0022-538X/79/08-0370/06$02.00/0
Host
Cell
DNA
Chain
Initiation
Protein
Requirements for
Replication
of
Bacteriophage
G4
Replicative-Form
DNA
LAWRENCE B. DUMAS,* SHERRY GOLTZ, AND DANA T. BENESH
Departmentof Biochemistryand MolecularBiology,NorthwesternUniversity, Evanston,Illinois60201
Received for publication 27 February 1979
Bacteriophages G4evl and G4bslaresimple temperature-resistant derivatives ofwild-typeG4 as demonstratedbyrestrictionendonuclease analyses. Therate ofreplicationof theduplex replicative-formDNAof thesephageswasnormal in dnaBand dnaC mutants of thehost,whereas the rate wasmarkedly reduced in adnaG hostmutant atthe restrictivetemperature.We concludethatG4 duplex DNAreplicationrequires the host cell dnaG protein, but not the dnaB anddnaC proteins. Thereasonsfor the differences betweenourconclusionsand those based onpreviously publisheddata aredocumented anddiscussed.
TheEscherichia colidnaGproteincatalyzes
the synthesis of an oligonucleotide primer on
bacteriophage G4 DNA in vitro without the assistance of the dnaB and dnaC initiation pro-teins (1, 14, 17). This also seems to be true in thecell,where theinfecting single-strandedG4 DNA molecule serves as template for the syn-thesis ofthecomplementarystrand (11).
Invivo studies of the host cell initiation pro-teinrequirements for replicationonthe duplex
G4replicative-form (RF)DNAarecomplicated
by the inherent temperature sensitivity of G4 RF DNA replication. It is desirable to use a
temperature-resistant strain of G4 when
study-ing the effects of temperature-sensitive host DNA synthesismutations onG4 RF DNA rep-lication. To this end, the host cell requirements for duplex RF DNA replication ofa tempera-ture-resistant bacteriophage, designated G4trl,
have been shown to include dnaB and dnaC proteins, aswellasthe dnaGprotein (3).These experiments,togetherwith thoseof Kodaira and Taketo using wild-type G4 (12), have been in-terpreted as demonstrating that bacteriophage G4 requires the hostcell dnaB and dnaC pro-teins, together with the dnaG protein, for the replicationof itsduplexRF DNA.
Recent experiments in ourlaboratory ledus to question the assignation of bacteriophage
G4trlas asimplevariantofwild-typeG4. There-fore,wecomparedthe structures of the RF DNA molecules isolated from cells infected with G4trl,wild-type G4,andtwoother temperature-resistant strains ofG4. We found G4trl to be differentfrom G4.Thus,G4trlisnot an appro-priate probe for determining the requirements
for G4 RF DNAreplication.
We attempted to use wild-type G4 to deter-mine therequirementsfor hostcell DNA chain
initiation proteins. However, theinherent tem-perature sensitivity of wild-type G4 made the interpretation of several key experiments
ex-tremelydifficult. In view ofthis,wedecided that wild-type G4 also should not be used toprobe thisevent. This prompted us toreexamine the replication of G4 duplex RF DNA in host cell DNA synthesis-defective mutants, using
tem-perature-resistantstrains ofG4.
MATERIALS AND METHODS
Bacteria andphagestrains. E. coli strains LD311 [uvrA- thyA- endI dnaB(Ts)] and LD332
[uvrA-thyA- endI- dnaC(Ts)] are temperature-sensitive mutantsof H502 (uvrA-thyA-endI-) described
pre-viously (5, 6). E. coli strain C2309 [uvrA- thyA(Ts)
dnaG(Ts)]wasobtainedfrom R.Calendar.
The bacteriophage strains used in these
experi-ments arelistedin Table 1,asarethe sources of the
phage and some of their distinguishing properties. Media and buffers. TPGA medium is TPG me-dium (15) with1 gofKH2PO4 and 10 g ofCasamino Acids perliter. SVB medium has been described (2). Buffer E consists of40mM Tris-acetate (pH 7.8), 5 mMsodium acetate, and1mM EDTA(16).
IsolationofphageRF DNA.Intracellular double-stranded RF DNAwasisolated from chloramphenicol-treated, phage-infected E. colicelLsby two different procedures. Host chromosomal DNA was removed fromlysates of infected cells by the procedure of either Godson and Vapnek (9) or Hansen and Olsen (10). Thephage RF DNAwasfurtherpurified using phenol extraction, precipitation with ethanol, RNase diges-tion of contaminating RNA, and sedimentation through isokinetic sucrosegradients. DialyzedDNA
sampleswerestored in 50mM Tris-hydrochloride-2 mMEDTA, pH 7.5,at-80°C.
