0022-538X/82/070191-10$02.00/0
Sequence of
the Long
Terminal
Repeat and Adjacent
Segments
of
the
Endogenous
Avian Virus
Rous-Associated
Virus
0
STEPHEN H. HUGHES
ColdSpring Harbor Laboratory, Cold Spring Harbor, New York 11724 Received 9 December1981/Accepted 9 March 1982
Rous-associatedvirus 0(RAV-0), anendogenouschicken virus, does not cause
disease when inoculated into susceptible domestic chickens. An infectious
unintegrated circularRAV-0 DNA was molecularly cloned, and thesequence of
the long terminal repeat (LTR) and adjacent segments was determined. The
sequenceof the LTRwasfoundto bevery similar to that ofreplication-defective
endogenous virusEV-1.LiketheEV-1 LTR, the RAV-0 LTR is smaller (278 base
pairs instead of 330) than the LTRs ofthe oncogenic members of the avian sarcomavirus-avian leukosis virusgroup. There is,however, significant
homolo-gy. The most striking differences are in the U3 region of the LTR, and in this
region thereare aseries ofsmall segments present in theoncogenicviruses which
areabsent in RAV-0.Thesedifferencesin theU3 regionof the LTRcouldaccount
forthe differences in the oncogenic potential of RAV-0 and the avian leukosis
viruses. I also compared the regions adjacent to the RAV-0 LTR with the
availableaviansarcomavirus sequences. Asegment ofapproximately200bases
tothe rightof the LTR (towardgag)isalmost identical in RAV-0 and thePragueC
strainofRoussarcomavirus. The segmentof RAV-0 which liesbetweenthe end
of theenvgeneandU3is approximately190 bases inlength. Essentiallythisentire
segmentis present between env and src inthe Schmidt-RuppinAstrainofRous
sarcomavirus.Mostofthissegmentisalsopresent betweenenv and src in Prague
C; however, in Prague C there isanapparent deletion of 40 bases in theregion
adjacent to env. In Schmidt-Ruppin A, but not in Prague C, about half of this segment is also present between src and the LTR. This arrangement has
implications for the mechanism by which src was acquired. The region which
encoded the gp37portionofenvappears tobeverysimilar in RAV-0 and theRous
sarcoma viruses. However, differences at the very end ofenv imply that the
carboxy termini of RAV-0, Schmidt-Ruppin A, and Prague C gp37s are
signifi-cantly different. Theimplications of theseobservationsareconsidered.
Rous-associated virus 0(RAV-0), an
endoge-nous chicken virus spontaneously produced by
certainlines ofinbred whiteleghornchickens, is
closely relatedtotheother members oftheavian
sarcomavirus-avianleukosis virus
(ASV-ALV)
group. It has, however, a distinct host range,
defined by the virus-encoded envelope gene,
and,incontrast totheotherASV-ALV viruses,
isnononcogenic eveninlong-term(over1year)
invivo infections of domestic chickens (7, 20).
The ASV-ALV viruses transform cells
by
atleasttwodistinctmechanisms: certain
oncogen-ic viruses thatcandirectlyandinevitably cause
transformationcarryhomologsofcellulargenes
(oncogenes) (1); the ALV viruses which lack
oncogenes can, at least insomecases,activate
resident cellular oncogenes after
integrating
nearby(14, 21, 22a,23). Asaconsequence, the
ALV viruses are less efficient in causing
neo-plastic diseases than are retroviruses carrying
oncogenes. RAV-0, which does not carry a
cellularoncogene, is apparently also incapable
of activating cellular oncogenes in domestic chickens by insertion since it does not cause
tumors in these birds. There are two
simple
hypotheses: that the RAV-0 proviruses do not
integrate in the same places that ALV
provi-rusesintegrate, i.e., nearthe cellular oncogene c-myc, or that RAV-0provirusescanintegratein
thesamesites, butfailtoactivate c-myc.
What-ever model is correct, there must be some
underlying difference between the
oncogenic
ALVs and RAV-0.
Sincethelongterminal repeat
(LTR)
region
ofviral DNA appears to contain the sequences
necessaryfor theinitiation of
transcription,
ter-mination of RNA and addition of the
polyade-nylatetail, and is
intimately
involvedinintegra-191
on November 10, 2019 by guest
http://jvi.asm.org/
tion, whichever model proves correct, the crucial difference(s) may reside within the LTR of the virus.
By T1 oligonucleotide fingerprinting (3), hy-bridization (22), and analysis with restriction
enzymes (25), the major differences detected
between theALVs andRAV-0 lie in the 3' end
of the RNA genome, and studies with in vivo
recombinants suggest, but do not prove, that
thisistheregion responsiblefor the difference in
oncogenic potential between the ALVs and
RAV-0 (6, 24, 29). I have cloned unintegrated
circular RAV-0 DNA and shown that the DNA
gives rise toreplicatingRAV-0 virus upon
trans-fection. Ihavesequencedthe LTR of this DNA and compared the RAV-0 sequence with the
sequences of other members of the ASV-ALV
group. Itis nosurprisethat the LTR of RAV-0 is
most closely relatedtothe LTRof the inactive
endogenous virus EV-1, but isrelated, although
lessclosely,tothe LTRsof the other ASV-ALV
viruses. TheRAV-0 LTR issignificantlyshorter
thanthe LTRofRSV,being278(insteadof330)
base pairs in length. The majordifferences
be-tweenRAV-0 and the other
replication-compe-tentASV-ALVviral DNAs lie in theU3segment
ofthe LTR.Thereareminor differences in other
noncodingregions: inU5, the segment
compris-ing the 5' leader, and in the segment upstream
from U3, whichcorrespondsmorecloselytothe
untranslatedregion ofRSVbetweenenvandsrc
than between src and the LTR. There are also
changes in the end of the env gene, the most
obvious of which appear to cause the RAV-0env
genetoterminate downstream of where the env
gene terminatesin RSV.
