JOURNAL OFVIROLOGY, 0022-538X/97/$04.0010
Feb. 1997, p. 1662–1666 Vol. 71, No. 2
Copyrightq1997, American Society for Microbiology
The Sequence Element of the Internal Ribosome Entry Site and a
25-Kilodalton Cellular Protein Contribute to Efficient Internal
Initiation of Translation of Hepatitis C Virus RNA
SHUETSU FUKUSHI,1* CHIE KURIHARA,1NARIHIRO ISHIYAMA,2FUMINORI B. HOSHINO,1
AKIRA OYA,1ANDKAZUHIKO KATAYAMA1
Basic Research Division, BioMedical Laboratories, Inc., 1361-1, Matoba Kawagoe-shi, Saitama 350-11,1and The Third Department of Internal Medicine, Kyorin University
School of Medicine, 6-20-2, Shinkawa Mitaka-shi, Tokyo 181,2Japan
Received 1 July 1996/Accepted 15 October 1996
Translation of hepatitis C virus (HCV) RNA is initiated by internal entry of ribosomes into the 5*noncoding
region (NCR). This process depends on genomic elements within the 5*NCR called the internal ribosome entry
site (IRES) and may involve host factors. The alpha-branch structure (nucleotides 47 to 67) of the HCV IRES
is considered acis-acting element critical for translation initiation because it is indispensable for translation
in vitro (S. Fukushi, K. Katayama, C. Kurihara, N. Ishiyama, F. B. Hoshino, T. Ando, and A. Oya, Biochem. Biophys. Res. Commun. 199:425–432, 1994). In order to further characterize the function of the alpha-branch, we determined whether sequence exchange within the alpha-branch had any effect on translation initiation. An in vitro translation study revealed that the stem sequences of this region played an important role in efficient
IRES function. In addition to several HeLa cell proteins, which had a binding affinity for the 5*NCR, a novel
25-kDa protein that specifically interacted with the HCV IRES was discovered. The binding affinity of the
25-kDa protein for the 5* NCR was correlated with the efficiency of translation initiation of HCV RNA,
indicating a critical role for the 25-kDa protein in HCV translation.
Extensive sequence analyses showed that the 59 noncoding region (NCR) of the hepatitis C virus (HCV) genome is highly conserved among different HCV isolates (3, 4, 16). This region consists of 330 or 341 nucleotides (nt) and has several short open reading frames (6, 12). A complex secondary structure has been proposed for the HCV 59NCR (2). These features, coupled with experimental evidence derived from expression studies of dicistronic mRNA in vitro and in vivo, indicate that translation of HCV RNA is initiated by binding of the ribo-some to an internal riboribo-some entry site (IRES) (5, 26, 27).
The translation strategy used by HCV is similar to that employed by members of the family Picornaviridae. In picor-naviruses, the IRES has a common sequence motif consisting of a pyrimidine-rich tract followed by an AUG triplet located about 20 nt downstream (10, 18, 19). This motif is designated Yn-Xm-AUG (Yn, pyrimidine-rich tract; Xm, any nucleo-tides). Several lines of evidence indicate that internal initiation is facilitated by the interaction of IRES RNA with cellular proteins such as p52 (La protein) and p57 (polypyrimidine tract-binding protein [PTB]) (7, 14, 22, 25). Picornaviruses infect host cells and use cellular factors for internal initiation of viral RNA synthesis, which takes place in the cytoplasm. Classical initiation factors, such as eIF-4 and eIF-2, involved in cap-dependent translation initiation also bind to the picorna-virus IRES (9, 15, 24).
The HCV IRES has a Yn-Xm-AUG motif similar to that of picornaviruses. Mutational analysis of pyrimidine tracts, how-ever, revealed that the helical structure associated with the pyrimidine-rich tract is more important for the function of the HCV IRES than the primary sequence of the tract (28). Thus, precise definition of the functional RNA motif of the HCV
IRES is more problematic than precise definition of the cor-responding motif of the picornavirus IRES. Moreover, wheth-er cellular factors intwheth-eract with the HCV IRES and what pos-sible implications such interactions would have for initiation of translation remain unclear.