Restriction endonuclease fragment analyses. The restrictionendonucleases HaeIII,HhaI, HincII, HpaII, and PstIwerepurchased from Bethesda Re-search Laboratories. Endonuclease EcoRI was pur-chased fromMiles Laboratories.Reactions withDNA 370
on November 10, 2019 by guest
http://jvi.asm.org/
PROTEIN REQUIREMENTS FOR G4 RF DNA REPLICATION 371
TABLE 1. Sourcesandpropertiesof bacteriophages
Phage Property Source
G4 Wildtype G. N.Godson
G4trl Temperatureresistant G. N. Godson
G4bsl Temperatureresistant R. C.Warner
G4evl Temperature resistant Thislaboratory
OX174am3 Lysisdefective R.L.Sinsheimer
*Kh-1 Hostrange mutant A.Taketo
werecarriedoutessentiallyasspecified by these
sup-pliers. The reactions (total volume,50p1)were
termi-nated after 1hby the addition of 10u1ofasolution
containing 2 Msucrose, 100mMEDTA(pH 8), and 0.04% (wt/vol)bromophenol blue.
The DNAfragmentswereseparated by
electropho-resison1%(wt/vol)agaroseslabgels containing buffer Eand 1 pgofethidium bromideperml,asdescribed
by Sugdenetal. (16). The running buffer (buffer E) included0.5 pgof ethidium bromideperml.
Electro-phoresiswascarriedout at 100V forapproximately 2.5h.Thegelswerephotographedover ashortwave
UVlight box,using Polaroidtype47film.
DNA synthesi8 rate measurements. Host bac-teriaweregrownat330ConTPGAmedium (80 ml)
supplemented with 2 pug of thymine per ml to cell
densities ofapproximately 3 x 105 cellsperml. The
cellswerecollectedbycentrifugation andsuspended
in 5 mlof SVB medium supplemented with10 mM
CaCl2. Phagewerethenadded, G4atamultiplicity of
infection of 10or 4X174am3atamultiplicity of
infec-tion of 3. After20min of incubationat330C without aeration, 75mlofTPGA mediumsupplemented with
1 pgof thymine and 30 pug ofchloramphenicolperml wasadded (zero time). Incubation wascontinuedat
330C for 120 min. At 20 and 40 min, portions of the culturewereshiftedtothe restrictivetemperature.
At 10-min intervals, 20 ml of each culture was
transferred to separate vessels containing 10 piCi of
[3H]thymidinein0.5 ml ofTPGA mediumand incu-bated for 2min either at33°C oratthe restrictive
temperature.Coldacetonewasaddedtoterminate the
pulse. Thecellswerecollectedby centrifugation and suspended in 1 ml of 50 mMsodium tetraborate-10 mMEDTA, pH 9. Lysozyme (200 pg)wasadded,and
themixtureswereincubated for15minatroom
tem-perature. Potassiumhydroxide wasthen addedto a
final concentration of0.3 N, and the lysates were
incubatedovernightat30°C. Calf thymus DNA carrier (200 pg) and 2 ml of cold 10%trichloroacetic acidwere
added. Theprecipitates werecollectedonglass fiber
filters.Theamountofacid-insoluble radioactivity in
eachsamplewasthenmeasured withaliquid
scintil-lationspectrometer.
RESULTS
Twopreliminary observationsraised the
pos-sibilitythatG4trl andwild-type G4were quite
differentphages. (See Table 1foradescription
of these and the other phages used in these experiments.) DNA synthesis on G4trl
single-strandedtemplateDNAwasdefectiveinsoluble
protein extracts of the E. coli dnaC defective
mutantPC79, whereas DNAsynthesison wild-typeG4DNAwas not(M. Bayneand L.Dumas, unpublished data). Second, G4bsl formed plaques with high efficiency at 41°C on the dnaB defective mutant host strain LD311, whereas G4trl did not (D. Benesh and L. Du-mas,unpublished data).Given thesedifferences,
we decided to determine whether G4trl is, in fact, asimple derivative ofG4 oranother bac-teriophage.