MATERIALS AND METHODS
Growth of virus; preparation and cloning of uninte-grated RAV-0 DNA. RAV-0 virus was obtained from Harriet Robinson, and a high-titer stock was grown on line15Bchicken embryo fibroblasts. One liter of virus was concentrated approximately30-fold by centrifuga-tion in a Beckman L19 rotor and used to infect two roller bottles of QT6 cells. Seventy-two hours after infection, viral DNA was prepared by the method of Hirt (15) and treated with RNase and pronase. The supercoiled forms of integrated viral DNA were en-riched by a modification of the acid-phenol extraction procedure of Zasloff et al. (9, 30). The recovery of supercoiled viral DNA was monitored at various stagesin the purification by gel electrophoresis, trans-fer to nitrocellulose, and hybridization with 32P-la-beled viral cDNA. Since mapping experiments did not reveal a restriction enzyme suitable for cloning inX that cleaved the viral genome only once (25), the circular viral DNA was partially digested withEcoRI,
which cuts circular RAV-0 DNA twice. This partially digested DNA was ligated to EcoRI-cleaved Charon 3Alac DNA. The resulting A chimeras were packaged in vitro and plated on DP50 SupF. Approximately 0.1% of the plaques contained viral DNA. We have
characterizedadozenclones,oneof which appears to containanentire copy of the viral genome andcanbe rescuedbytransfection of the cloned DNA into chick-encells. The clone(R8) has been mapped with restric-tion endonucleases and the transfer procedure of Southern (27), and it apparently contains a normal copyof theviral genome. R8wascleavedto
comple-tion with HindIlI and EcoRI and subcloned into pBR322. A clone was derived that begins in the
HindlIlsite inenvand ends in theEcoRIsitenearthe end of gag. DNA from this clone was digested to
completion withHindllland treated with Ba131
(Be-thesda Research Laboratories). The ends of the Bal-treated DNAwererepairedwith the Klenowfragment of Poll and ligated to ClaI linkers (Collaborative Research, Inc.). A series of cloneswasderived with
ClaIsites distributed every 200to300 basepairs in the region between Hindlll and the LTR.
Sequence determination.DNAfrom the clones with ClaI linkers insertedwasdigestedwithClaIandSstI
and labeled in thedigestionmix with[o-32P]dCTP(300
Ci/mmol) and the Klenow fragment of PolI. The resultingfragments,labeleduniquelyattheClaIsite, werefractionatedon a1%agarosegel, electroeluted,
purified bychromatography onDEAE-Sephacel,and
sequenced bythechemicalproceduresof Maxam and Gilbert(19).This sequencewasconfirmedby isolating
a HindIlI to SstI fragment from R8, digesting to
completionwithHinfl, labelingtheresulting fragments
with the Klenow fragment of Poll, separating the
labeledstrandsonapolyacrylamide geland sequenc-ing bothseparatedstrands,andbysequencingin both
directions from the BstEII site in theprimer-binding siteadjacenttothe LTR.
RESULTS
Sequence of the RAV-O LTR and adjacent
regions. TheRAV-0LTRis 278 basepairs long,
smaller than that of any other known
replication-competent retrovirus. Figure 1 shows the
se-quenceof theLTRand theadjacentregions. The
RAV-0 LTR has the structural features that
characterize all retrovirus LTRs (H. E.Varmus,
R. Swanstrom, and D. Baltimore, in J. Coffin,
N. Teich, H. E. Varmus, and R. Weiss, ed.,
Molecular Biology ofTumor Viruses: RNA
Tu-mor Viruses, in press). The sequence
TATA-TAA ispresent at a position 23 bases from the
site where RNA was initiated (Fig. 1). The
sequence usually associated with
polyadenyla-tion, AATAAA, is located 24 bases before the
apparent site ofpolyadenylation. A polypurine
tract(AGAGAGGGGA) is located immediately
adjacent to the left end of the LTR. The
se-quence of theprimer-binding site would suggest
that, like Rous sarcoma virus (RSV) (5, 12),
RAV-0uses a tRNA trp primer. The ends of the
RAV-0 LTR form a small imperfect inverted
repeat (six of the first seven and seven of the
first nine base pairs matched; see Fig. 1). In
