultimately allowing for the fusion of the viral and cellular membranes (69). These dynamic processes mean that only cleaved Env is functional for fusion (51). Uncleaved Env is able to bind CD4 and may therefore play a role in providing a stable form of Env capable of tethering the virus to the target cell (58, 74).
Data obtained over the last several years have continued to unveil the extraordinary plasticity of Env and the unexpected complexity of both the tertiary (13, 43, 44, 80) and quaternary (30) structures. Recently, it has been shown by cryo-electron microscopy that HIV has on average 14 (⫾7) Env spikes per virion (85). It is not possible to know from this analysis what proportion of the Env spikes exist in a functional cleaved form, though recent evidence suggests that many alternate Env forms * Corresponding author. Mailing address: Seattle Biomedical
Re-search Institute, 307 Westlake Ave. N, Suite 500, Seattle, WA 98019. Phone: (206) 256-7338. Fax: (206) 256-7229. E-mail: Nancy.Haigwood @sbri.org.
䌤Published ahead of print on 12 September 2007.
13037
on November 8, 2019 by guest
http://jvi.asm.org/
are likely to be present (55). These include gp41 spikes and monomeric gp120 and gp41 (55), as well as cleaved and un-cleaved oligomeric Env (19, 37, 48, 82).
Mounting evidence suggests that uncleaved Env and fully processed Env are antigenically different (5, 35, 36, 61). A key distinction between antibodies that are able to neutralize the virus and those that are not is the ability to bind functional oligomeric Env. While both neutralizing and nonneutralizing antibodies can bind monomeric and possibly uncleaved Env, only NAbs are able to bind functional trimers (55, 61). Like-wise, uncleaved gp160 may be preferentially recognized by nonneutralizing antibodies (58). This model helps explain why CD4 binding site (CD4bs)-directed monoclonal antibodies (MAbs) can exhibit dramatic differences in neutralization; the MAbs b6 and b12 compete for virion capture, but only b12 is able to bind functional oligomeric Env. Likewise, b12 is po-tently neutralizing while b6 is not (61). Similarly, there is a growing body of evidence to suggest that uncleaved Env and fully processed Env are immunogenically different. It has been suggested that one role of uncleaved gp160 may be to act as a decoy antigen and elicit antibodies that are not neutralizing against functional, fully cleaved Env (6, 61). Antibodies that are elicited against cleavage-incompetent gp140 show only modest neutralization of functional full-length Env trimerized on a virion (21, 67). Although oligomeric gp140 immunogens elicit better cellular and humoral immune responses than mo-nomeric gp120 (2, 3, 40, 83), gp160 presented on virus-like particles continues to generate the best immune response (49). The factors responsible for differences in Env processing remain unclear. The cleavage of gp160 by furin is inherently inefficient (32, 76), potentially due to a less-than-ideal confor-mation of the protease recognition site on gp160. Although in vitro modification of the recognition site can enhance cleavage efficiency, these mutations are not found to occur naturally in circulating variants of HIV (6). Incomplete Env cleavage does not appear to be purely an artifact of the pseudotype method, as it has also been observed previously in replication-compe-tent virions (19, 20, 48). Higher levels of furin can result in enhanced processing, so producer cell effects likely exist (6, 39). However, even within the same cell type, viruses with the same cleavage motifs still demonstrate a range of cleavage efficiencies (6, 58), implying that other factors such as protein folding must play a role. It is well established that one of the functions of carbohydrates on glycoproteins is to mediate pro-tein folding (see reference 34 for a review). It therefore stands to reason that differences in glycosylation may result in differ-ences in folding. Since the enzymatic activity of furin relies at least in part on the conformation of the protein surrounding the cleavage site (22), it follows that changes in folding may impact cleavage efficiency. In other viral systems, such as Ebola and influenza viruses, different levels of cleavage have been associated with increased virulence (23, 28, 33). Furthermore, higher levels of cleavage would be expected to result in a higher proportion of functional Env spikes and may therefore require greater amounts of NAbs, thereby operating as an additional escape mechanism. In this study, we provide evi-dence from an animal model of infection with simian/human immunodeficiency virus 89.6P (SHIV89.6P) that relative
modi-fications in gp120 distant from the cleavage site can impact Env processing. We show additionally that the level of processed
Env can impact infectivity and sensitivity to neutralization by a range of NAbs.
MATERIALS AND METHODS
Viral stock and selection of variants.SHIV89.6Pwas passaged twice through
samples ofMacaca nemestrinaperipheral blood mononuclear cells depleted of
CD8⫹T cells, the in vivo titers were determined, and the virus was used to
challengeM.nemestrinamacaques intrarectally with 50 50% macaque infectious
doses (18). The sequence of the proviral Env from the peripheral blood mono-nuclear cells used to grow the virus was identical to the published sequence of KB9 (accession no. U89134); a form of gp160 from a clone representing the consensus was chosen as the wild type (WT) for subsequent experiments. Ap-proximately 10 gp120 proviral clones were previously generated from each of nine macaques at 32 weeks postinfection (wpi) and demonstrated to have several convergent changes in potential N-linked glycosylation (PNG) sites (7). Clones were chosen to represent all combinations of PNG site changes seen within each macaque. The resulting 24 total gp120 clones were subcloned into a gp160 expression plasmid (see below) and tested for functionality with a cell-cell fusion assay. Ultimately, 20 Env variants were able to express Env at a reliable level for detection and to mediate fusion. These clones were used to create pseudoviruses. The sequencing of the variable 1 (V1)-to-constant 5 (C5) region of these 20 clones was described previously in reference 7, and the sequences have been assigned GenBank accession numbers DQ105094 to DQ105195. Sequencing of the C1 region was added for the present analysis.
Sequence analysis.The new sequence from C1 was added to the previously published V1-to-C5 sequence (7) using Invitrogen Vector NTI Advance 10. Sequences were aligned with ClustalX (77) and manually edited using BioEdit 5.0.9 (31). The Los Alamos National Laboratories SeqPublish program http: //www.hiv.lanl.gov/content/hiv-db/SeqPublish/seqpublish.html was used to gener-ate the final output. The diversity of the variant population was calculgener-ated in MEGA 3.1 (42) using the Kimura two-parameter model with pairwise deletions (transition-to-transversion ratio of 2). For an analysis of overall divergence, the variant population was compared to the WT sequence of 89.6P and KB9 (ac-cession no. U89134) by using MEGA 3.1 and the parameters described above. The range of nucleotide distances between any two variants was calculated using MEGA 3.1 pairwise analysis and the Kimura two-parameter model with pairwise deletions.
Subcloning of gp120 gene into expression plasmid.The gp120 clones were digested out of the 2.1 TOPO-TA vector with MfeI and KpnI (New England Biolabs, Beverly, MA), and the gp120 fragment was ligated into pEMC* with T4 DNA ligase (New England Biolabs, Beverly, MA). MAX Efficiency DH10B
competent cells (50l) were transformed with 5.5 ng of total plasmid DNA per
the instructions of the cell manufacturer (Invitrogen, Carlsbad, CA) and grown at 30°C for 24 h. The plasmid was purified with a QIAGEN miniprep kit and sequenced for verification of the insert.
Pseudovirus construction.The viral backbone plasmid Q23⌬Env was kindly provided by Julie Overbaugh (47). 293T cells were transfected with Fugene 6 per the instructions of the manufacturer (Roche, Indianapolis, IN). Cells were plated
to 50% confluence into a T75 culture flask; 4g of total DNA at a 20:1
backbone/Env plasmid ratio was prepared in a mixture of 12l of Fugene and
188l of Dulbecco’s modified Eagle’s medium (DMEM). Virus was harvested
48 h later, spun at 2,000 rpm for 10 min, and stored at⫺70°C until use.