Restriction endonucleasefragment anal-yses. DuplexRF DNAmoleculeswereisolated from phage-infected cells. The analyses of the products of digestion with endonuclease PstI
showed that G4, G4trl, G4bsl, and G4evl RF DNA molecules all carryasingle cleavagesite recognized by this enzyme. Endonuclease EcoRI
also recognized a single cleavage site on G4, G4bsl,and G4evl RFDNA,butnositeonG4tr1 DNA. These data (not shown) suggest that
G4trl RF DNA is different from that ofG4, G4bsl,andG4evl.
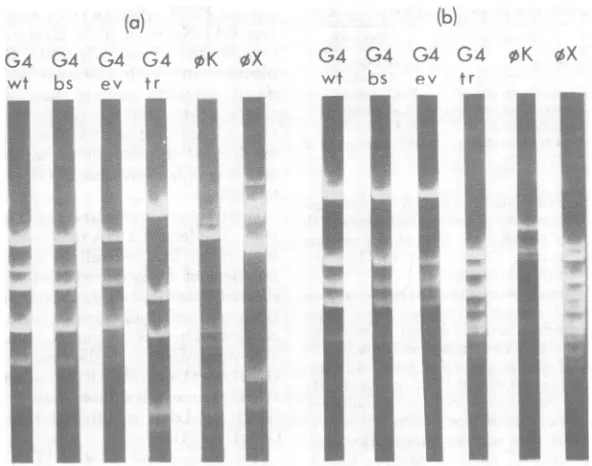
The uniqueness ofG4trl DNA is even more apparentfrom the data shown inFig. 1. Diges-tion of G4, G4bsl, and G4evl RF DNA with endonuclease HpaII yielded DNA fragments
withidentical sizes uponseparation by
electro-phoresis in agrose gels (Fig. la). The number and sizes of fragments generated from G4trl
DNAweredifferent from those of the other G4
phages and from 4Kh-1 and 4X174. Similarly, Fig. lb shows that digestion ofG4, G4bsl, and
G4evl RF DNA molecules with endonuclease
HincIIyieldedidentical
products.
Thefragment
pattem ofG4trl DNAagain differed from the other G4phages, from4Kh-1 , and fromfX174.
It isclear,therefore,that G4trlisunlikethese otherphages. Agarosegel electrophoresisof
en-donuclease HaelII and HhaI digests yielded
data consistent with thisconclusion.
Hostproteins
required
for G4 RF DNAreplication.
Since G4trl isnot asimple
deriv-ative ofG4,previously published
datausing
thisphage (3) cannotbe taken asindicative of host cell DNAsynthesisprotein requirementsfor G4 RF DNAreplication. Inaddition,the data ob-tainedusingwild-type G4 (12)are notsufficient forestablishingthesehostreqirementsbecause of the inherent temperature
sensitivity
of the phage. We found that the inhibition of G4 RF DNA replicationobserved, for example,inthe dnaCmutanthostatthe restrictivetemperature (12)wasalso evident intemperature-insensitiverevertants of the mutant host
(unpublished
data). Thus,datafromtheseexperimentscannot
be usedtoargue that the host cell dnaC
protein
isrequiredfor G4 RFDNAreplication.
Rather,
experiments of this kind must be carried out withG4temperature-resistantphages.Wehave
VOL. 31,1979
on November 10, 2019 by guest
http://jvi.asm.org/
(ci
(b!
G4
GdG0
> KOX
G04
G40G
04
0K 0wt L) e I"t bs ev
tf-Pjl!~~~~~~~~~~~~~~~1W
E
*
S:*
X |
E
|
.-..~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.
.,....
|
FIG. 1. Agarose gelelectrophoresisoftherestriction endonucleasedigestionproducts of RF DNA Phage G4,G4bsl,G4evl, G4trl, pKh- 1,and4X174am3RFDNAmoleculeswerecleaved withHpaII (a)andHincII (b). Thedigestionproductswereseparated by electrophoresisasdescribed in thetext.
used G4evl and G4bsl inaseries of suchexper- G4bsl (data not shown) were consistent with
iments. thisconclusion.