addition to these general structural features
present in allretrovirus LTRs, the RAV-0 LTR
showsconsiderable sequence homology with the
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 43, 1982
GCTTGTAGTT
CAACATCA
A
TT~GTPTEEAG
AACACAGGTC
C I AC G
A
I
T
GG+GC
HinfI
E
I48
E
GCA
BstEII PMB
EV-1 sequence
1CGC
NGU
S8tI
C ST9G
SEQUENCES IN
RAV-O
193t
HEHHH2CCC ~HCGCTC4ATI
HinfI
TIGTAG
C~GEG
TTAGIGA G TEI
AAC TAICCC AA CA
© Ppr U3begns
I~~~G
'TCG~~~~~~
0TATATCTG A) ~~~u5beginls
ATTCG IcCC A T4 GTCCA
NTG
CGIC4XC T ATU H
CH
TendsI
GGGXGVIGG8
AEIGCGX;T4
999AC9%999G AT99%99999
GACTTTCCTTG
IcGI
EIA
cc
CGCACCG9IGCGP
~T~9TT%9~
GAAAAGCG4TT
I~E~
AATT9AST9T
GTCCG
AT9G4GHinfI
ITTAA G|CTA TAG
CA
C
A9~9:AkIC:k9
GCAAGACAT
CGTTCTGTAA
T
CGA
c c
cATT
ATCT C CGT CTCA
H~AccI
C
CCGCG
9CCTGCT9
GCTTACGTGTCT
EV-1 sequenoe
lbegins
0
TCTCCG
4 CGC
CCATATGCTC
GTAT
AC GA
Rbegins
99999
9ISCG
c
ie
I GAI
TA
IT TT 4
LR9BIA
CTGC Ia98 8 M
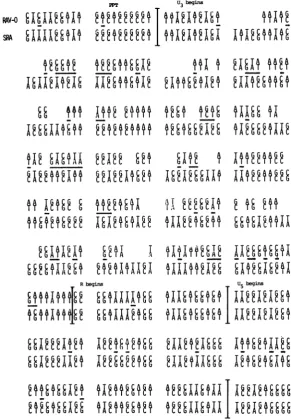
FIG. 1. Sequence of the RAV-O LTR and adjoining regions. The sequence of the RAV-O LTR and the
adjacent regionswasdeterminedbythemethods ofMaxamandGilbert(19).Aseriesof subcloneswasprepared
by inserting ClaI linkers (vertical arrows) afterdigestion with BaIl3 exonuclease for various times. These subclonesprovidedadditionalsequencing startsites in theregionsin and around theRAV-OLTR. Prominent structuralfeatures suchasthebeginningand endof theU3, R,andU5regions,thepolypurine tract(PPT),the
primer-bindingsite(PBS),and the endof theenvgenearemarkedonthe sequence(theTAAcodonatthe end of envisboxed).The inverted repeatsattheendsof theLTRaremarkedbyarrowsunderthe sequence. Differences between theRAV-O sequence and thepublishedsequenceof EV-1 (5)areshownasdifferentbasesgivenabove theRAV-Osequence.Theavailable EV-1 sequence is shorter than the sequenceIhavederived forRAV-O,and theextentof the EV-1 sequence is denotedbyapairof bars above the RAV-O sequence. An A-Tpairpresentin theRAV-Osequence and absent in EV-1is denotedbyanopencircle.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.60.453.71.567.2]RAV-0O
E
E
SRA GINTXC&AT
AS
MA
I GI T
&&
TT
RU
C AIT&TTA
EMU4I
WMW
EPT U3 begins
MMERI
IMPH
E2GEE
M
M Ic
MI M?
c444
Mc&
EM
Rbeginis
EWPIM
HUNME
_ _
&TTAT
&C&&IIACT&
&&CCMTC
IC422
RM
GGICCEIC
ACC&CCT&GCA12T&IGCATE AWMEA4M4
U.beginls
ATT&CTA IC&
ICT&T&HT
UUMHEAT&&
I
MENEM~
MENEM
I
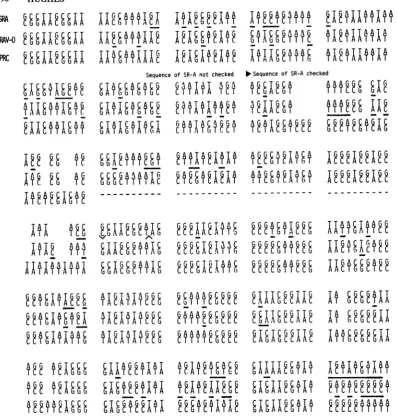
FIG. 2. Comparison of the RAV-O LTR and the LTR of the SR-A strain of RSV (28). The LTR of RAV-O is significantly smaller than the LTR of the other members of the ASV-ALV group. The differences reside principally in the U3 region. I have aligned the U3 regions wherever it was possible, leaving gaps in the RAV-O sequences at positions where the sequences present in SR-A are obviously absent inRAV-O. In some small regions there are no obvious homologies. These regions are indicated by small bars between the RAV-O and SR-Asequences. The distribution of barsbetween the sequences and of gaps within the sequences shows directly the disposition of the differences in the two sequences. The more modest changes inU5and R are alsoindicated in this same fashion. This complete sequence of theRAV-OUsisin perfectagreement with a partial sequence of
RAV-O"strong stop" DNA(Swanstrom, unpublished data). PPTdesignates thepolypurine tract.
LTRs of the other members of the ASV-ALV the LTR ofRAV-O and the LTR of EV-1. Two
group. TheRAV-O LTRis strikingly similar to are transitions, one is a transversion, and the
the LTRof EV-1 (16), adefective endogenous fourth is an additional A-T base pair in the
RAV-virus, and is less closely related to the replica- 0 sequence (Fig. 1). This additional A-Tpair,
tion-competent members of the ASV-ALV near the left end ofU3, is part of a sequence
group. Thereare onlyfour differences between which gave, when the sequence was derived in
1102
PUMP
HUI IM
2402M
.
MEE 14
14
EE21442
MICIMIC2
4 4 . 2 2 . . 2 2
E
11
MR
E
IIE MW
MMU
.HHHM
'f
AA
T.i
HICUP
I 4
4 E
E I
E E
.