Determination of pseudovirus titers.Pseudovirus titers on Tzm-bl cells were determined in quadruplicate by using a 96-well-plate format. Fivefold serial dilutions of virus were added to complete medium (DMEM, 10% fetal calf
serum, 1%L-glutamine, 1% penicillin-streptomycin) in the presence of 7.5g of
DEAE-dextran/ml, and the plates were incubated at 37°C for 1 h. Each well
received 100l of Tzm-bl cells resuspended in medium at 105
cells/ml. Forty-eight hours later, cells were lysed directly on the neutralization plates for 2 min
by using 100l of Bright-Glo luciferase assay substrate (Promega, Madison, WI)
and immediately analyzed for luciferase activity with a luminometer. For neu-tralization assays, the amount of 200 50% tissue culture infective doses was calculated according to the method of Reed and Muench (63). For comparison of relative degrees of infectivity among variants, the luciferase relative light units (RLU) normalized to the p24 level are reported. Calculations were carried out as follows: pseudovirus titers on Tzm-bl cells were determined for four or eight replicates. The RLU for a viral dilution of 1:125, the dilution at which all pseudoviruses were beyond the point of cytopathicity and within the linear range, were obtained. The background value was subtracted for each assay, and the value for the input virus was normalized relative to the picograms per microliter of p24.
13038 BLAY ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
Neutralization assay.Plasma, soluble CD4 (sCD4), MAbs, and HIV immu-noglobulin (HIVIG) were tested for the ability to neutralize each pseudovirus. Plasma samples were heat inactivated at 56°C for 1 h and spun at 14,000 rpm for 5 min to remove coagulants; twofold serial dilutions were tested. Stably trans-formed Chinese hamster ovary (CHO) cells were used to produce sCD4, tested
at a final concentration of 8.5g/ml. The following reagents were obtained
through the NIH AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH: HIV-1 MAb IgG1b12 was obtained from Dennis Burton and Carlos Barbas (10, 11), and HIV-1 gp120 MAbs 2G12, 4E10, and 2F5 were obtained from Hermann Katinger and purchased from ImmunoAG (Vienna, Austria) (9, 62, 75). HIVIG (lot no. IHV-50-11) was a kind donation from David Montefiori. The Tzm-bl neutralization assay was performed in 96-well plates as described previously (79). Two hundred 50% tissue culture infective doses of virus were added to serial dilutions of either plasma, sCD4, MAb, or HIVIG in
the presence of 7.5g of DEAE-dextran/ml in a total volume of 150l of
medium (DMEM, 10% fetal calf serum, 1%L-glutamine, 1%
penicillin-strepto-mycin), and the plates were incubated for 1 h at 37°C. Each well received 100l
of Tzm-bl cells resuspended in medium at 105
cells/ml. Forty-eight hours later,
cells were lysed directly on the neutralization plates for 2 min by using 100l of
Bright-Glo luciferase assay substrate (Promega, Madison, WI) and immediately analyzed for luciferase activity with a luminometer. All samples were tested in triplicate, and a random sampling of plasma-virus combinations was replicated in multiple independent assays to verify reproducibility within 2.5-fold. All values were
calculated with respect to virus only [(value for virus⫺value for cells only)⫺
(value for plasma⫺value for cells only)]/(value for virus⫺value for cells only)].
For assays with plasma, the reciprocal of the mean (or geometric mean) dilution necessary to achieve 50% neutralization is reported. Note that the sampling times for the late-time-point plasma samples ranged from 33 to 53 wpi. The lowest dilution of prebleed plasma tested (1:100) never achieved 50% neutral-ization. For sCD4, MAbs, and HIVIG, the concentration of antibody required to
obtain 50% neutralization (the 50% inhibitory concentration [IC50]) is reported.
Quantitative Western blot analysis.Pseudoviruses were centrifuged at 4°C for 10 min at 2,000 rpm to remove cellular debris and then pelleted to a
50-fold-higher concentration through a 20% glycerol cushion for 2 h at 50,000⫻gin an
SW40 Ti rotor. Pellets were lysed in 1% Triton–phosphate-buffered saline (PBS), subjected to a vortex, and incubated at 37°C for 1 h. Protein agarose gel elec-trophoresis was performed on 4 to 12% Tris-bis minigels by using the NuPAGE system (Invitrogen, Carlsbad, CA) under reducing conditions. Recombinant SF162 gp120 was kindly provided by Novartis (Emeryville, CA) as a size standard
and positive control;⌬Env pseudovirus was used as a negative control. Gels were
blotted onto a nitrocellulose membrane (Invitrogen, Carlsbad, CA), blocked with Odyssey blocking buffer (LI-COR Biosciences, Lincoln, NE), and probed simul-taneously for Env and p24. Env was detected with 1:8,000 anti-gp120-anti-gp140 polyclonal rabbit plasma. p24 was detected with 1:1,000 HIV-1 p24 Gag MAb (no. 24-3) obtained through the AIDS Research and Reference Reagent Pro-gram, Division of AIDS, NIAID, NIH, from Michael H. Malim (25, 72, 73). Primary antibodies were prepared in a mixture of 1:1 Odyssey blocking
buffer–1⫻PBS and 0.2% Tween. Blots were washed with 0.1% Triton X-100–1⫻
PBS. Secondary antibodies were 1:15,000 IRDye 700DX goat anti-rabbit IgG and 1:15,000 IRDye 800DX goat anti-mouse IgG (LI-COR Biosciences, Lincoln,
NE) prepared in a mixture of 1:1 Odyssey blocker–1⫻PBS, 0.2% Tween, and
0.02% sodium dodecyl sulfate. Membranes were scanned with the Odyssey in-frared imaging system (LI-COR, Lincoln, NE), and integrated intensities (i.i.) were calculated with a user-defined background. The total amount of Env was calculated from the sum of the i.i. of gp120 and the i.i. of gp160; values were normalized relative to the level of p24 in the same blot. The percent cleavage was
determined for each variant by the following equation: (i.i. of gp120)⫼(i.i. of
gp120⫹i.i. of gp160). Each pseudovirus was analyzed in two to six independent
assays.
Gag p24 enzyme-linked immunosorbent assay.Gag p24 was quantified using an HIV-1 p24 antigen capture assay kit (AIDS vaccine program, National Cancer Institute-Frederick Cancer Research and Development Center, Frederick, MD). Pseudovirus lysates were prepared as described above; twofold serial dilutions
were tested in duplicate. Twofold serial dilutions of 10 ng of HIVMNlysate/ml
were used as a positive control. Plates were coated with HIV-1 p24-specific MAb and blocked with 0.5% casein. Gag p24 was detected with 1:400 rabbit anti-HIV-1 p24 sera, followed by 1:100 goat anti-rabbit IgG (heavy-plus-light-chain) peroxidase-labeled antibody (KPL), and the reaction was developed with
3,3⬘,5,5⬘-tetramethylbenzidine substrate. The reaction was stopped with 1 N
H2SO4, and the results were read on a luminometer atA450toA650by using
SoftMax Pro (Molecular Devices, Sunnyvale, CA).
Statistical analyses.To test for associations between variables, Spearman’s correlation coefficient was used. In MAb analyses in which the level of
neutral-ization was outside the range tested, the values were reported to be one digit beyond the range in order to perform statistical calculations (for example, greater than 30 was calculated as 31). To compare measures between individual pseudovirus variants and the WT, each variant was tested multiple times and a mean value for the variant was obtained. Then the ratio of each variant value to the WT value was calculated, and Wilcoxon signed-rank tests were performed to test if these variant ratios were significantly different from 1, as described in reference 66. The infectivity levels of different variants were analyzed with an
independentttest. Unless otherwise noted, all statistical analyses performed
were two-tailed with an alpha level of 0.05.