We infected mutant and revertant host cell In similarexperiments, G4evl RF DNA rep-culturescontaining30,ugofchloramphenicolper licationrates weremeasuredatpermissive and ml withG4evl,shiftingportionsof the cultures restrictive temperatures in the dnaCdefective
totherestrictive temperatureat20and40min mutanthost LD332 andits
temperature-insen-afterinfection. The cells had been treated with sitiverevertant. Theresults of theseexperiments mitomycinC before infection toselectively in- areshown inFig.3.Again,shiftstotheelevated
hibit host DNAsynthesis.TheratesofG4 DNA temperaturedidnotresultinmorereduction of
replication were measuredat 10-min intervals. therate of RF DNA replicationin themutant
Essentially all of the radioactive DNA made hostthan in therevertant (compare Fig. 3Ato
under the conditions of thesepulse-labelexper- B).The fact that the dnaCproteinwasdefective iments in all host strains used was found in under theseconditionswasdocumented by the
duplexRF DNA(datanotshown). observation that 4X174 RF DNA replication
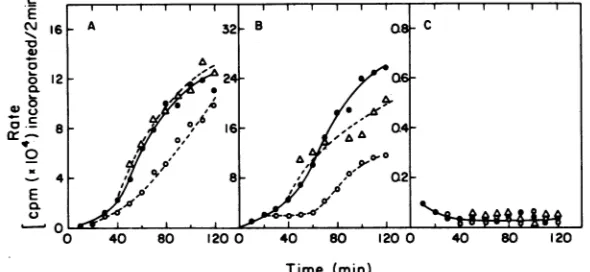
Figure2showsthe results ofmeasurementsof was extremely slow, even at 330C (Fig.
30).
ratesofG4evl RFDNAreplicationinthe dnaB Previous experiments (6, 13) have shown that mutanthost LD311.Shifts from the permissive the dnaCprotein activity is diminished inthis temperature to the restrictive temperature re- host even at the permissive temperature. The sulted in similar changes in rate of RF DNA amounts of labeled DNA synthesized in the replicationin the mutant host(Fig.2A)andthe 4X174-infected culture were near the
back-temperature-insensitiverevertanthost(Fig. 2B). ground levels seen in uninfected cell cultures The rate of 4X174 RF DNA replication was under these conditions and more than 20-fold markedly reducedupon shifting to
400C
at 20 lower than the amountssynthesized in4X174-and40 min after infection (Fig.
20),
indicating infectedrevertantcultures(datanotshown). thatthednaBproteinwasdefective attheele- Since G4 RF DNA replication was no morevated temperature. The observationthatG4evl temperature sensitive in themutant hostthan RF DNA replication was not inhibited under in the revertant, the dnaC protein does not conditionsinwhichthednaBproteinwasdefec- appear to be essential. The results of similar tive indicates that thisprotein is notessential experiments withG4bsl (datanotshown) were forreplication oftheduplexG4DNAmolecule. consistentwiththisconclusion.
The results of analogous experiments using The hostcell dnaGproteinisessentialforG4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.108.404.77.310.2]PROTEIN REQUIREMENTS FOR G4 RF DNA 373
c E
C\M
E.0
0 0
0 .
-o
cr c
0 E
CL,a
Time(min)
FIG. 2. RF DNA replicationratesinthe dnaBmutanthoststrain LD311. (A) G4evl -infected LD311; (B) G4evl-infected LD311R,aspontaneoustemperature-resistantrevertantof LD311; (C) XX174-infected LD311. Symbols: *,rates at33°C;0,rates at40°C, shifted from33to40°Cat20min;A,rates at40°C, shiftedat40 min.
E
CX
16 A 328 Co i2 -'24 Q6
a
0 40 80A120,016,P40 80 120 0Q 40 80 120
0~~~~~~~~~~,
0 40 80 120 0 40 80 1200 40 120
[image:4.504.106.404.60.223.2]Time (min)
FIG. 3. RF DNA replicationratesinthe dnaC mutant hoststrain LD332. (A) G4evl-infected LD332; (B) G4evl-infected LD332R,aspontaneoustemperature-resistantrevertantof LD332; (C) 4xX174-infected LD332. Thesymbolsarethesame asinFig. 2.
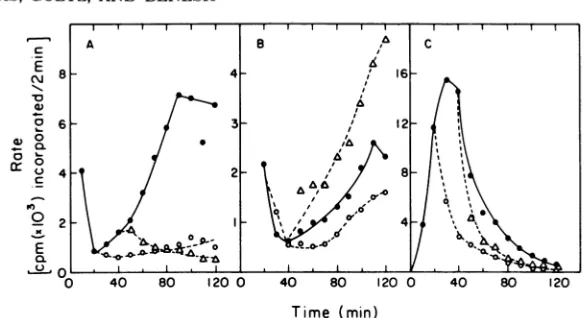
RF DNAreplication. Figure4showsthatG4evl RF DNA replicationrates were moresensitive to shifts to the restrictive temperature in the mutanthost(Fig. 4A)thanintherevertant(Fig. 4B).Thiscontrol demonstrates thatthe
require-ments for essential host cell proteins can be
clearly demonstrated withthesemeasurements.