I
UUIOEAU
UIAMME
INMEW
.NHEHH
HMMH
MH
.HE
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.94.386.77.498.2]one direction, a serious compression artifact, RAV-0 LTR, or simple mutations. The
T,
oligo-and I initially misread the sequence. Since this nucleotide used by Tsichlis and Coffin (29) to
misreading of the RAV-0 sequence gave a se- distinguish the endogenous and exogenous
vi-quence identical to that published for EV-1, it is ruses(Cx and
Cn,
respectively) derive from thepossible that EV-1 and RAV-0 are identical in region of U3 immediately adjacent to R. The
this region and that there is an error in the relevant sequences were
GCCACCATCAA-published sequence of EV-1 (16). The GCCACCATCAA-published TAACG in RAV-0, which gave rise to the
T,
sequence of EV-1 includes about 60 base pairs to oligonucleotide 08, and GATACAATAACG in
the right of U5; these are identical in RAV-0. Schmidt-Ruppin A (SR-A), which gave rise to
About 25 base pairs of sequence from the left of the
T,
oligonucleotide C (see Fig. 2).the EV-1 U3 have been published; there are two There is a large open reading frame ending 165
changes from RAV-0, both transitions (Fig. 1). bases upstream of the LTR that is likely to
Thesimplest explanation of this similarity is that encode env. Not only is this the only sizable
EV-1, and the other endogenous chicken virus- open reading frame (see Fig. 4), but there is also
es, arose by germ line infection of a virus very considerable homology with the proposed env
closely related to RAV-0 (10, 17, 18, 25). gene of the SR-A (8) and Prague C (Pr-C) (D.
Because the sequences of the EV-1 LTR and Schwartz, personal communication) strains of
the RAV-0 LTR are nearly identical, the rela- RSV. When these sequences are compared in
tionship between the RAV-0 LTR and the RSV the region which corresponds to the carboxy
LTR is very similar to the relationship between terminus of env, a divergence is found. The Pr-C
the LTRs of RSV and EV-1 (16). The RAV-0 env gene is longer than that of SR-A, and the
LTRis 55 base pairs shorter than the RSV LTR, sequence of the RAV-0 env gene, which is closer
and the sequences could be aligned as though to that of SR-A than to that of Pr-C, is the
the RAV-0 LTR had suffered numerous small longest of the three. The inferred amino acid
deletions or, conversely, as though the RSV sequence of these three env genes is given in
LTR had acquired numerous small insertions Fig. 3; that of the DNA is given in Fig. 4.
(Fig. 2). Even when the two sequences are We have sequenced the region between env
aligned optimally, there are still small regions of and src in SR-A and found significant differences
nonhomology.These could be explained as dele- between our data (J. Sorge and S. Hughes,
tions from the RSV LTR, insertions into the unpublished data) and those published by
Czer-RAV- IEL GLY VAL SER
ASPRPD
ILEU GLYSERTR
EARiG
SRAR eV" It of R .E1 itI of of It It
GLY Wi PHE GLY GLY ILEU GLY
GLUW
TI ALA VAL HIS LEJ LEUIt LEJ
LEUJ
It It It it It to It N N N It It M It R It R It oit
It of tL.EU N N N'~ N
LYS GLY LEIJ LE LEJ GLY LEU VAL VAL ILEJ LEU LEJ LEJ VAL
if if i It if of i it NIt I I N R It IN It I NIIt it It o Nf I I N It It
VAL CYS LEUJ PRI) CYS LUE LE GLN IWIE VAL SER SER SER ILEI
i n to It of NI t of PHE
nI
N t it Nf i n II N ItN N M
ET
EU CYS GLY ASN ARGARi LYIsET ILE ASN AS ERSE ER
TYR
HIS THRGLU
YRn N aN Rtt SER ASN of to N R
NIf
N n NASNN
GLU LYSPRO
HISGLY
ILEJ
LYS LYS lEU GlN LYS AU CYS ARG
GIN
PRO GLU ASN GLY ALA VAL [image:5.504.114.398.400.634.2]Ap3
N MET " GLY GLY ALA VALFIG. 3. Inferred amino acid sequences of the ends of the env genes ofSR-A,Pr-C,andRAV-O.Theimplied amino acid sequences of the env genes of SR-A (8), Pr-C (Schwartz,personalcommunication),andRAV-Ohave beenaligned by maximizing homology at the nucleic acid and protein levels. The nucleic acid sequences from the region at the end of env are compared in Fig. 4.
VOL.43,1982
SEQUENCES
IN RAV-0 195on November 10, 2019 by guest
http://jvi.asm.org/
196
SRA
HUGESITEA
J.129VS
ViSSR4A888V
M
RoL.