RESULTS
Subcloning of gp120 variant sequences into gp160 expres-sion plasmids with identical cleavage sites.Differences in gp41 are known to impart several structural effects on the overall Env protein (1, 16, 26, 45, 57). In order to determine the potential role that gp120 has in Env processing, the mainte-nance of a constant gp41 region was critical to this analysis. Previously, we reported on a cohort of macaques that were challenged with SHIV89.6Pand monitored for approximately 1
year postinfection (7, 18). Twenty-four representative gp120 variants that developed over 32 weeks of infection in nine of these macaques were chosen for this analysis. While providing a collection of different gp120 sequences evolved in vivo during infection, these variants all arose from the same starting virus and, therefore, also provided the opportunity to analyze the phenotypes of these gp120 variants fused to the invariant gp41 from the inoculum. This approach was chosen to minimize possible complications that may arise from mismatched gp120 and gp41 sequences.
The WT 89.6P full-length gp160 sequence was cloned into the pEMC* expression vector. The gp120 sequence corre-sponding to the region from C1 to C5 was digested out of the expression plasmid and replaced with the variant gp120 se-quences. The primary cleavage site between gp120 and gp41 is REKR at positions 508 to 511 (32). A secondary cleavage site exists upstream, at KAKRR (residues 500 to 504); however, the 89.6P WT sequence for the upstream cleavage site replaces the first K with an R (resulting in RAKRR). Our subcloning strategy preserved both the primary and secondary cleavage sites, holding them constant across all variant pseudoviruses (Fig. 1A).
293T cells were transfected with each expression plasmid and tested for Env expression and the ability to mediate fusion. One variant failed to express Env, and two variants expressed Env but did not mediate fusion (data not shown). The remain-ing 21 variants successfully complemented the Q23⌬Env ge-nome to create infectious pseudovirus. The level of envelope expression of one variant was too low to be reliably measured, and this variant was excluded from further study. Ultimately, we examined 20 variant pseudoviruses and a WT 89.6P Env that was created from the consensus Env sequence from the inoculum. This collection of variants represented a 1.3% di-vergence from the original inoculum and 1.9% diversity. Al-though several PNG sites were well conserved among all vari-ants in this study, a number of PNG sites were added, removed, or shifted in relation to those in the WT sequence. PNG site sequons are shaded in gray in Fig. 1B, showing that the col-lection of variants represented a range of PNG site patterns. Several point mutations occurred throughout the variants and included several changes that would be predicted to have a
on November 8, 2019 by guest
http://jvi.asm.org/
13040 BLAY ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
possible impact on the protein structure, such as changes in prolines and charged residues. Many of the observed sequence changes from WT 89.6P occurred in multiple variants; how-ever, none of the variants possessed the exact same collection of changes (Fig. 1B).
Pseudovirus envelopes exhibit a range of degrees of proteo-lytic processing.Each variant Env was used to create 5 to 10 ml of pseudovirus, which was then purified to remove cellular debris and pelleted through a 20% glycerol cushion. The pel-leted pseudoviruses were lysed and analyzed by quantitative Western blotting under reducing conditions in order to deter-mine the relative amounts of virion-associated gp160 and gp120. The amount of gp120 relative to the total amount of Env (gp160 ⫹gp120) is reported as the percent cleaved. A recombinant gp120 protein was used in each assay as a size
[image:5.594.48.543.71.449.2]standard for the identification of gp120 (Fig. 2A, lane 1, gp120). Pseudovirus was created in the absence of Env to serve as a negative control and, as expected, was not reactive with anti-Env antiserum (Fig. 2A, lane 2, no Env). Each Env variant was analyzed for cleavage multiple times, and there was good reproducibility of results (data not shown). In a subset of blots, the membrane was probed with the MAb 2F5 in order to verify the location of the gp160 band (data not shown). An analysis of virion-associated Env revealed a surprising range of cleavage efficiencies despite identical cleavage motifs. Approximately 15% of total Env on the WT pseudovirus existed in a cleaved form, while the majority of variants possessed a higher per-centage of cleaved Env (median, 23%; Wilcoxon signed-rank test;P ⫽0.003.) Only 1 variant showed a significantly lower level of cleavage than the WT, 10 variants showed significantly FIG. 1. Variant gp120 sequences were subcloned into a gp160 expression plasmid. (A) The 89.6P subcloning strategy is shown as a linear schematic, with the primary cleavage site between gp120 and gp41 shown as a heavy black line. The 89.6P variant gp120 sequences were subcloned into an expression vector containing the 89.6P-KB9 sequence by using a KpnI restriction site in the region corresponding to gp120 C1 and an MfeI restriction site in the region corresponding to gp120 C5 (denoted by arrowheads). The insertion of variant sequences occurred N-terminal of both the primary (REKR) and secondary (RAKRR) cleavage sites, shown in bold. (B) The sequence of the gp120 region that was spliced into the gp160 expression plasmid is shown for each variant. Sequences are aligned to the full-length WT 89.6P, and the N- and C-terminal splice sites are shown as arrowheads and dashed lines. Identical residues are represented by dashes, and insertions are shown as dots. PNG sites are highlighted with gray shading and labeled according to the HXB2 reference strain. The variable regions are labeled at the top of the alignment. Both the primary and secondary cleavage sites are shown in bold.
on November 8, 2019 by guest
http://jvi.asm.org/
higher levels of cleavage than the WT, and 9 variants were not significantly different from the WT (Fig. 2A).
In order to examine if the envelope detected in our system represented a substantial amount of contamination from mem-brane-associated Env in microvesicles independent of virions, we performed a control experiment using envelope transfec-tion only. 293T cells were transfected with the WT 89.6P gp160 expression plasmid in the absence of the genomic backbone plasmid. Supernatants were harvested in the same manner as all pseudoviruses in the study: 48 h posttransfection, the su-pernatants were centrifuged to remove cellular debris, pelleted through a 20% glycerol cushion at 50,000⫻gfor 2 h, lysed with
1% Triton, and analyzed by Western blotting for the presence of Env. Neither gp120 nor gp160 was detected under these conditions (Fig. 2A, lane 3, Env only).
The high levels of exogenous protein expression that exist with a transfection system can theoretically distress the pro-ducer cell and lead to the incomplete processing of the desired protein. This effect may lead to an artifact wherein higher levels of envelope expression result in less efficiently processed envelope. To examine whether this consequence explained the observed differences in envelope cleavage, we used a quanti-tative Western blot to determine the levels of expression of total Env (gp120⫹gp160) normalized to the p24 level. Our FIG. 2. Pseudovirus envelopes exhibit a range of degrees of proteolytic processing, despite identical cleavage motifs. (A) A compilation of representative gels for each variant is shown. Recombinant gp120 is shown as a positive control, and the⌬Env pseudovirus (no Env) is shown as a negative control. Supernatant from an envelope-only transfection is shown as a control for membrane-associated envelope (Env only). The percentage of gp120 relative to total Env is reported as percent cleaved. Variants with cleavage levels significantly different from that of the WT in an independentttest with a two-tailedPvalue of⬍0.05 are denoted byⴱ. Variants are grouped according to the macaque from which they were cloned. NA, not applicable. (B) The normalized total Env amount and the relative amounts of gp120 and gp160 for each variant are shown. The percent cleavage is shown on the right for convenience.
13042 BLAY ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
pseudovirus preparations were comprised of a clade A back-bone expressing Gag with decreased reactivity to the clade B-derived antibody used to detect Gag in our assays. There-fore, the amount of Gag detected in our system was lower than that reported in other studies (48). The total Env/p24 ratio for WT 89.6P was set at 100; the values for all other variants were calculated relative to the WT value (Fig. 2B). Overall, variants had less total Env than the WT (median ratio of variant Env to WT Env, 0.36; Wilcoxon signed-rank test;P⫽0.003). Impor-tantly, there was not a significant correlation between Env expression and the percentage of cleaved Env (Spearman’s correlation;P⫽0.4).
