Vol.30,No. 1
JouRNAL OFVIROLOGY, Apr.1979, p. 1-13 0022-538X/79/04-0001/13$02.00/0
Analysis of
Coliphage
Lambda Mutations That Affect Q Gene
Activity: puq, byp, and nin5
NAT STERNBERGt* D LYNNENQUISTtLaboratory ofMolecular Genetics, National Institute of Child Health and Human Development, National
Institutesof Health,Bethesda, Maryland 20014 Receivedfor publication 13 September 1978
We describe in this paper the isolation and characterization of a class of
mutations, designatedpuq, thatallow phage X to grow better under conditions
that limit the synthesis of the phage Q gene product. These mutations were
locatedbetween phage genes P andQ, a region of the X chromosome containing twogeneN-independent mutations, nin5and
byp,
that we also show tobe puq mutations. Whereas the puq-3 and puq-16 mutations probably map under the nin5 deletion, the byp mutation maps between this deletion and the QX-Q<80crossoverpoint.These mutationslikelyactbyincreasing thesynthesis of the Q
gene product. We demonstrate that the clear-plaque phenotype and reduced
lysogenization frequency of byp mutants depend on increased Q gene activity. The significance of these results in understanding how transcription proceeds
throughtheP-Q region of the A genome is discussed.
When coliphage A infects Escherichia coli,
oneoftwomodesofphagedevelopmentensues. In thelysogenic mode, the phage coexists with
thehost withoutkilling it.Inthelytic mode, the phage rapidly reproduces and, in the process,
kills the host. The lysogenic response is the result ofrepressionof mostphage functions by
the phage cI gene product. The faithful
segre-gationofrepressed phageDNA(prophage)
dur-ing cell division is ensured by integration of
prophage DNA into the hostgenome, whereit isreplicated passively bythe host. In contrast,
during the lytic response, the phage DNA is actively replicatedand transcribed, phage par-ticlesareassembled,andcell lysisoccurs.Inthis
report, weanalyze the action of several phage functions involvedinlytic growth.
Regulation of transcription during the lytic growth cycle dependsonthe actionof the prod-uctsofphagegenes Nand
Q (16).
Inthe absence of theN geneproduct,
transcription
thatiniti-atesatpromoterstothe left
(pL)
andright
(pR)
of the immunity region
(Fig.
1) terminates atsites tLand tR,,respectively (20, 21, 27,
37).
In the presence ofthe N geneproduct,transcription
continues beyond these termination sites. By mutation,newpromoters(e.g.,c17or
ri5b)
have been isolated that initiate transcription to therightoftR1 (13, 23,25,26). This abnormal
tran-tPresent
addres:
CancerBiology Program, NCI Frederick CancerResearchCenter, Frederick,MD21701.tPresentaddress:Laboratoryof MolecularVirology, Na-tional Cancer nsitute, National Institutes ofHealth,
Be-thesda,MD20014.
scription terminates beforereaching gene Q if theN geneproductisabsent, but continues into
geneQif the N geneproduct ispresent(9).This
resultsuggested the existence ofasecond right-ward ternination site in addition to
tR,.
Thissite, designated tR2,waslocated between genes P and Q by analysis of two novel mutations, nin5(a5.4%deletion) and
byp
(5,8, 14, 19,28). Phage carrying eitheroneofthesemutations nolonger requirethe N geneproductfor transcrip-tion through tR2, although such read through wasenhanced in the presence of this product. Phagecarryingthenin5 deletionneed noother mutations to grow in the absenceof theNgene
product, whereas the c17 promoter mutation must be included with the
byp
mutation to achieve N independence. Because the N geneproductmustalsoactattRl for normal
synthesis
ofphagereplication proteins,it issurprisingthat
nin5phagegrowinthe absenceofthisproduct.
Thenin5 mutationmighteliminatethis
poten-tialreplication defect byeitherpermittingmore efficient
utilization
of the availablereplication
proteinsorincreasingtheirsynthesis.
The transcription of latephagegenes (genes
S, R, and A through J [Fig. 1]) is initiated
primarily atp'R
andrequirestheQgeneprotein(18). The modeofaction of theQgeneproduct
isunclearalthoughit has beensuggestedthat it
acts like the Ngene product in allowing tran-scription to continue beyond termination sites downstreamfromP'R (28).
Inthispaperwe describe theisolation, map-ping, and characterization ofmutationsthat
on November 10, 2019 by guest
http://jvi.asm.org/
ENQUIST
an tL PL
A J lnt rMed redp Wm N ci O P a S R
PR tmi c617 dCSb tR2 PR
L
L.hiimnht43 J I
lMwuMq21
..le nl
I
'l _a opue-3 16
-I
I
K
3.2 90.6
[image:2.501.73.397.66.231.2]Pam3Paau/ byp OAO wm21QmM57
FIG. 1. Geneticmap ofphage A. Geneticdistances are not drawn to scale, although the relative positions
ofthe genes andsitesarecorrect. The structural genes are placed between the two DNA strands, and the
genetic sitesareplacedabove and below the DNA strands.pL, PR,andP'Rrefer to the phagepromotersites; tL,tRi,andtR2refertothephage terninator sites;cl7and ric5b referto newpromotermutations;and att refers tothephage attachmentsite. Thetwoarrowsindicatethedirection ofphagetranscriptionduringlyticgrowth.
The region betweenphagegenesP andQisexpandedwith the A map coordinates of the left and right ends
ofthe nin5deletionandtheQA-Q80crossoverpointshown(11). low thephagetogrowbetter whenthesynthesis
of the Q gene product is limiting. Included
among these mutations are puq mutations, whose isolationsaredescribedinthispaper,and
the nin5andbypmutations.