_
_%99I%
_ _ _ _PRC
TCGGAACGGXT
TI4G4ATIII
ICG4I
4
T4AC4G
4CI4I
Sequence ofSR-A not checked *Sequence of SR-A checked
Cl;CC
ACGA
CAATCGCTCGTG
CTA AT
TCT
TCAGTAGCGcI
T
TU
G
~
C AG
H
IG TT T C
A
_~~~~~~~~~
C
TAT ACcTAT A A
T(
A
TCG
AT
GG~~GC44TTA
C4CGGCCTT~~
TACGAT
TGGGGG
ITATCGICTA --- ---
--I~
A
T
G
A
C e C I I G C C T
IrGC
C C C I I GC9I
4 CG
CC
T
ITIC
TTT
GIICGC4
AT
G
CGCCGICTA
T
CCGCCGGGATTCGG
T
G4CI(
A
G
GGCTGCIATIGCG TIcIT I 444CGCC
C
IAc IEGC
CT [image:6.504.59.457.66.483.2]_ _ _ __
FIG. 4. Comparisonof the sequences fromenvtothe LTR of RAV-O with sequencesfromenvtowardsrcin SR-A and Pr-C. TheregionofRAV-0 from the end ofenvtothe LTRwasalignedwith theendofenvandthe
in-tergenic regionbetweenenvandsrcof bothPr-C(Schwartz,personalcommunication)andSR-A(8). Wehave
checked a portion of the SR-A sequence in thisregion, using subclones derivedfrom the same X clone (9)
sequenced byCzernilofskyetal.(8)and foundsignificantdifferences in theregionbetweenenvandsrc(J.Sorge
andS. Hughes,unpublisheddata).TheSR-A sequencegivenin thefigureisacompositeof thepublisheddata
andourunpublishedrevision. Theportionnotchecked is indicated in thefigure, tothe leftof thearrowhead;
thosethathavebeen checkedareto therightof the arrowhead.Sinceaportionof thesequence foundinRAV-O
and SR-A near the end of the env gene is apparently deleted in the Pr-C sequence (Schwartz, personal
communication),Ihave indicatedthisapparentdeletionbyagapin thePr-C sequencefilled inbyadotted line. Smaller deletions(usuallyasinglebasepair) areshownasgaps. Lack ofhomologybetweenSR-A and RAV-0
andPr-C andRAV-o are indicated by barsbetween theRAV-o andSR-A andthe RAV-0andPr-C sequences.
nilofsky et al. (8) (Fig. 4). Although we did not In the case ofSR-A, the region ofhomology
sequenceenvinSR-A,itispossiblethatsomeof continuesbeyond envand extendsfor about 150
the differences between the SR-A and RAV-0 nucleotides, which are
apparently
noncoding,
sequences aredue to errors. Such errorscannot,
diverging
nearthepointinthe RAV-0sequencehowever, accountforthe
significant
differences where thepolypur
netractadjacenttothe LTRbetween the Pr-C and RAV-0 env
sequences
begins
and in SR-Aabouthalfway
between env(Fig. 4). and src
(Fig.
4). Asimilarcomparison
withPr-CJ.VIROL.
196 HUGHES
on November 10, 2019 by guest
http://jvi.asm.org/
VOL4319 V-'19
RAV--O
CTATT
'AGT
TG5 CT CT
G
T
G G
A TT A G
PRC
T
A
TAA ECTC
A A
C
GGA
GA
C
A C
CTAATCCC
G
c~
~~~~~G
GTTAIGC GAT T GGGC GGCCCGCCT T G
TCATACAT ccCGCTTTccG CCGCcCAACCG
IACATAAG
GCC IIA
TCXC G GC
TA CG GC
AT~GT9EG%GG
TT~GTEE~
TATATTCCGTA C CGA AGCTGCCGA A CAT G
PPT
TCI4CIH;IGH
PCiAAGT
ACCATCCCFIG. 5. Comparison of the sequences between env and the LTR of RAV-O with the sequences between src and the LTR of Pr-C andSR-A. The regions upstream of the LTR are compared for the three viruses. Lack of homology is indicated as a bar between the sequences being compared. The polypurine tract is marked (PPT). Thedivergence of Pr-C from the other two viruses is clearly evident by an examination of the frequency and the extent of the barred region between the RAV-0 and Pr-Csequences.
revealsanapparentdeletion,extending from the end of env for about 40 bases. Beyond this
"deletion" thehomologyresumes,and there isa
polypurinetractinPr-C betweenenvandsrcata
site corresponding in relative position to the
polypurinetractin RAV-0 whichliesadjacentto
the LTR.
It is also possible to comparethe sequences
which lie betweensrcandthe LTR of the RSVs
with the region between env and the LTR of
RAV-0. Again,RAV-0 ismorecloselyrelatedto
SR-A than toPr-C (Fig. 5). There is inSR-A a
large direct repeat which flanks src (8). This repeatwasfoundtobehomologoustopartofthe
region between env and the LTR of RAV-0. A
directcomparison of thesequencesbetweensrc
andtheLTR ofSR-A with the regionbetween
envand the LTR of RAV-0 reveals aregion of
homology beginning at the LTR of about 120
base pairs in length; the last 40 base pairs
(approximately)adjacenttoenvin RAV-0 donot
haveaclearhomologin theregionbetweensrc
and the LTR of SR-A. In Pr-C, the region of
homology is muchshorter; only about the first
20base pairs adjacent to the LTR are found in theregionbetweensrcand the LTRof Pr-C. On
the other side of the LTR, the untranslated
leaderto therightofU5isvery
similar
in RAV-0and Pr-C,althoughthere aredifferences(Fig. 6).
Itispossiblethatthisregionisimportantasthe
recognitionsignalforpackagingRNAinto
viri-ons (26) and that the conservation of the
se-quence inthisregionreflects this requirement.