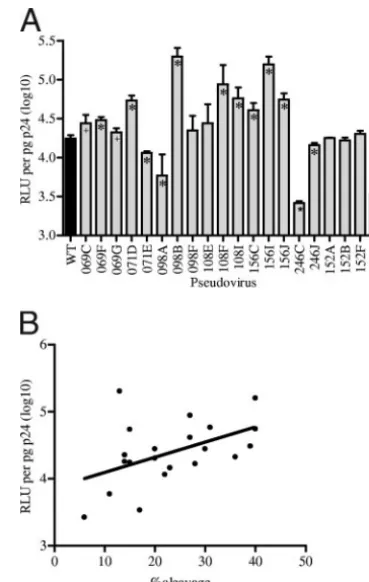
Infectivity of pseudoviruses correlates with percentage of envelope cleavage. With the gp120 variants demonstrating a sevenfold range in the levels of virion-associated functional Env, we hypothesized that there would be a concomitant effect on viral infectivity. To determine the relative infectivity of the variants, average RLU were normalized to p24 levels for each pseudovirus. RLU were obtained using a viral dilution of 1:125, the dilution at which pseudoviruses were all beyond the point of cytopathicity and within the linear range. The deter-mination of viral titers and the p24 enzyme-linked immunosor-bent assays were performed with the same preparation of a given pseudovirus in order to remove possible experimental variation introduced by multiple preparations. The levels of infectivity of 15 of the 20 variants were significantly different from that of the WT (two-tailed independent t test); only variants 152A, 152B, 152F, 108E, and 098F did not differ from the WT (P⬎0.05). Of the 15 variants that were significantly different, 11 showed increased infectivity and 4 showed de-creased infectivity (Fig. 3A). As predicted, the level of infec-tivity showed a positive correlation with the percentage of envelope cleavage (Spearman’s correlation;r⫽0.45;P⫽0.04) (Fig. 3B).
Correlation between percent cleavage and sensitivity to neu-tralization by macaque plasma.Since our panel of pseudoviral variants was derived from the evolution of the same starting virus in multiple macaques, we were interested in testing the ability of the plasma from individual macaques to neutralize autologous variants as well as variants obtained from other macaques. We therefore performed a matrix of neutralization comparing the entire panel of 20 variants against the anti-HIV envelope NAbs that were generated in vivo in different ani-mals. We tested the neutralization potential of the latest plasma sample available from each macaque, collected be-tween 33 and 53 wpi as noted in Fig. 4.
Sixteen of the 20 envelopes tested represented escape vari-ants that were more resistant than WT 89.6P to autologous NAbs. For example, plasma from the late-time-point (39-wpi) sample from macaque 156 was able to achieve 50% neutral-ization of WT 89.6P at a dilution of 1:5,244. All three variants derived from macaque 156 at 32 wpi (156C, 156I, and 156J) were more difficult to neutralize than the WT. In contrast to the dilution required for the WT, a plasma dilution of only 1:779 was required to achieve 50% neutralization of 156J, while 156C and 156I were not neutralized at the lowest tested dilution of 1:100 (Fig. 4, first column, rows 156C, 156I, and 156J).
Of the 20 Envs tested, 4 were not escape variants. The 098A Env provides an interesting example of a variant that was
substantially more sensitive than the WT to autologous plasma (50% neutralization titers against the pseudoviruses were 18,000 and 2,258, respectively). However, 098A was not neu-tralized by prebleed plasma and is therefore not inherently sensitive to nonspecific neutralization (data not shown). Ma-caque 152 had undetectable levels of NAbs against the WT 89.6P virus (Fig. 4, top row). The three variants derived from macaque 152 (152A, 152B, and 152F) can therefore not be described as true autologous escape variants within the limits of detection of this study. Nevertheless, we were interested in testing the sensitivity of these variants to plasma from ma-caques with a more robust NAb response. The final exception was macaque 168. The only variant derived from macaque 168 was the previously described Env variant that was excluded from analyses due to low Env expression. Although there were no autologous variants tested against plasma from macaque 168 in this matrix, we did complete an analysis of 168 plasma against heterologous variants.
[image:7.594.326.511.68.359.2]Fifteen of the 16 variants that mediated escape from autol-ogous NAbs also mediated escape from NAbs found in heter-ologous macaques in the cohort (variant 107C is the excep-tion). One hundred sixty combinations of late-time-point plasma samples and heterologous pseudoviruses were tested FIG. 3. Infectivity of pseudoviruses. (A) The infectivity of each pseudovirus is expressed as the average number of RLU per picogram of p24; error bars denote standard deviations. The WT pseudovirus is shown in black. Variants significantly different from the WT in an independentttest with a two-tailedPvalue of⬍0.001 are denoted by ⴱ, and those with aPvalue of⬍0.05 are denoted by⫹. (B) Two-sided Spearman’s correlation analysis showed a positive correlation between the percentage of envelope cleavage and infectivity (RLU per pico-gram of p24). Each pseudovirus variant is represented by an individual point.
on November 8, 2019 by guest
http://jvi.asm.org/
(Fig. 4). The majority (74%) of these pairings were unable to achieve 50% neutralization at the lowest dilution of plasma tested (1:100). When heterologous plasma samples were able to neutralize a variant, the level of neutralization achieved was typically lower than the level achieved against the WT. For example, plasma from macaque 156 was able to neutralize 50% of the 069F variant at a dilution of 1:693, while the same macaque 156 plasma neutralized 50% of the WT 89.6P at a dilution of 1:5,244. Overall, the relative sensitivity of a variant to heterologous plasma was generally consistent with the sen-sitivity of the variant to autologous plasma. In some cases, such as that of variant 069G, the pseudovirus was resistant to neu-tralization by all heterologous plasma samples tested. In con-trast, pseudovirus 098A, which showed enhanced sensitivity to autologous NAbs, also displayed enhanced sensitivity to het-erologous NAbs. Intriguingly, the ability of plasma from each macaque to neutralize the pseudoviral variants correlated in-versely with the percentage of cleavage demonstrated in the variants. The one exception was macaque 069, which did not show this correlation. Spearman coefficients and two-tailedP val-ues for the correlation between percent cleavage and neutraliza-tion by individual macaque plasmas are reported in Fig. 4.
Correlation between percent cleavage and sensitivity to neu-tralization by MAbs and HIVIG.In order to investigate if the observed correlation between Env cleavage and polyclonal
ma-caque plasma extended to a more heterologous polyclonal antibody source, we tested the panel of variants against pooled heterologous HIVIG from HIV-positive donors. The WT 89.6P pseudovirus required 850 g of HIVIG/ml for the achievement of 50% neutralization (Fig. 5A). There was a weak trend toward a correlation between greater resistance to HIVIG and a higher level of Env cleavage, though unlike that for autologous plasma, this correlation did not reach signifi-cance (two-tailed Spearman’s correlation;r⫽0.36;P⫽0.105). Not surprisingly, neutralization by MAb 2G12 was the ex-ception to the observed correlations with Env cleavage. The 2G12 epitope comprises the glycans N295, N332, and N392 (71). These glycans were present on WT 89.6P, resulting in an IC50of 2G12 of 2g/ml. Residues N386 and N448 participate
peripherally in the 2G12 epitope. N386 is not present on WT 89.6P but was added onto the majority of variants (Fig. 1B), resulting in a predicted increase in sensitivity to 2G12. The variants with glycan N386 added were typically the variants with higher levels of cleavage and, likewise, were more sensi-tive to neutralization by 2G12. This trend was seen as an inverse correlation between Env cleavage and sensitivity to 2G12 (two-tailed Spearman’s correlation; r ⫽ ⫺0.52; P ⫽ 0.015) (Fig. 5A).