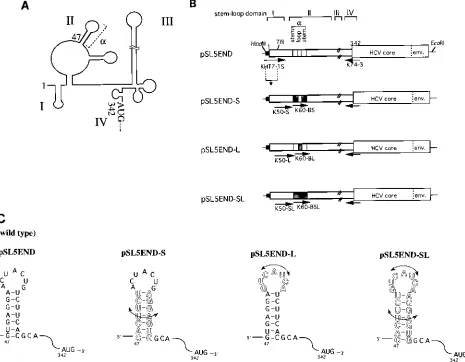
We previously reported that the translation efficiency pro-moted by an HCV 59NCR that lacked the 46 nt preceding the alpha-branch of stem-loop domain II (Fig. 1A) was compara-ble to that promoted by the wild-type 59NCR, whereas further truncation involving a beta-branch significantly reduced trans-lation efficiency (5). These data suggested that an alpha-branch structure in proximity to the 59functional region of the HCV IRES may contain a cis-acting element for translation initia-tion.
Characterization of the alpha-branch.In order to gain
fur-ther insight into the function of the alpha-branch, we construct-ed a series of alpha-branch mutants derivconstruct-ed from pSL5END, which had the full-length HCV 59NCR (nt 1 to 341) and the HCV core-envelope region (nt 342 to 968) as a reporter gene (5), by using PCR-mediated mutagenesis with the primers pre-sented in Fig. 1B. The sequences of the PCR primers used for cDNA construction are presented in Table 1. Plasmid pSL5END contained a wild-type sequence of the alpha-branch; this plas-mid resulted when cloned 5END cDNA (5) was subcloned, by PCR with primers KHT7-1 and K74-3 (5), into a pSL1180 vec-tor (Pharmacia) under the control of T7 promoter and with a
HindIII restriction site at its 59terminus. Plasmid pSL5END-S contained an exchange sequence within the alpha-branch stem region. Plasmid pSL5END-L contained an inverted loop se-quence of the alpha-branch. Plasmid pSL5END-SL contained an inverted alpha-branch sequence. The K60 primers that con-tained alpha-branch mutations were used to amplify mutant HCV 59 NCRs, followed by additional amplification of the HCV 59 NCR with K50 primers in combination with anti-sense K74-3 primer. The HindIII-TfiI fragment of wild-type * Corresponding author. Mailing address: Basic Research Division,
BioMedical Laboratories, Inc., 1361-1, Matoba Kawagoe-shi, Saitama 350-11, Japan. Phone: 81-492-32-0440. Fax: 81-492-32-5480.
1662
on November 9, 2019 by guest
http://jvi.asm.org/
pSL5END containing the terminal region of the wild-type 59 NCR and the T7 promoter sequence was ligated to these mutants to construct mutants with a full-length 59NCR. These mutants were constructed so that the secondary structure of the alpha-branch could be maintained (Fig. 1C). RNAs tran-scribed in vitro with T7 RNA polymerase from EcoRI-digested plasmid DNAs were translated in RRL and S10 cytoplasmic extracts prepared from suspension-cultured HeLa S3 cells (23). Translation reactions were performed with the optimum
[image:2.612.75.540.73.435.2]KCl concentration (72 mM) for HCV IRES function (5). The translation efficiencies of the RNA transcripts are shown in Fig. 2. Mutations in the loop sequence of the alpha-branch (5END-L) had little effect on translation efficiency in RRL extract (99.27%, compared with 100% for the wild-type se-quence, 5END-C) (Fig. 2, upper panel). Interestingly, the trans-lation efficiency was drastically reduced when the stem regions of the alpha-branch were exchanged (5END-S; 47.25%). When the alpha-branch was totally inverted, translation was virtually FIG. 1. (A) Schematic map of the predicted RNA secondary structure of HCV 59NCR RNA (2). The major stem-loop domains, I through IV, and the alpha-branch structure are represented. (B) Schematic representation of constructs containing alpha-branch mutations used to test for translation initiation of HCV. Primers used to amplify cDNA constructs are indicated by arrows. The stem and alpha-branch loop region mutations of the constructs are shaded. Black bars represents the T7 promoter sequence. Numbers indicate the nucleotide position in the HCV 5END cDNA sequence (5). env., envelope. (C) Sequence and structure models for alpha-branches containing stem and/or loop mutations. Outlined symbols depict exchanged nucleotides relative to the wild-type sequence.