DISCUSSION
Although RAV-0 is an endogenous
virus,
itreplicatesreasonably well,and thesmallRAV-0
LTR thus contains all of the information
re-quired in a retrovirus LTR. Since the RAV-0
LTR is very similar to the LTR of EV-1, a
defectiveendogenousvirus whichistranscribed
atverylowlevels(13),it is
unlikely
thatthe poortranscription ofEV-1 is due to a defect in the
EV-1 LTR.The fewchangesbetweenthe EV-1
and RAV-0 LTRs are not in
regions
like the"TATAA" box (2), which wecan
recognize
asbeing involvedin
transcription.
Inaddition,
EV-VOL.43, 1982
SEQUENCES
IN RAV-0 197on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.132.373.69.353.2]U5 PBS
PRC
HRACGC214A
I
CTGG
CTCG§ 2
T0RHUI
A
A
242qa%%
G
C
[image:8.504.137.376.77.384.2]T0HH~P
I
A
242248%%
G
GT
FIG.~~~~~~~
6.Coprio of thsqeCe
of thuTrasae
UG
lede seune beteeCGIATGA
and gag in Pr- and4RSE2UH4E
HEHEUEE
HHH
RAV-O. Most of the regions between Us and gag in Pr-C and RAV-O are shown for comparison. Lack of homology is shown by a bar between the sequences.
1 becomestranscriptionallyactive after azacyti-dinetreatment,whichsuggeststhat,under
prop-ercircumstances,the EV-1 LTRcanfunction to
initiatetranscription (11). These data taken
to-gether with earlier studies suggest either that
there maybe contextual constraints on
expres-sion ofproviruses (4)orthat DNAmodification mayplayamajor roleinregulation ofexpression (11), orboth.
Eventhough the RAV-O isacompetent repli-cating virus, it doesnot,despiteaclear homolo-gywith the ALV viruses, induceanyneoplasia
indomesticchickens. Previous studies
suggest-ed, but didnotprove,thatthecrucial difference
inoncogenic potential resides within the LTR (6, 24, 29). Both hybridization andT1 oligonucleo-tide fingerprinting experiments show that the
major differences between the ALVs andRAV-O
areinthe regionatthe 3' endof viralRNA,and
my sequenceanalysishasdemonstrated that the
differencesliemostlywithin the U3 region. The ALVscaninduce leukosis by integratingnextto
the cellular oncogene c-myc and enhancing
c-myctranscription (14, 21, 22a, 23).
RAV-O does not induce leukosis in domestic
chickens; therefore, at some level it does not
duplicateanevent orprocessproduced byALV.
Thereareseveralpossibilities, some morelikely
than others. It is possible the RAV-O cannot
infect theappropriatetargetcell,althoughthis is
unlikely since subgroup E ALVsareoncogenic
(6, 24). It is alsopossible,though unlikely,that
RAV-Oactivatesc-mycbutkills thetargetcell.It
seems more likely either that RAV-O does not
integrate near c-myc orthat RAV-O proviruses
integratednear c-mycdo not enhance
transcrip-tion to a level sufficient for transformation.
Differences intheU3region of the LTRs of ALV
andRAV-Ocould alter the"specificity"of
inte-gration, ifthere is any specificity in retrovirus
integration.Alternatively,the signals for
regula-tionofRNA transcription that reside in the U3
region of the LTR could be responsible for
changes in the ability of RAV-O to activate
adjacentgenessuch as c-myc. Comparing the U3
regions ofALV and RAV-O does not resolve
theseissues, although it is temptingtospeculate
thatwhatever theRAV-OLTRlacks in terms of
on November 10, 2019 by guest
http://jvi.asm.org/
199
sequence may be responsible forthe
concomi-tantloss of oncogenic potential.The fact that the
differences between the ALV and RAV-0 U3s
aredistributedover thewhole U3 region makes a
direct testof this hypothesis difficult.
Do host factors influence the oncogenic
po-tential of the avian viruses, differentiating
be-tween theoncogenic ALVs andRAV-0?A
pre-liminary reportfromWeiss and Frisby suggests
that hostfactorsmayplayakeyrole;they have
found RAV-0 to be tumorigenic in Sonnerat's
jungle fowl (R. Weiss and D. Frisby, in D. F.
Yolm, ed., 10th International Symposium for
Comparative ResearchonLeukemiaand
Relat-ed Diseases,inpress). Althoughthesample size
was small, about half of the RAV-0-infected
birds died ofapparent lymphoid leukosis, and
several more died from "wasting disease."
Even in Sonnerat's jungle fowl, however, the
RAV-0-inducedtumorsappeared on average
sig-nificantly later, and at a lowerfrequency, than
tumorsinducedby the subgroup E recombinant
RAV-60.
Itis not yet clear why RAV-0 is oncogenic in
these particular birdsand not in ordinary
chick-ens, but Sonnerat's jungle fowl completely lack
RAV-0-relatedendogenous viruses, and RAV-0
replicates to a very high titer in these birds.
Whetherone ofthese factors, or others, as yet
undiscovered, distinguishes the oncogenic
sus-ceptibility of chickens and Sonnerat's jungle
fowl, it is clear the differences between the
hosts, as wellas differences between the
virus-es, canprofoundly alter the incidenceof
tumori-genesis. This implies that ahostfactor, or
fac-tors, is involved. It would be particularly
interestingtolearn whether theRAV-0-induced
tumorspresentinSonnerat's jungle fowlarethe
resultofc-mycactivation by RAV-0.