[image:8.594.136.451.69.361.2]As a surrogate for the accessibility of the CD4bs, variants were tested for sensitivity to neutralization by sCD4 and FIG. 4. Correlation between percent cleavage and sensitivity to neutralization by macaque plasma. Each row represents a pseudovirus, with the WT in the top row. The percentage of envelope cleavage (%clv) associated with each pseudovirus is listed for convenience, and variants are ordered by increasing levels of cleavage. Columns represent late-time-point plasma samples from each of nine macaques; the week in which the plasma sample was obtained is shown in parentheses. The mean inverse dilution of plasma required to achieve 50% neutralization is reported for each macaque. Boxes are shaded with a grayscale gradient such that a lack of neutralization is shown in white and high sensitivity to neutralization is shown in dark gray. The percent cleavage was tested for a correlation with sensitivity to neutralization by each macaque’s plasma, and two-sided Spearman’s correlation coefficients (r) andPvalues are reported.
13044 BLAY ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
IgG1b12. Fifty percent neutralization of WT 89.6P was ob-tained at 30g of IgG1b12/ml. The IC50s for the majority (13
of 20) of the variants were⬎30g of IgG1b12/ml, the highest concentration tested in this study. Seven variants demonstrated a substantial increase in sensitivity, with variant 246C needing less than 0.12g of IgG1b12/ml for 50% neutralization (Fig. 5A). WT 89.6P was resistant to neutralization by 8.5 g of sCD4/ml, the highest concentration tested in this study. While half of the variants remained resistant to this concentration of sCD4, IC50s for 10 of the variants ranged from 0.375 to 8.4
g/ml. (Fig. 5A). As seen with autologous macaque plasma and in the trend with heterologous HIVIG, the ability of both IgG1b12 and sCD4 to neutralize the variant pseudoviruses correlated directly with the percentage of Env cleavage (two-tailed Spearman’s correlation;r⫽0.46 andP⫽0.036 andr⫽ 0.51 andP⫽0.017, respectively).
Because levels of cleavage differed among the variants, we were particularly interested in the neutralization profiles of the pseudovirions with MAbs directed to the gp41 membrane-proximal regions (MPRs). In this study, as previously noted, the gp120 variant sequences were all subcloned into an expres-sion plasmid that contained the WT 89.6P Env gp41 sequence. Despite having identical MPRs, the reconstructed variants demonstrated a range of sensitivities to gp41-directed MAbs 4E10 and 2F5. For each variant, the IC50s of MAbs 4E10 and
2F5 are reported in Fig. 5A. The neutralization of WT 89.6P required 10g of the MAb 4E10/ml. Although the majority of variants were also neutralized by 4E10, they required signifi-cantly higher concentrations of the MAb than the WT, dem-onstrating that changes in gp120 mediated escape from neu-tralization by the gp41-directed MAb. We once again observed a correlation between percent cleavage of the variant Env and the amounts of MAbs 4E10 and 2F5 necessary to achieve 50% neutralization (two-tailed Spearman’s correlation;r⫽0.75 and P ⫽ 0.0001 and r ⫽ 0.61 and P⫽ 0.003, respectively). The direct correlation between the percentages of cleavage of the panel of variants and the concentrations of MAbs 4E10 and 2F5 is plotted in Fig. 5B.
DISCUSSION
In this study, we have focused on the characteristics of HIV Env variants that arose in different macaques infected with SHIV89.6P over a time frame of approximately 1 year. We
[image:9.594.119.467.69.346.2]showed previously that these variants arose during the devel-opment of NAbs directed to the inoculum (7). Surprisingly, these gp120 variants demonstrated marked differences in the efficiency of Env cleavage, despite possessing identical cleav-age sites and gp41 domains. A higher proportion of properly cleaved Env should result in a higher probability that any three FIG. 5. Correlation between percent cleavage and sensitivity to neutralization by sCD4, MAbs, and HIVIG. (A) Each row represents a pseudovirus, with the WT in the top row. The percentage of envelope cleavage (%clv) associated with each pseudovirus is listed for convenience, and variants are ordered by increasing levels of cleavage. Each column represents a neutralizing agent. The concentration (micrograms per milliliter) of antibody required to achieve 50% neutralization is reported. Boxes are shaded with a grayscale gradient such that a lack of neutralization is shown in white and high sensitivity to neutralization is shown in dark gray. The percent cleavage was tested for a correlation with sensitivity to neutralization by HIVIG, sCD4, and each MAb. Two-sided Spearman’s correlation coefficients (r) andPvalues are reported. (B) Scatter plots depict the direct correlation between percentages of cleavage and the concentrations of MAbs 4E10 and 2F5 required for 50% neutralization (neut). Spearman’s correlation coefficients (r) and two-tailedPvalues are reported.
on November 8, 2019 by guest
http://jvi.asm.org/
monomers will combine as a functional spike. Virions with more functional spikes should result in a more infectious viral pool and would be predicted to require a higher level of anti-body to achieve neutralization, as there are data to suggest that each functional trimer must be occupied to neutralize HIV-1 (81). These predictions were supported by the data presented in this study.
Due to the inherently artificial nature of the pseudoviral approach, it is prudent to speculate if differences in Env cleav-age are an artifact of the system. Though this possibility has not been formally ruled out, we find it improbable, as others have reported that improperly processed Env is frequently incorpo-rated into a virion and can exist in a number of nonfunctional forms (20, 35, 36, 38, 48, 55, 58). It is also possible that the envelope we detected in our analysis of variant pseudovirus was largely membrane-associated envelope independent of ac-tual virions. It has been suggested previously that envelope can be nonspecifically targeted to multivesicular bodies (MVBs) and form virus-like structures independent of Gag (4, 29). In a pseudoviral system in which large amounts of viral proteins are being artificially translated by transfected cells, it is possible that large amounts of envelope are being sequestered in MVBs. If this was the case, we would expect this effect to be independent of Gag, and therefore, transfection with Env only should also result in the formation of Env-embedded MVBs. Although we cannot rule out this possibility definitively, the fact that we were unable to detect the presence of any Env in the absence of the genomic backbone suggests that MVB con-tamination did not play a significant role in the outcome of our experiments.
In this study, we looked at virion-associated Env and com-pared cleaved, intact Env that had the potential to be func-tional to uncleaved Env that was unable to mediate fusion. In addition to these two forms of Env, it is expected that there will also be gp41 spikes that remain virion associated from shed gp120. Further work is under way to examine questions regard-ing Env stability and gp41 spikes, as well as any differences between efficiencies and/or levels of stability of oligomeriza-tion. For this study, we chose to focus on the cleaved (and intact) versus uncleaved Env but recognize that there are likely other forms of Env that may also play an interesting role in this overall issue.
HIV virions are produced even when the level of Env cleav-age is exceptionally low. The virions maintain the ability to establish infection, but at a significantly lower rate and de-creased level (37, 65). Since only cleaved gp160 is fusion com-petent (51) and gp160 cleavage is naturally an inefficient pro-cess (6, 32, 76), it is reasonable to expect that Env changes that improve cleavage will also enhance HIV infectivity. In this study, we documented that escape variants generated from multiple macaques possessed significantly higher amounts of virion-associated cleaved Env than the WT and, importantly, that these increases in cleavage correlated with increased in-fectivity. As suggested by these data, Env processing may be an integral component of the virulence of different HIV variants, a phenomenon seen with influenza and Ebola viruses. The cleavage efficiency of the influenza virus hemagglutinin glyco-protein is typically hampered by a suboptimal furin cleavage site; however, two hemagglutinin subtypes that cause signifi-cant morbidity and mortality (H5 and H7) both possess the
optimal polybasic cleavage motif (28, 53, 70). With Ebola virus, all known lethal strains possess a canonical furin cleavage site while the nonlethal Reston strain possesses a suboptimal furin cleavage site (23, 70). Although these examples demonstrate ways in which the alteration of the cleavage site directly im-pacts the phenotype, structural changes that impact accessibil-ity for cleavage may be expected to have similar effects.