DISCUSSION
Bacteriophages G4evl and G4bsl have been designatedasderivativesofwild-typeG4onthe
basisof theidenticalfragmentpatternsobtained
upondigestionwithendonucleasesEcoRI, PstI,
HincII, HpaIII, HaeIII, and HhaI. G4evl and G4bslarelessinherentlytemperaturesensitive
thanwild-typeG4.Measurements of theratesof G4evl RF DNA replication at the restrictive temperature in dnaB and dnaCmutantsofE. coli and their revertants show that RF DNA
replication continues atnormal rates. This in-dicates that theprotein products of these two genes are not essential for normal G4evl RF DNAreplication. Incontrast, RF DNA
replica-tionisinhibitedattherestrictivetemperaturein adnaG mutanthost. Theproduct of thishost geneisrequiredfornormal RF DNAreplication. These observations are consistent with those
madeusingG4bsl-infectedcells.
Ourconclusion that G4evl and G4bsl phages do not require the host cell dnaB and dnaC
proteinsfornormal RFDNAreplicationdiffers from theinterpretations of the data from pre-vious expenmentswithG4trl (3) and G4 wild-type (12). The restriction endonuclease digest data presentedhere demonstrate that G4trl is differentfromG4.Therefore,the factthat G4trl
requiresthe host cell dnaB and dnaCproteins forRF DNAreplicationis notinconsistentwith the observation that G4 does not. Rather, it
V
.40 80 120 0 40 80 1200 40 80 120
VOL. 31,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.105.400.278.414.2]C)
C\ "I
0.
E.0
C
9 2
E
Oa 0
B a
A
l
_ a
/
, 0 °~~~~~~0
_0 ' 'O
4
.
40 80 120 0 40 80 120 0 40 80 120
4
3
2
12
8
Time (min)
FIG. 4. RF DNAreplication rates in the dnaG mutant host strain C2309. (A) G4evl-infected C2309; (B)
G4evl-infectedC2309R, a spontaneous temperature-resistant revertant of C2309; (C)4X174-infected C2309. Thesymbols are the same as inFig.2.
supports ourcontention that G4trl isa misno-merforaphage that isquite different from G4. Also,ourdata fromwild-typeG4-infected host mutants (data not shown) do not differ from those published previously (12), but our inter-pretations do. We donotinterpretthe temper-aturesensitivityof RF DNAreplicationin dnaB and dnaC mutant hosts to be due to the dnaB anddnaCmutations, respectively,sincewe ob-served similar temperature sensitivity in
tem-perature-insensitiverevertantsof these host mu-tants. Instead, we argue that valid interpreta-tions require the use oftemperature-resistant
strainsof G4 and data from infections of revert-anthostsaswellas mutanthosts.
Thus,therequirementsforG4 RF DNA rep-lication are similar to those of other group II isometricphagessuchas
4K,
St-1,anda-3 (forreview, seereference4), in that the dnaG
pro-tein, but not the dnaB and dnaC proteins, is required for DNA chain initiation. GroupI iso-metric phages, suchas
4X174,
S13, and4A
(4),however, do require the dnaB and dnaC pro-teinstoassist thednaGproteininthisprocess. ThednaGprotein is abletocatalyzethe syn-thesis ofoligonucleotideprimersonthe freeviral strandsofG4 DNA in vitro and in vivo without the assistance of the dnaB and dnaC proteins (1,11, 14, 17). Thus, the host cell DNA initiation protein requirements for circular G4 RF DNA replication are consistent with the replication modelproposed by Fiddesetal. (7) and with the
simplest version oftherolling-circle model(8). Accordingtothesemodels,cleavage of the viral DNAstrand of the duplexRFmolecule atthe origin of replication provides a 3'-OH primer terminus forchainelongation, using the comple-mentary strand astemplate.No de novo chain initiationisrequired. Displacementof the viral
template strand from its 5' end eventually ex-poses the same de novo chain initiation site recognized by the dnaGprotein duringthe con-version of the free infecting viral strandto its duplex replicative form early in infection. De novo chain initiation catalyzed by the dnaG proteinoccurs atthissamesite during RF rep-lication without the need for the dnaB and dnaC proteins. G4 escapes the need for these latter two initiation proteins, as do other group II isometric phages, simply because the viral strand carries a site recognized by the dnaG protein. Asingledenovochaininitiation siteon the viral strand, therefore, is sufficient for the RF DNAreplicationprocess.