TABLE 1. Primers for cDNA construction
Primer Sequence (59339)a
KHT7-1S...AAGGTACCAAGCTTTAATACGACTCACTATAGCCAGCCCCCTGATGGGGGCGACACT K60-BS...TAATACGACTCACTATACACTTCTACTACTGAGGAGTGGCAGAAAGCGTCTAGCCATGG K60-BL ...TAATACGACTCACTATAGTGAGGAGTCATCTTCACGCAGAAAGC
K60-BSL ...TAATACGACTCACTATACACTTCTGTCATCAAGGAGTGGCAGAAAGCGTCTAGCCATGG K50-S...CATGAATCACTCCCCTCACTTCTACTACTGAGGA
K50-L ...CATGAATCACTCCCCTGTGAGGAGTCATCATCTT K50-SL ...CATGAATCACTCCCCTCACTTCTGTCATCAAGGA
aT7 promoter sequence is underlined.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.58.561.637.717.2]abrogated (5END-SL; 22.78%). Inhibitory effects of mutations on translation efficiency were much clearer when these mu-tants were translated in a HeLa S10 lysate (Fig. 2, middle panel). Northern blot analysis indicated that the added wild-type and mutant RNAs were equally stable during translation reactions in HeLa S10 lysate (Fig. 2, lower panel). These re-sults demonstrated that the primary sequence of the stem region within the alpha-branch is an important element of the HCV IRES. The reduced translation efficiency of the mutants may have resulted from a loss of the ability of the 59NCR to bind to cellular factors that catalyze internal ribosome entry. We speculate that the stem region of the alpha-branch might be a cis-acting element that could participate in such interac-tions, although we could not eliminate the possibility that the introduced mutations could lead to substantial changes in the intact structure of the 59NCR.
Binding of cell proteins to 5*NCRs containing alpha-branch
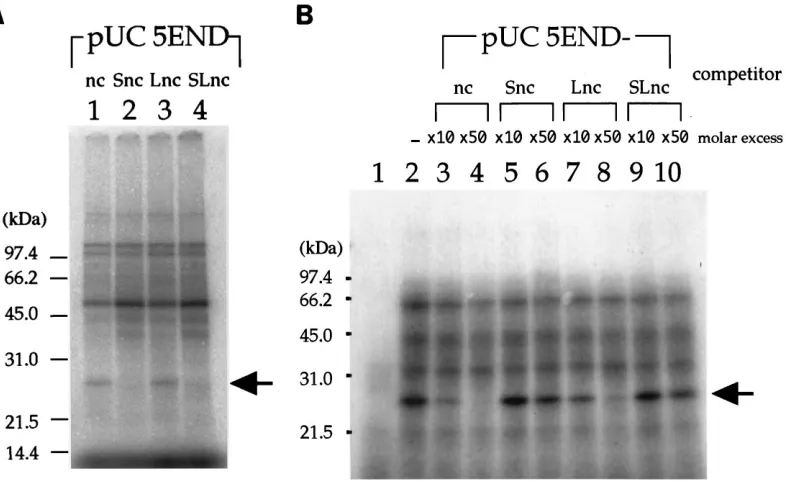
mutations. In order to look for cellular factors required for
translation initiation of HCV RNA, we first incubated HeLa S10 lysate with excess amounts of the RNA transcripts of pUC5END-nc, -Snc, -Lnc, and -SLnc that contained wild-type or mutant, full-length 59NCR fragments (nt 1 to 347) but did not contain a reporter gene (Fig. 3A) and subsequently added the wild-type HCV RNA for translation. Surprisingly, all four RNAs inhibited translation of the reporter RNA, although inhibition by pUC5END-nc and -Lnc was slightly stronger than that by pUC5END-Snc and -SLnc (Fig. 3B). Inhibition was not observed when unrelated RNA was preincubated with HeLa proteins (Fig. 3C, lanes 6 to 8). These results suggested that the HCV 59NCR maintained an affinity with several cellular fac-tors, which probably are involved in initiation of translation of HCV regardless of the presence of mutations. Translation ef-ficiency, however, dramatically decreased when the stem se-quence of the alpha-branch was changed (Fig. 2), suggesting the presence of another critical factor(s) that specifically inter-FIG. 2. Effects of alpha-branch mutations on translation promoted by the
HCV 59NCR. Lanes: (2), no template RNA; C, wild-type 59NCR as a positive control; S, mutations in the stem sequence; L, mutations in the loop sequence; SL, substitution of both stem and loop sequence. One microgram (for RRL) or 0.5 mg (for HeLa S10) of each RNA per 12.5ml of reaction mixture were translated as previously described (5). Translated products were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and the resulting autoradiograph is shown. The translation activities of mutated RNAs calculated by the MAC BAS program (Fuji) are also shown as intensities of the radioactivity of translation products normalized to the translation activity of pSL5END (lane C, 100%). RNA templates in the HeLa S10 reaction mixture were analyzed for integrity by Northern hybridization with [32
[image:3.612.65.290.67.267.2]P]UTP-labeled K74-3 oligonucleotide (lower panel).