Aside from the LTR sequences, the most
obvious differences between the RSVs which
have been sequenced and theregionof RAV-0
DNAIhavesequencedare attheveryendof the
envgene.If thesequencesattheendsof theenv
genesof Pr-C (Schwartz,
personal
communica-tion), SR-A(8), and RAV-0are
compared,
thereisaclearhomologythatbreaks downatthevery
end of theenvgene.All three envgenes appear
to terminate in different places,
giving
primarytranslation products whichare verydifferent at
theircarboxy termini. Thedramatic differences
in predicted amino acid sequence suggeststhat
the extremecarboxyendofgp37is
probably
not functional.I wassurprisedtofindacomplete
homolog
of theregion betweenenv and the LTRof RAV-0(excluding the polypurine
tract)
in theregion
betweenenvandsrcofthe SR-Astrain of ASV.
Beyondtheapparentdeletionatthe end ofenv,
Pr-C alsocontainsverysimilarsequences.In
Pr-Cthere isevenapolypurine tract present atthe
same position relative to env as the polypurine
tract which is adjacent to the LTR inRAV-0. If
we assume that RAV-0 never contained src,
then this homology presumably is the result of theway src was acquired by the ancestor ofthe
RSVviruses. This antecedent virus presumably
hadconsiderablehomologytoRAV-0,although,
unless there were additional recombination
events, it hada U3region similarto thatofthe
oncogenicmembers of theALV-ASV group.
Itis unlikelythat, when src was first acquired,
it was simply inserted into the untranslated
region betweenenvand the LTR,sinceinSR-A
the src gene is flanked by an essentially
com-plete copy of sequence which in RAV-0 lies
betweenenv and the LTR. Theidea thatsrcwas
notmerely inserted intotheregion betweenenv
and the LTR is reinforced by the presence of
part ofU3 upstreamofsrc in thePr-C and B77
strains of RSV (Mardon, personal
communica-tion;Swanstrom,personal communication). Itis
likely thattwoviral genomes together with
cellu-lar srcwere required to create SR-A. Through
integration or the actions of reverse
transcrip-taseorboth,c-srcwas apparently added to the
end ofaviralgenomewiththeconcomitantloss
of U3. To complete the virus,asecondcopyof
thesequencefound betweenenvand the LTR of
RAV-0wasthenadded to the end of src together
withU3;almostcertainly a second viral genome
wasthe donor of this segment.
The region between the end of U5 and the
beginningofgag appears tobehighly conserved
between Pr-C RSV and RAV-0. The
primer-binding site is identical, which suggests that
RAV-0,like ASV,uses tRNA trp as aprimerfor
DNAsynthesis. The conservation of therestof
this region argues that there is some functional
constraintonwhat is believedtobeanoncoding
region. Experiments witha mutantvirus witha
150-base-pair deletion in this region imply that
theregionisimportantfor efficientpackagingof
RNAinto virions (26), alikely explanation for
theconservation ofthese sequences.
ACKNOWLEDGMENTS
IthankAnn Mutschler for experttechnical assistance and mycolleagues,especiallyJ.Sorge,G.Payne,and J.Fiddes, forhelpful discussions.
This research was supported byaPublic Health Service grant from theNationalInstitutesof Health.
ADDENDUM IN PROOF
Additionalsequencingin theregionof SR-A which encodes thecarboxyterminus ofgp37does notagree withthe datapublishedbyCzernilofskyetal.(8).The newDNA sequence(J. Sorge, unpublisheddata) pre-dictsthatthecarboxyterminus of SR-Agp37isquite similartothecarboxyterminus ofRAV-0 gp37.
on November 10, 2019 by guest
http://jvi.asm.org/
LITERATURE CITED
1. Bishop, J. M. 1981. Enemies within: the genesisof retro-virus oncogenes. Cell 23:5-6.
2. Breathnach, R., and P. Chambon. 1981.Organization and expression of eucaryotic split genes coding forproteins. Annu. Rev.Biochem. 50:349-383.
3. Coffin, J., M. Champion, and F. Chabot. 1978.Nucleotide sequencerelationships between the genomesofan endog-enous and an exogenous avian tumor virus. J. Virol. 28:972-991.
4. Cooper, G. M., and H. M. Temin. 1976. Lack of infectivity of the endogenous avianleukosisvirus-related genes in the DNAof uninfectedchicken cells. J. Virol. 17:422-430. 5. Cordell-Stewart, B., J. M. Taylor, W. Rhode, H. M.
Goodman, and J. M. Bishop. 1974. Transfer RNAs of Rous sarcomavirus and theinitiationof DNAsynthesis by viral reversetranscriptase, p. 117-135. In A. Kaplan (ed.), Viraltransformationandendogenous viruses, Pro-ceedingsof the 1974 Abraham FlexnerSymposium. Aca-demic Press, Inc., New York.
6. Crittenden, L. B., W. S. Hayward, H. Hanafusa, and A.M. Fadly.1980.Inductionof neoplasmsby subgroup E recombinants of exogenous and endogenous avian
retro-viruses(Rous-associatedvirus type 60). J. Virol. 33:915-919.
7. Crittenden,L.B., R.L.Witter,and A. M.Fadly.1979. Lowincidence oflymphoidtumorsin chickens continu-ously producingendogenous virus. Avian Dis. 23:646-653.
8. Czernilofsky,A.P., A. D. Levinson, H. E.Varmus, J. M. Bishop,E.Tischer, and H. M.Goodman. 1980.Nucleotide sequenceofanavian sarcomavirus oncogene (src)and proposedamino acid sequence for geneproduct. Nature (London)287:198-203.