An important observation from these data was that an in-crease in the amount of cleaved Env correlated with inin-creased resistance to macaque plasma and a trend toward increased resistance to heterologous HIVIG. Testing of the pseudovi-ruses against several known MAbs and sCD4 further revealed an enhanced ability of variants that arose over time to become more resistant to neutralization than the WT. Importantly, several of the variants demonstrated enhanced resistance to MAbs directed against the MPR of gp41, despite the fact that the pseudoviruses all possessed the same gp41 sequence and differed only in regions of gp120 not previously known to influence Env cleavage. This finding further demonstrates that changes in gp120 can mediate the sensitivity of gp41 to neu-tralization and is in agreement with the findings in a previous report which demonstrated that specific glycans on gp120 can impact sensitivity to neutralization by gp41-directed MAbs (50). It has been argued by other groups (50) that the change in sensitivity to neutralization is likely due to steric hindrance of the epitope, while our data suggest an alternate explanation for modified cleavage. Overall, we saw that the majority of pseudoviruses exhibited increased resistance and that a given pseudovirus possessed a fairly broad trend of resistance: resis-tance to NAbs in plasma tended to correspond with resisresis-tance to HIVIG, sCD4, IgG1b12, 4E10, and 2F5.
The one exception in this study was neutralization by the MAb 2G12. In direct contrast to the results for all other neu-tralizing ligands tested, higher levels of Env cleavage corre-lated with increased sensitivity to 2G12. The epitope of this antibody comprises three central glycans and two peripheral glycans that are less crucial for recognition of the antibody (78). All of the pseudoviruses, including the WT, possessed the three central glycans and one of the peripheral glycans. How-ever, the peripheral glycan N386 was not present on the WT. For the several pseudoviruses with this glycan present, 2G12 neutralization was enhanced. The MAb 2G12 is a highly unique antibody, exhibiting an unusual crossover of the Fab fragments, which creates an antigen recognition site capable of detecting densely packed glycans (78). It is therefore unlikely that the macaques in this study possessed antibodies that rec-ognized the 2G12 epitope, and thus, the virus was not under selective pressure to limit the glycans that make up the epitope. This was nonetheless a unique situation in which the epitope was known to be directly affected in many of the pseudoviruses. This exception underscores that even though there may be an upstream mechanism with broad effects on neutralization, di-rect changes to specific epitopes are an important means of escape. Nevertheless, the lack of correlation between 2G12 and other modalities of neutralization tested in this study un-derscores the fact that mutations that allow the escape of a specific epitope are not broadly applicable. It is arguably evo-lutionarily favorable for the HIV to utilize more-global mech-anisms for escape.
The accessibility of the CD4bs and MPR represents a
13046 BLAY ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
marker for the structural configuration of the Env. The in-creased resistance of the described variants to sCD4 and MAb b12 may have resulted from the perturbation of the CD4bs. Likewise, the increased resistance to 4E10 and 2F5 may have resulted from alterations in the accessibility of the MPR. How-ever, it seems unlikely that an increase in properly processed Env would lead to a decrease in the structural integrity of the Env. We therefore favor an alternate explanation. Several lines of evidence suggest that the ability of an antibody to be neu-tralizing rests simply upon the ability of that antibody to rec-ognize functional, oligomeric Env (58, 60, 61). This concept was elegantly demonstrated when an artificial epitope was en-gineered into the HIV V4 loop and a MAb directed against the epitope was shown to potently neutralize the virus (64). Only one functional Env spike is necessary to mediate infection (82), and only one NAb need bind a spike for the entire spike to be neutralized (81). If both avidity and epitope presentation are held constant, the more functional spikes a given virion has, the more NAb will be required to neutralize all spikes. For example, a virion with two spikes of which one is functional will be neutralized by a single antibody 50% of the time. In con-trast, a virion with two of two spikes that are functional will be resistant to neutralization by a single antibody, since one func-tional spike will always remain unbound. Therefore, the virion with more functional spikes will require a higher titer of anti-body, i.e., be more resistant to neutralization. It has been demonstrated previously that viruses that evolve within a pa-tient often require higher concentrations of plasma for neu-tralization (26, 54, 68, 80). Changes in epitopes clearly play an important role in mediating escape from NAbs that develop over time (14, 52). Increases in the number of functional spikes per virion may serve as another form of escape.
It remains unclear precisely how changes in Env modify cleavage efficiency. As mentioned previously, different variants exhibit a range of cleavage efficiencies even when all variants possess identical cleavage sites and are tested in the same system (6, 58). This observation is further supported by data from this study, as all 20 variants possessed the same furin cleavage site yet a range of cleavage efficiencies was observed. However, in this study, the entire cleavage sites and gp41 regions in all variants were identical, and therefore, the effects on cleavage can be attributed to features of gp120. The ques-tion remains as to what specifically in gp120 played the prom-inent role in enhancing Env cleavage. Lentivirus envelopes are heavily glycosylated, and changes in the number and positions of carbohydrates have been shown previously to play an im-portant role in mediating escape from NAbs (7, 12, 41, 80). However, carbohydrates are also known to affect protein fold-ing and, thus, may alter the accessibility of the cleavage site by producing more tightly packed gp120 (34). Support for this model comes from work by Reynard et al. (65) in which the removal of certain key glycans or clusters of glycans impaired the processing of Env. The modification of complex PNG sites may also lengthen the duration of Env transit through the Golgi network, thereby increasing the availability of Env to furin. In addition, the location of key glycans may provide steric hindrance to the accessibility of furin to its substrate. Though enticing, PNG site changes were not the only differ-ence between these variants and the WT; notably, changes in charged residues were also common. Additional studies are
currently under way to further elucidate how specific changes in Env may affect processing.
Previous studies have documented that HIV can evolve to simultaneously escape NAbs and become more infectious (46, 48, 59). Leavitt and colleagues (46) presented evidence that several changes in gp120 can alter structure and thereby me-diate this dual phenotype. In this study, we provide additional evidence that several different and, as far as we can determine, unrelated changes in gp120 correlate with increased infectivity and increased resistance to neutralization. We further demon-strate that these phenotypes may be attributed in part to changes in Env cleavage that are mediated by characteristics of the gp120 subunit of Env. These findings have implications for Env immunogen selection and design, and they suggest that a more comprehensive analysis of the contributions of individual mutations to these phenotypes is warranted.
ACKNOWLEDGMENTS
This study was supported by PHS grant P01-AI54564 (N.L.H.), the University of Washington Center for AIDS Research Immunology Core (P30-AI27757), a Center for AIDS Research predoctoral fellow-ship (P30-AI27757 [W.M.B.]), a grant from the Murdock Charitable Trust (N.L.H.), and donations from the James B. Pendleton Charitable Trust. Virus stocks, MAbs, and cell lines used in neutralization assays were obtained from the NIH AIDS Research and Reference Reagent Program.
We thank Nicole Doria-Rose and Catherine Blish for critical re-views of the manuscript and Shiu-Lok Hu and Jaisri Lingappa for helpful discussions.
REFERENCES
1.Affranchino, J. L., and S. A. Gonzalez.2006. Mutations at the C-terminus of the simian immunodeficiency virus envelope glycoprotein affect gp120-gp41
stability on virions. Virology347:217–225.
2.Barnett, S. W., S. Lu, I. Srivastava, S. Cherpelis, A. Gettie, J. Blanchard, S. Wang, I. Mboudjeka, L. Leung, Y. Lian, A. Fong, C. Buckner, A. Ly, S. Hilt, J. Ulmer, C. T. Wild, J. R. Mascola, and L. Stamatatos.2001. The ability of an oligomeric human immunodeficiency virus type 1 (HIV-1) envelope an-tigen to elicit neutralizing antibodies against primary HIV-1 isolates is im-proved following partial deletion of the second hypervariable region. J.
Vi-rol.75:5526–5540.