FIG. 3. Inhibition of HCV translation by HCV 59NCR fragments containing alpha-branch mutations. (A) Schematic diagram of HCV 59NCR constructs used to test for inhibition of HCV translation. The alpha-branch and T7 promoter region are represented as in Fig. 1A. Plasmids were transcribed with T7 RNA polymerase after linearization with ScaI. (B) Translation of HCV RNA in the presence of full-length 59NCR fragments (nt 1 to 347) containing alpha-branch mutations. Lanes: 1, no template RNA; 2, no competitor RNA; 3 and 4, 5 and 6, 7 and 8, and 9 and 10, increasing amounts of competitor RNA transcribed from pUC5END-nc, -Snc, -Lnc, and -SLnc, respectively. Competitor RNAs were incubated with HeLa S10 lysate before the template RNA (0.5mg/12.5ml of reaction mixture) transcribed from pSL5END was translated. (C) Translation of HCV RNA in the presence of alpha-branch or unrelated RNA. Lanes: 1, no template RNA; 2, no competitor RNA; 3 to 5, alpha-branch RNA synthesized by custom services (Takara); 6 to 8, increasing amounts of unrelated RNA tran-scribed from HindIII-digested plasmid pMUG5-2, a cDNA clone of the 65 nt of the mumps virus leader region constructed as pMV393 (13).
1664 NOTES J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
acts with the HCV 59NCR, including the primary sequence of the stem region of the alpha-branch.
Cellular initiation factors, such as eIF-2 and eIF-4, present in cap-dependent systems are known to be involved in the cap-independent system promoted by the IRES of picornavirus (9, 15, 24). It is possible that these cellular factors are also involved in HCV IRES function, since the translation of the luciferase gene, which has no IRES structure on its 59 NCR, was inhibited by the addition of RNA transcripts from the HCV 59NCR (data not shown). Therefore, the observed re-sults of translation inhibition assays may have been due to the absorption of initiation factors by competitor RNA.
We were not able to obtain direct evidence that the stem region of the alpha-branch was specifically recognized by cel-lular factors. An inhibition of HCV translation was not ob-served when RNA transcripts of the alpha-branch were added as competitors (Fig. 3C, lanes 3 to 5). This result indicated that the alpha-branch by itself may not be sufficient for binding of cellular factors. In a cap-dependent system, several initiation factors appear to bind in a specific sequence in order for translation to occur (17, 21). The IRES of HCV may require sequential binding of factors, and the alpha-branch alone may not be sufficient for protein binding.