ACKNOWLEDGMENTS
Thisworkwassupported byPublic HealthServiceresearch
grant AI-09882 and researchcareerdevelopmentaward AI-70632toL.B.D.from the NationalInstituteofAllergyand Infectious Diseases. S.G. was supported in partby Public Health ServicetraininggrantGM-07291 from the National InstituteofGeneralMedicalSciences.
LITERATURE CITED
1. Bouche, J.-P.,K.Zechel,and A.Kornberg.1975.dnaG
geneproduct,arifampicin-resistantRNApolymerase,
initiates conversion of asingle-stranded coliphage DNA
toitsduplex replicativeform. J. Biol. Chem.
270:5995-6001.
2.Denhardt, D. T., and R. L. Sinsheimer. 1965. The
process of infection with bacteriophage *X174. III. Phage maturation andlysisaftersynchronized infec-tion. J.Mol. Biol. 12:641-646.
3. Derstine, P.L.,L. B.Dumas,andC. A.Miller.1976. Bacteriophage G4 DNAsynthesisin temperature-sen-sitive dnamutantsof Escherichia coli. J. Virol. 19: 915-924.
4. Dumas, L.B. 1978.Requirementsfor hostgeneproduct,
inreplicationofsingle-strandedphageDNAinvivo,
341-359.In D. T.Denhardt,D.Dressler,and D. S.Ray
(ed.),Single-strandedDNAphages.ColdSpringHarbor
Laboratory, Cold Spring Harbor, N.Y.
A
16
II II1. Ct
& 11
'CLA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.113.406.66.225.2]5. Dumas, L B., and C. A.Miller. 1974. Inhibition of bacteriophageOX174DNAreplication in dnaBmutants of Escherichia coli C. J.Virol. 14:1369-1379. 6. Dumas, LB., C. A. Miller, and M. L Bayne. 1975.
Rifampin inhibition of bacteriophageOX174parental replicative-form DNA synthesis inanEscherichiacoli dnaCmutant.J.Virol. 16:575-580.
7. Fiddes, J. C., B. G.Barrell,and G. N. Godson.1978. Nucleotidesequencesof theseparateorigins of synthe-sis of bacteriophage G4viral and complementary DNA strands. Proc.Natl. Acad. Sci. U.S.A. 76:1081-1085. 8. Gilbert, W., and D. Dressler. 1968. DNAreplication:
the rolling circle model. Cold Spring Harbor Symp. Quant. Biol. 33:473-484.
9. Godson, G. N., and D. Vapnek. 1973. A simple method forpreparinglargeamountsofOX174RFsupercoiled DNA. Biochim.Biophys. Acta 299:516-520.
10. Hansen, J. B., and R.H. Olsen. 1978. Isolation of large
bacterialplasmids and characterization of the P2 incom-patibilitygroupplasnids pMGI and mMG5. J. Bacte-riol. 135:227-238.
11. Kodaira, K-I., and A. Taketo. 1977. Conversion of bacteriophage G4 single-stranded viral DNAto
double-stranded replicative form in dna mutantsof Esche-richia coli. Biochim. Biophys. Acta 476:149-155. 12. Kodaira,K.-I.,and A.Taketo. 1978.Replication of G4
progenydouble-stranded DNA indnamutants
ofEsch-erichia coli. J. Biochem. (Tokyo)83:971-976. 13.Kranias, E. G., and L B. Dumas. 1974. Replication of
bacteriophagefX174 DNA inatemperature-sensitive dnaCmutantofEscherichia coli C. J.Virol. 13:146-154.
14.Rowen,L, and A. Kornberg. 1978. Primase, the dnaG protein of Escherichia coli. Anenzymewhich starts DNAchains. J. Biol.Chem. 253:758-764.
15.Sinsheimer, R. L, B. Starman, C. Nagler, and S. Guthrie.1962.Theprocessofinfection with bacterio-phageOX174.I.Evidenceforareplicativeform. J. Mol. Biol. 4:142-160.
16. Sugden, B., B. DeTroy, R. J. Roberts, and J. Sam-brook. 1975. Agarose slab-gel electrophoresis equip-ment.Anal. Biochem. 68:36-46.
17. Wickner,S. 1977. DNAorRNApriming of bacteriophage G4 DNAsynthesis by Eacherichia coli dnaG protein. Proc.Natl. Acad. Sci. U.S.A. 74:2815-2819.
on November 10, 2019 by guest
http://jvi.asm.org/