3.Beddows, S., M. Franti, A. K. Dey, M. Kirschner, S. P. N. Iyer, D. C. Fisch, T. Ketas, E. Yuste, R. C. Desrosiers, P. J. Klasse, P. J. Maddon, W. C. Olson, and J. P. Moore.2007. A comparative immunogenicity study in rabbits of disulfide-stabilized, proteolytically cleaved, soluble trimeric human immuno-deficiency virus type 1 gp140, trimeric cleavage-defective gp140 and
mono-meric gp120. Virology360:329–340.
4.Bess, J. W., Jr., R. J. Gorelick, W. J. Bosche, L. E. Henderson, and L. O. Arthur.1997. Microvesicles are a source of contaminating cellular proteins
found in purified HIV-1 preparations. Virology230:134–144.
5.Binley, J. M., R. W. Sanders, B. Clas, N. Schuelke, A. Master, Y. Guo, F. Kajumo, D. J. Anselma, P. J. Maddon, W. C. Olson, and J. P. Moore.2000. A recombinant human immunodeficiency virus type 1 envelope glycoprotein complex stabilized by an intermolecular disulfide bond between the gp120 and gp41 subunits is an antigenic mimic of the trimeric virion-associated
structure. J. Virol.74:627–643.
6.Binley, J. M., R. W. Sanders, A. Master, C. S. Cayanan, C. L. Wiley, L. Schiffner, B. Travis, S. Kuhmann, D. R. Burton, S.-L. Hu, W. C. Olson, and J. P. Moore.2002. Enhancing the proteolytic maturation of human
immu-nodeficiency virus type 1 envelope glycoproteins. J. Virol.76:2606–2616.
7.Blay, W. M., S. Gnanakaran, B. Foley, N. A. Doria-Rose, B. T. Korber, and N. L. Haigwood.2006. Consistent patterns of change during the divergence of human immunodeficiency virus type 1 envelope from that of the inocu-lated virus in simian/human immunodeficiency virus-infected macaques.
J. Virol.80:999–1014.
8.Bourinbaiar, A. S.1994. The ratio of defective HIV-1 particles to
replica-tion-competent infectious virions. Acta Virol.38:59–61.
9.Buchacher, A., R. Predl, K. Strutzenberger, W. Steinfellner, A. Trkola, M. Purtscher, G. Gruber, C. Tauer, F. Steindl, A. Jungbauer, et al. 1994. Generation of human monoclonal antibodies against HIV-1 proteins: elec-trofusion and Epstein-Barr virus transformation for peripheral blood
lym-phocyte immortalization. AIDS Res. Hum Retrovir.10:359–369.
10.Burton, D. R., C. F. Barbas III, M. A. Persson, S. Koenig, R. M. Chanock,
on November 8, 2019 by guest
http://jvi.asm.org/
and R. A. Lerner.1991. A large array of human monoclonal antibodies to type 1 human immunodeficiency virus from combinatorial libraries of
asymp-tomatic seropositive individuals. Proc. Natl. Acad. Sci. USA88:10134–10137.
11.Burton, D. R., J. Pyati, R. Koduri, S. J. Sharp, G. B. Thornton, P. W. Parren, L. S. Sawyer, R. M. Hendry, N. Dunlop, P. L. Nara, et al.1994. Efficient neutralization of primary isolates of HIV-1 by a recombinant human
mono-clonal antibody. Science266:1024–1027.
12.Chackerian, B., L. M. Rudensey, and J. Overbaugh.1997. Specific N-linked and O-linked glycosylation modifications in the envelope V1 domain of simian immunodeficiency virus variants that evolve in the host alter
recog-nition by neutralizing antibodies. J. Virol.71:7719–7727.
13.Chen, B., E. M. Vogan, H. Gong, J. J. Skehel, D. C. Wiley, and S. C. Harrison.2005. Structure of an unliganded simian immunodeficiency virus
gp120 core. Nature433:834–841.
14.Dacheux, L., A. Moreau, Y. Ataman-Onal, F. Biron, B. Verrier, and F. Barin.
2004. Evolutionary dynamics of the glycan shield of the human immunode-ficiency virus envelope during natural infection and implications for
expo-sure of the 2G12 epitope. J. Virol.78:12625–12637.
15.Davis, M. R., J. Jiang, J. Zhou, E. O. Freed, and C. Aiken.2006. A mutation in the human immunodeficiency virus type 1 Gag protein destabilizes the
interaction of the envelope protein subunits gp120 and gp41. J. Virol.80:
2405–2417.
16.Dey, A. K., K. B. David, P. J. Klasse, and J. P. Moore.2007. Specific amino acids in the N-terminus of the gp41 ectodomain contribute to the stabiliza-tion of a soluble, cleaved gp140 envelope glycoprotein from human
immu-nodeficiency virus type 1. Virology360:199–208.
17.Dorfman, T., F. Mammano, W. A. Haseltine, and H. G. Gottlinger.1994. Role of the matrix protein in the virion association of the human
immuno-deficiency virus type 1 envelope glycoprotein. J. Virol.68:1689–1696.
18.Doria-Rose, N. A., C. C. Pierce, M. T. Hensel, W. F. Sutton, N. Sheikh, P. Polacino, L. Kuller, Y. D. Zhu, S. L. Hu, D. Anderson, and N. L. Haigwood.
2003. Multigene DNA prime-boost vaccines for SHIV89.6P. J. Med.
Prima-tol.32:218–228.
19.Dubay, J., S. Dubay, H. Shin, and E. Hunter.1995. Analysis of the cleavage site of the human immunodeficiency virus type 1 glycoprotein: requirement
of precursor cleavage for glycoprotein incorporation. J. Virol.69:4675–4682.
20.Dubay, J. W., S. J. Roberts, B. H. Hahn, and E. Hunter.1992. Truncation of the human immunodeficiency virus type 1 transmembrane glycoprotein
cy-toplasmic domain blocks virus infectivity. J. Virol.66:6616–6625.
21.Earl, P. L., W. Sugiura, D. C. Montefiori, C. C. Broder, S. A. Lee, C. Wild, J. Lifson, and B. Moss.2001. Immunogenicity and protective efficacy of
oligomeric human immunodeficiency virus type 1 gp140. J. Virol.75:645–
653.
22.Falcigno, L., R. Oliva, G. D’Auria, M. Maletta, M. Dettin, A. Pasquato, C. Di Bello, and L. Paolillo.2004. Structural investigation of the HIV-1 envelope glycoprotein gp160 cleavage site 3: role of site-specific mutations.
Chembio-chem5:1653–1661.
23.Feldmann, H., V. E. Volchkov, V. A. Volchkova, and H. D. Klenk.1999. The glycoproteins of Marburg and Ebola virus and their potential roles in
patho-genesis. Arch. Virol. Suppl.15:159–169.
24.Fenouillet, E., and J. C. Gluckman.1992. Immunological analysis of human immunodeficiency virus type 1 envelope glycoprotein proteolytic cleavage.
Virology187:825–828.
25.Fouchier, R. A., B. E. Meyer, J. H. Simon, U. Fischer, and M. H. Malim.
1997. HIV-1 infection of non-dividing cells: evidence that the amino-termi-nal basic region of the viral matrix protein is important for Gag processing
but not for post-entry nuclear import. EMBO J.16:4531–4539.
26.Freed, E. O., and M. A. Martin.1996. Domains of the human immunodefi-ciency virus type 1 matrix and gp41 cytoplasmic tail required for envelope
incorporation into virions. J. Virol.70:341–351.
27.Gabuzda, D. H., A. Lever, E. Terwilliger, and J. Sodroski.1992. Effects of deletions in the cytoplasmic domain on biological functions of human
im-munodeficiency virus type 1 envelope glycoproteins. J. Virol.66:3306–3315.
28.Garten, W., and H.-D. Klenk.1999. Understanding influenza virus
pathoge-nicity. Trends Microbiol.7:99–100.