We next performed UV-induced cross-linking analyses to visualize proteins that bind to32P-labeled 59 NCRs. We used
full-length 59 NCRs transcribed from pUC5END-nc, -Snc, -Lnc, and -SLnc (Fig. 3A) as RNA probes. In order to find cellular proteins that bound to the HCV 59NCR during trans-lation initiation, we first incubated RNA probes with HeLa proteins under the salt conditions that were optimal for HCV RNA translation experiments. Protein-RNA complexes were attached covalently with a UV cross-linker (Stratagene) on ice
[image:4.612.115.508.70.311.2]for 15 min followed by incubation with 10 mg of RNase A. Labeled proteins were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and autoradiographed. As shown in Fig. 4A, several proteins from the HeLa S10 lysate cross-linked to all the RNA probes. These proteins may con-tain cellular factors involved in initiation of HCV RNA trans-lation that bind to the 59 NCR regardless of the presence of alpha-branch mutations. Cross-linking of the 25-kDa protein to the pUC5END-nc and -Lnc probes was detected (Fig. 4A, lanes 1 and 3). We increased the KCl concentration (120 mM) in the binding reaction mixture to obtain maximum binding of the 25-kDa protein to pUC5END-nc. Because translation of HCV RNA in 120 mM KCl retained about 80% of the activity obtained under optimized translation conditions (data not shown), we believe that cross-linking of the 25-kDa protein was reflected in IRES function. Moreover, in order to eliminate any nonspecific RNA-binding proteins, we added abundant nonspecific competitor RNAs [160 mg each of poly(I) and poly(I)zpoly(C) per ml] to the reaction mix. We were able to identify the 25-kDa protein more clearly (Fig. 4B, lane 2), whereas the intensities of other proteins were reduced. These results indicated that the 25-kDa protein bound specifically to the HCV 59 NCR. Competitive UV cross-linking assays showed that the binding of the 25-kDa protein to the HCV 59 NCR was greatly inhibited by pUC5END-nc and -Lnc (Fig. 4B, lanes 3, 4, 7, and 8). On the other hand, pUC5END-Snc and -SLnc had little inhibitory effect (Fig. 4B, lanes 5, 6, 9, and 10). These results revealed that the 25-kDa protein specifically bound to pUC5END-nc and -Lnc, both of which had a wild-type sequence in the stem region of the alpha-branch. It seems quite possible that this 25-kDa protein plays a critical role in HCV RNA translation, because the 59NCR RNA of FIG. 4. UV-induced cross-linking analysis of HeLa S10 proteins that bind to the HCV 59NCR. (A) Complex formation was carried out under the same salt conditions as those for HCV RNA translation. Portions (0.05 pg each) of [32
P]UTP-labeled full-length 59NCR fragments (nt 1 to 347) transcribed from ScaI-digested pUC5END-nc (lane 1), -Snc (lane 2), -Lnc (lane 3), and -SLnc (lane 4) were incubated with 10mg of HeLa S10 proteins for 10 min at 308C in a buffer containing 5.2 mM HEPES (pH 7.5), 72 mM KCl, 1.0 mM MgCl2, 0.78 mM magnesium acetate, 0.52 mM dithiothreitol, 3.8% glycerol, 0.75 mM ATP, and 1.0 mM GTP. (B) Competition by HCV 59NCR fragments for cross-linking of the 25-kDa protein. Complex formation was carried out under modified conditions [120 mM KCl and in the presence of 160mg each of poly(I) and poly(I)zpoly (C) per ml]. Lanes: 1, no protein; 2, no competitor HCV RNA; 3 and 4, 5 and 6, 7 and 8, and 9 and 10, increasing amounts of competitor RNA transcribed from pUC5END-nc, -Snc, -Lnc, and -SLnc, respectively. Competitor RNAs were incubated with HeLa S10 proteins before the addition of [32
P]UTP-labeled probe RNA transcribed from pUC5END-nc. The 25-kDa protein is indicated by an arrow.
on November 9, 2019 by guest
http://jvi.asm.org/
pUC5END-nc and -Lnc had efficient IRES function in the translation experiment (Fig. 2).
We suggest that the 25-kDa protein is different from PTB, which is reported to interact with IRES elements and to par-ticipate in initiation of translation of picornaviruses (7, 22), because the alpha-branch of the HCV 59NCR does not con-tain a pyrimidine-rich sequence. At present, the role of PTB in HCV IRES function remains unclear. The purified PTB was not enough to restore HCV IRES function when it was added to PTB-immunodepleted translation reaction mixtures, sug-gesting the importance of other proteins that associate with PTB (1). Furthermore, Kaminski et al. (11) reported that, in the case of the picornavirus IRES, PTB functioned as a pro-moter of correct folding of the IRES rather than as a central catalyst of internal ribosome entry. They also showed that the recombinant PTB did not bind to the HCV 59NCR and did not stimulate the function of the HCV IRES (11). The molecular mass of the 52- to 57-kDa proteins cross-linked to the HCV 59 NCR corresponded to the size of PTB (Fig. 4); therefore, we are now examining whether these proteins include PTB.