9. DeLorbe,W., P.Luciw,H. M.Goodman, H.E.Varmus, andJ.M.Bishop. 1980.Molecularcloningand character-ization ofavian sarcoma virus circular DNA molecules. J. Virol. 36:50-61.
10. Frisby, D., R. A. Weiss,M.Roussel, and D.Stehelin. 1979. The distribution of endogenous chicken retrovirus
se-quencesin the DNAofgalliform birds does not coincide with avian phylogenetic relationships. Cell 17:623-634.
11. Groudine, M., R. Eisenman, and H. Weintraub. 1981. Chromatin structureof endogenousretroviral genes and activation of an inhibitorof DNA methylation. Nature (London) 292:311-317.
12. Harada, F., R. C. Sawyer, and J. C. Dahlberg. 1975. A primer ribonucleic acid for initiation of in vitro Rous sarcomavirus deoxyribonucleic acid synthesis. J. Biol. Chem.250:3487-3497.
13. Hayward, W. S., S.B.Braverman, and S. M. Astrin. 1980. Transcriptionalproducts and DNA structure of endoge-nousavian proviruses. Cold Spring Harbor Symp. Quant. Biol. 44:1111-1121.
14. Hayward, W. S., B. G. Neel, and S.Astrin. 1981. Activa-tionof a cellular onc gene by promoter insertion in ALV-induced lymphoid leukosis. Nature (London) 290:475-480.
15. Hirt, B.1967.Selectiveextractionofpolyoma DNA from infected mouse cell cultures. J. Mol. Biol. 26:365-369.
16. Hishinuma,F., P. J. DeBona, S. Astrin,and A. M. Skalka. 1981.Nucleotide sequence of the acceptor site and termi-ni of the integrated avian endogenous provirus ev-J:
integration creates a 6 bp repeat of host DNA. Cell 23:155-165.
17. Hughes, S. H., F. Payvar, D.Spector, G. Payne,R. T. Schimke, H. Robinson,J.M.Bishop, and H. E. Varmus. 1979. Heterogeneityofgeneticloci in chickens:analysis of endogenous viraland non-viral genes by cleavageof DNAwithrestriction endonucleases.Cell 18:347-359. 18. Hughes, S., K. Toyoshima, J. M. Bishop, and H. E.
Varmus.1981.Organizationof theendogenousproviruses of chickens:implicationsfororiginandexpression. Virol-ogy108:189-207.
19. Maxam, A., andW.Gilbert. 1980.Sequencingend-labeled DNA with bare-specific chemical cleavages. Methods Enzymol. 65:499-560.
20. Motta, J. V., L.B. Crittenden, H. G.Purchase, U. A. Stone,W.Okazaki, and R. L. Witter. 1975. Low oncogen-icpotentialof avianendogenousRNAtumorvirus infec-tion orexpression.J.Natl. Cancer Inst. 55:685-689. 21. Neel, B. G.,W.S.Hayward, H. C.Robinson,J.Fang,and
S. M. Astrin. 1981. Avianleukosis virus-inducedtumors
have common proviral integration sites and synthesize discretenewRNAs: oncogenesis by promoterinsertion. Cell 23:323-334.
22. Neiman, P., S. Das, D. MacDonnel, and C. McMillin-Helsel. 1977. Organization of the shared and unshared sequences in the genomes of chicken endogenous and
sarcomaviruses. Cell 11:321-329.
22a.Payne, G. S., J. M. Bishop, and H. E. Varmus. 1982. Multiplearrangements ofviral DNA andanactivated host oncogene(c-myc)inbursallymphomas.Nature(London) 295:209-214.
23. Payne,G.S., S.A.Courtneidge,L.B.Crittenden,A.M. Fadly,J. M.Bishop, and H.E. Varmus.1981.Analysisof avian leukosis virus DNA and RNA in bursaltumors:viral geneexpression is not requiredfor maintenance of the
tumorstate.Cell 23:311-322.
24. Robinson, H. L.,M. N.Pearson,D. W. DeSimoneP.N. Tsichils,andJ.M.Coffin. 1980.Subgroup-E avian-leuko-sis-virus-associateddiseasein chickens. ColdSpring Har-borSymp.Quant.Biol. 44:1133-1142.
25. Shank, P., S. Hughes, and H. E. Varmus. 1981.Restriction endonuclease mapping of the DNA of Rous associated virus 0 reveals extensive homology in structure and sequence with avian sarcoma virus DNA. Virology 108:177-188.
26. Shank, P. R., and M. Linial. 1980. Avian oncovirus mutant(Se2lQlb)deficient ingenomic RNA: character-ization ofadeletionin theprovirus.J.Virology 36:450-456.
27. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol.Biol. 98:503-517.
28. Swanstrom, R., W. DeLorbe, J. M. Bishop, and H. E. Varmus. 1981. Sequencing of cloned unintegrated avian
sarcomavirus DNA reveals direct and inverted repeats similar to those in transposable elements. Proc. Natl. Acad.Sci. U.S.A.78:124-128.
29. Tsichlis,P. N.,and J. M. Coffin. 1980. Role of the C region in relative growth rates of endogenous and exogenous avian oncoviruses. Cold Spring Harbor Symp. Quant. Biol. 44:1123-1132.
30. Zasloff, M., G.D.Ginder, and G. Felsenfeld. 1978. A new methodfor the purification of covalently closed circular DNA molecules. Nucleic Acids Res.5:1139-1152.
on November 10, 2019 by guest
http://jvi.asm.org/