29.Gluschankof, P., I. Mondor, H. R. Gelderblom, and Q. J. Sattentau.1997. Cell membrane vesicles are a major contaminant of gradient-enriched
hu-man immunodeficiency virus type-1 preparations. Virology230:125–133.
30.Gorny, M. K., L. Stamatatos, B. Volsky, K. Revesz, C. Williams, X.-H. Wang, S. Cohen, R. Staudinger, and S. Zolla-Pazner.2005. Identification of a new quaternary neutralizing epitope on human immunodeficiency virus type 1
virus particles. J. Virol.79:5232–5237.
31.Hall, T.2005. BioEdit, version 5.0.9 ed. Ibis Therapeutics, Carlsbad, CA. 32.Hallenberger, S., V. Bosch, H. Angliker, E. Shaw, H.-D. Klenk, and W.
Garten.1992. Inhibition of furin-mediated cleavage activation of HIV-1
glycoprotein gpl60. Nature360:358–361.
33.Hatta, M., P. Gao, P. Halfmann, and Y. Kawaoka.2001. Molecular basis for
high virulence of Hong Kong H5N1 influenza A viruses. Science293:1840–
1842.
34.Helenius, A., and M. Aebi.2001. Intracellular functions of N-linked glycans.
Science291:2364–2369.
35.Herrera, C., P. J. Klasse, E. Michael, S. Kake, K. Barnes, C. W. Kibler, L.
Campbell-Gardener, Z. Si, J. Sodroski, J. P. Moore, and S. Beddows.2005. The impact of envelope glycoprotein cleavage on the antigenicity, infectivity, and neutralization sensitivity of Env-pseudotyped human immunodeficiency
virus type 1 particles. Virology338:154–172.
36.Herrera, C., C. Spenlehauer, M. S. Fung, D. R. Burton, S. Beddows, and J. P. Moore.2003. Nonneutralizing antibodies to the CD4-binding site on the gp120 subunit of human immunodeficiency virus type 1 do not interfere with
the activity of a neutralizing antibody against the same site. J. Virol.77:1084–
1091.
37.Iwatani, Y., K. Kawano, T. Ueno, M. Tanaka, A. Ishimoto, M. Ito, and H. Sakai.2001. Analysis of dominant-negative effects of mutant Env proteins of
human immunodeficiency virus type 1. Virology286:45–53.
38.Karlsson, G. B., F. Gao, J. Robinson, B. Hahn, and J. Sodroski.1996. Increased envelope spike density and stability are not required for the neutralization resistance of primary human immunodeficiency viruses. J.
Vi-rol.70:6136–6142.
39.Kirschner, M., V. Monrose, M. Paluch, N. Techodamrongsin, A. Rethwilm, and J. P. Moore.2006. The production of cleaved, trimeric human immu-nodeficiency virus type 1 (HIV-1) envelope glycoprotein vaccine antigens and infectious pseudoviruses using linear polyethylenimine as a transfection
reagent. Protein Expr. Purif.48:61–68.
40.Kothe, D. L., J. M. Decker, Y. Li, Z. Weng, F. Bibollet-Ruche, K. P. Zammit, M. G. Salazar, Y. Chen, J. F. Salazar-Gonzalez, Z. Moldoveanu, J. Mestecky, F. Gao, B. F. Haynes, G. M. Shaw, M. Muldoon, B. T. M. Korber, and B. H. Hahn.2007. Antigenicity and immunogenicity of HIV-1 consensus subtype B
envelope glycoproteins. Virology360:218–234.
41.Kraft, Z., N. R. Derby, R. A. McCaffrey, R. Niec, W. M. Blay, N. L. Haigwood, E. Moysi, C. J. Saunders, T. Wrin, C. J. Petropoulos, M. J. McElrath, and L. Stamatatos.2007. Macaques infected with a CCR5-tropic simian/human immunodeficiency virus (SHIV) develop broadly reactive anti-HIV
neutral-izing antibodies. J. Virol.81:6402–6411.
42.Kumar, S., K. Tamura, and M. Nei.2004. MEGA 3: integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief.
Bioinformatics5:150–163.
43.Kwong, P. D., M. L. Doyle, D. J. Casper, C. Cicala, S. A. Leavitt, S. Majeed, T. D. Steenbeke, M. Venturi, I. Chaiken, M. Fung, H. Katinger, P. W. Parren, J. Robinson, D. Van Ryk, L. Wang, D. R. Burton, E. Freire, R. Wyatt, J. Sodroski, W. A. Hendrickson, and J. Arthos.2002. HIV-1 evades anti-body-mediated neutralization through conformational masking of
receptor-binding sites. Nature420:678–682.
44.Kwong, P. D., R. Wyatt, J. Robinson, R. Sweet, J. Sodroski, and W. Hendrickson.1998. Structure of an HIV-1 gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature
393:648–659.
45.Lambele, M., B. Labrosse, E. Roch, A. Moreau, B. Verrier, F. Barin, P. Roingeard, F. Mammano, and D. Brand.2007. Impact of natural polymor-phism within the gp41 cytoplasmic tail of human immunodeficiency virus type 1 on the intracellular distribution of envelope glycoproteins and viral
assembly. J. Virol.81:125–140.
46.Leavitt, M., E. J. Park, I. A. Sidorov, D. S. Dimitrov, and G. V. Quinnan, Jr.
2003. Concordant modulation of neutralization resistance and high infectiv-ity of the primary human immunodeficiency virus type 1 MN strain and
definition of a potential gp41 binding site in gp120. J. Virol.77:560–570.
47.Long, E. M., S. M. Rainwater, L. Lavreys, K. Mandaliya, and J. Overbaugh.
2002. HIV type 1 variants transmitted to women in Kenya require the CCR5 coreceptor for entry, regardless of the genetic complexity of the infecting
virus. AIDS Res. Hum Retrovir.18:567–576.
48.Louder, M. K., A. Sambor, E. Chertova, T. Hunte, S. Barrett, F. Ojong, E. Sanders-Buell, S. Zolla-Pazner, F. E. McCutchan, J. D. Roser, D. Gabuzda, J. D. Lifson, and J. R. Mascola.2005. HIV-1 envelope pseudotyped viral vectors and infectious molecular clones expressing the same envelope gly-coprotein have a similar neutralization phenotype, but culture in peripheral blood mononuclear cells is associated with decreased neutralization
sensi-tivity. Virology339:226–238.
49.McBurney, S. P., K. R. Young, and T. M. Ross.2007. Membrane embedded HIV-1 envelope on the surface of a virus-like particle elicits broader immune
responses than soluble envelopes. Virology358:334–346.
50.McCaffrey, R. A., C. Saunders, M. Hensel, and L. Stamatatos.2004. N-linked glycosylation of the V3 loop and the immunologically silent face of gp120 protects human immunodeficiency virus type 1 SF162 from
neutral-ization by anti-gp120 and anti-gp41 antibodies. J. Virol.78:3279–3295.
51.McCune, J. M., L. B. Rabin, M. B. Feinberg, M. Lieberman, J. C. Kosek, G. R. Reyes, and I. L. Weissman.1988. Endoproteolytic cleavage of gp160 is
required for the activation of human immunodeficiency virus. Cell53:55–67.
52.Mo, H., L. Stamatatos, J. E. Ip, C. F. Barbas, P. W. Parren, D. R. Burton, J. P. Moore, and D. D. Ho.1997. Human immunodeficiency virus type 1 mutants that escape neutralization by human monoclonal antibody IgG1b12.
J. Virol.71:6869–6874.
53.Molloy, S. S., L. Thomas, C. Kamibayashi, M. C. Mumby, and G. Thomas.
1998. Regulation of endosome sorting by a specific PP2A isoform. J. Cell
Biol.142:1399–1411.
54.Moog, C., H. J. Fleury, I. Pellegrin, A. Kirn, and A. M. Aubertin.1997.
13048 BLAY ET AL. J. VIROL.