A requirement of the HCV core coding sequence immedi-ately downstream of an authentic AUG codon for IRES func-tion was recently reported (8, 20). Translafunc-tion competifunc-tion assays or UV cross-linking experiments should reveal the in-fluence of the core coding region on binding affinity for cellular proteins. Further characterization of cellular proteins, such as the 25-kDa protein, may help in further understanding the HCV translation mechanism.
We thank Akio Yamada for helpful discussions.
This research was supported by grants from the Viral Hepatitis Research Foundation of Japan.
REFERENCES
1. Ali, N., and A. Siddiqui. 1995. Interaction of polypyrimidine tract-binding protein with the 59noncoding region of the hepatitis C virus RNA genome and its functional requirement in internal initiation of translation. J. Virol.
69:6367–6375.
2. Brown, E. A., H. Zhang, L. H. Ping, and S. M. Lemon. 1992. Secondary structure of the 59nontranslated regions of hepatitis C virus and pestivirus genomic RNAs. Nucleic Acids Res. 20:5041–5045.
3. Bukh, J., R. H. Purcell, and R. H. Miller. 1992. Sequence analysis of the 59 noncoding region of hepatitis C virus. Proc. Natl. Acad. Sci. USA 89:4942– 4946.
4. Cha, T.-A., J. Kolberg, B. Irvine, M. Stempien, E. Beall, M. Yano, Q.-L.
Choo, M. Houghton, G. Kuo, J. H. Han, and M. S. Urdea.1991. Use of a signature nucleotide sequence of hepatitis C virus for detection of viral RNA in human serum and plasma. J. Clin. Microbiol. 29:2528–2534.
5. Fukushi, S., K. Katayama, C. Kurihara, N. Ishiyama, F. B. Hoshino, T.
Ando, and A. Oya.1994. Complete 59noncoding region is necessary for the efficient internal initiation of hepatitis C virus RNA. Biochem. Biophys. Res. Commun. 199:425–432.
6. Han, J. H., V. Shyamala, K. H. Richman, M. J. Brauer, B. Irvine, M. S.
Urdea, P. Tekamp-Olson, G. Kuo, Q. L. Choo, and M. Houghton.1991. Characterization of the terminal regions of hepatitis C viral RNA: identifi-cation of conserved sequences in the 59untranslated region and poly(A) tails at the 39end. Proc. Natl. Acad. Sci. USA 88:1711–1715.
7. Hellen, C. U., G. W. Witherell, M. Schmid, S. H. Shin, T. V. Pestova, A. Gil,
and E. Wimmer.1993. A cytoplasmic 57-kDa protein that is required for translation of picornavirus RNA by internal ribosomal entry is identical to
the nuclear pyrimidine tract-binding protein. Proc. Natl. Acad. Sci. USA
90:7642–7646.
8. Honda, M., L.-H. Ping, R. C. A. Rijnbrand, E. Amphlett, B. Clarke, D.
Rowlands, and S. M. Lemon.1996. Structural requirements for initiation of translation by internal ribosome entry within genome-length hepatitis C virus RNA. Virology 222:31–42.
9. Jackson, R. J., S. L. Hunt, J. E. Reynolds, and A. Kaminski. 1995. Cap-dependent and cap-inCap-dependent translation: operational distinctions and mechanistic interpretations. Curr. Top. Microbiol. Immunol. 203:1–29. 10. Jang, S. K., and E. Wimmer. 1990. Cap-independent translation of
encepha-lomyocarditis virus RNA: structural elements of the internal ribosomal entry site and involvement of a cellular 57-kD RNA-binding protein. Genes Dev.
4:1560–1572.
11. Kaminski, A., S. L. Hunt, J. G. Patton, and R. J. Jackson. 1995. Direct evidence that polypyrimidine tract binding protein (PTB) is essential for internal initiation of encephalomyocarditis virus RNA. RNA 1:924–938. 12. Kato, N., M. Hijikata, Y. Ootsuyama, M. Nakagawa, S. Ohkoshi, T.
Sug-imura, and K. Shimotohno.1990. Molecular cloning of the human hepatitis C virus genome from Japanese patients with non-A, non-B hepatitis. Proc. Natl. Acad. Sci. USA 87:9524–9528.
13. Leopardi, R., V. Hukkanen, R. Vainionpa¨a¨, and A. A. Salmi.1993. Cell proteins bind to sites within the 39noncoding region and the positive-strand leader sequence of measles virus RNA. J. Virol. 67:785–790.
14. Meerovitch, K., Y. V. Svitkin, H. S. Lee, F. Lejbkowicz, D. J. Kenan, E. K. L.
Chan, V. I. Agol, J. D. Keene, and N. Sonenberg.1993. La autoantigen enhances and corrects aberrant translation of poliovirus RNA in reticulocyte lysate. J. Virol. 67:3798–3807.
15. Meyer, K., A. Petersen, M. Niepmann, and E. Beck. 1995. Interaction of eukaryotic initiation factor eIF-4B with a picornavirus internal translation initiation site. J. Virol. 69:2819–2824.
16. Okamoto, H., S. Okada, Y. Sugiyama, S. Yotsumoto, T. Tanaka, H.
Yo-shizawa, F. Tsuda, Y. Miyakawa, and M. Mayumi.1990. The 59-terminal sequence of the hepatitis C virus genome. Jpn. J. Exp. Med. 60:167–177. 17. Pause, A., N. Methot, Y. Svitkin, W. C. Merrick, and N. Sonenberg. 1994.
Dominant negative mutants of mammalian translation initiation factor eIF-4A define a critical role for eIF-4F in cap-dependent and cap-indepen-dent initiation of translation. EMBO J. 13:1205–1215.
18. Pilipenko, E. V., A. P. Gmyl, S. V. Maslova, Y. V. Svitkin, A. N. Sinyakov, and
V. I. Agol.1992. Prokaryotic-like cis elements in the cap-independent inter-nal initiation of translation on picornavirus RNA. Cell 68:119–131. 19. Pilipenko, E. V., A. P. Gmyl, S. V. Maslova, G. A. Belov, A. N. Sinyakov, M.
Huang, T. D. Brown, and V. I. Agol.1994. Starting window, a distinct element in the cap-independent internal initiation of translation on picornaviral RNA. J. Mol. Biol. 241:398–414.
20. Reynolds, J. E., A. Kaminski, H. J. Kettinen, K. Grace, B. E. Clarke, A. R.
Carroll, D. J. Rowlands, and R. J. Jackson.1995. Unique features of internal initiation of hepatitis C virus RNA translation. EMBO J. 14:6010–6020. 21. Rhoads, R. E. 1993. Regulation of eukaryotic protein synthesis by initiation
factors. J. Biol. Chem. 268:3017–3020.
22. Rojas-Eisenring, I. A., M. Cajero-Juarez, and R. M. del Angel. 1995. Cell proteins bind to a linear polypyrimidine-rich sequence within the 59 -untrans-lated region of rhinovirus 14 RNA. J. Virol. 69:6819–6824.
23. Rose, J. K., H. Trachsel, K. Leong, and D. Baltimore. 1978. Inhibition of translation by poliovirus: inactivation of a specific initiation factor. Proc. Natl. Acad. Sci. USA 75:2732–2736.
24. Scheper, G. C., A. A. Thomas, and H. O. Voorma. 1991. The 59untranslated region of encephalomyocarditis virus contains a sequence for very efficient binding of eukaryotic initiation factor eIF- 2/2B. Biochim. Biophys. Acta
1089:220–226.
25. Schmid, M., and E. Wimmer. 1994. IRES-controlled protein synthesis and genome replication of poliovirus. Arch. Virol. Suppl. 9:279–289. 26. Tsukiyama-Kohara, K., N. Iizuka, M. Kohara, and A. Nomoto. 1992.
Inter-nal ribosome entry site within hepatitis C virus RNA. J. Virol. 66:1476–1483. 27. Wang, C., P. Sarnow, and A. Siddiqui. 1993. Translation of human hepatitis C virus RNA in cultured cells is mediated by an internal ribosome-binding mechanism. J. Virol. 67:3338–3344.
28. Wang, C., P. Sarnow, and A. Siddiqui. 1994. A conserved helical element is essential for internal initiation of translation of hepatitis C virus RNA. J. Vi-rol. 68:7301–7307.
1666 NOTES J. VIROL.