MATERIALS AND METHODS
Bacterial andphage strains.The bacterial and
phagestrains used in this paper are listed in Table 1.
Media. Tryptone broth contains 10 g of tryptone
(Difco),5gofNaCl,and 5 ml of 1 MTrisperliterof water.TBMMB1istryptone brothsupplementedwith 0.4%maltose, 10mMMgSO4,and1
,g
of vitaminB1perml. TB top and bottomagarsaretryptonebroth
supplementedwith 7 or 12 g of agar(Difco)perliter, respectively. Eosin methylene blue-agar (EMBO) plates were prepared as previously described (30). TMGbuffer contains 10 mMTris-hydrochloride, pH
7.4, 10 mMMgSO4,and 0.1%gelatin.
Standard phageprocedures. Phagestocks were
prepared from purified plaques by the plate lysate method (29).Single-step growth experiments(33) and
phage crosses (32) wereperformed aspreviously de-scribed. Whenpotentialrecombinantplaquesfor per-tinent mutationswerescreened, thefollowing pheno-types were used:(i)c1ts857,clearplaquesat
420C;
(ii) by, clear plaques at32°C and the ability to form plaques on strains NS377 and NS577 (33); (iii) nin5, the ability to form plaques on strains NS377 and NS577 and a decreased sensitivity of the phage to EDTA inactivation (24,34); and (iv)puq, theabilityto allow a Qam phage to form a plaque on strain CA169(seebelow). Phageusedtoprepare extracts for thelysozymeassay werepurifiedfromplate lysates by
banding in aCsCl equilibriumdensity gradient (34).
Measurementoflysozymesynthesis.A culture of strain W3350 growinginexponential phaseatlol celsper ml was concentratedbycentrifugation (SS34
Sorvallrotorfor10minat6,000 rpm), and thepellet
was suspended in 1/5 volume of a 10 mM MgSO4
solution. A 2-mlaliquot of this cell suspension was infected withphageat amultiplicity of infection (MOI) of fourphagepercell. After5minat370Cforphage adsorption,the infectedcellsweredilutedwith 8 ml of tryptonebroth, andthe culturewasgrownwith
vig-orousaeration at37°C.Samples (1 ml)wereremoved atdesignated intervals, chilled to4°C, andsonically
treated with the fine probe of a Branson Sonifier (setting 3) for two 20-s intervals. The debris was removedbycentrifugation (15min at6,000 rpm in an SS34Sorvall rotor),and thesupernatantwasused to assaylysozymeaspreviouslydescribed(33).One unit oflysozymeis that amountofenzyme whichproduces
0.001decrease in cell absorbanceat 600 nm(A6s)in1
min. The extract was adjusted so that the enzyme level used in each assay reduced cell absorbance 0.1 to 0.2
Awos/inin.
Thebackgroundlysisof sensitized cells alone was 0.0005 Awo/min, and the activity isex-pressedasunitsper 100
pl
of infected cells.Measurement ofIy8ogenization frequency. A culture of strain YMC or 594growinginexponential phase at 10'cells per ml inTBMMB1 at 320Cwas concentrated by centrifugation, and the pellet was
suspendedin 1/10 volume ofa10mMMgSO4solution. Thecellswerestarved for30min at320C,and then 0.1-mlsamples(10W cells) wereinfectedat anMOI of either 5 or 0.2phageperinfected cell. After10min for phage adsorption at320C,dilutionsof these infected
cells were spread on EMBO plates on which 108 Ah80(attA-int)Ac selector phage had previously been
spread. These plates were incubated overnight at
320C,and colonieswerescoredasAlysogens.As the
infecting A phage in these experiments carried the cIts857mutation,wecouldindependently confirmthat thesecolonieswerelysogens bytheirinabilitytogrow at420C.Thelysogenizationfrequencyis the number oflysogensper number of infected cells, the latter
beingthecelltiteratthemomentof infection in the MOI=5experimentand 20% of this titer in theMOI
=0.2experiment.Wealso measured thelysogenization
frequency byspreadingtheinfectedcellson EMBO
I
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
PHAGE A MUTATIONS AFFECTING Q GENE ACTIVITY
TABLE 1. Bacteria and phagestrains
Strain Pertinent genotype Comments/source/reference
supF
YMC(A) YMC(Ximm434) YMC(Ximm2lSam7)
sup+8trr sup+
supE
supC
sup+nusA-1rif'-2
NS377(.o80psu3+)
N215(sup+)
OW
recA
XcIts857Pam80+(Qam501-attR)'
AcIts857Pam80+(Qam501-attR)"
X(attL-Pam80)AQam21+
A(attL-Pam80)AQam21+
X(attL-Pam80)AQam21+ X(attL-Pam80)Anin5Qam21+ X(attL-Oam29)APam3+nin5Qam21+
Phage
AcIts857Qam2l XcIts857Qam57 XcIts857Qam5Ol
AcI+Qam21
Aimm2lcItsSam7
AcIts857byp
Ximm434byp
XcIts8570am29byp
AcIts857b3pQam21
XcIts857nin5 AcIts857PamSOnin5 XcIts857nin5Qam2l
AcIts857nin5byp
AcIts857nin5bypQam21 AcIts857Pam80nin5byp AcIts857Pam3nin5Qam2l
XcIts857puq-3 (orpuq-16)Qam2l XcIts857puq-3 (or puq-16)
AcI+puq-3(orpuq-16)Qam2l Ah8Oatt8Oimm434(QSR)4080 Ah8O(attAint)ac
12;usedto assayAammutants
6 7 3 4
33;used to assayAbypand Anin5 phage
33; used to assay Abyp am or
Anin5am mutants 35
Phage strains were either
ob-tained from the National In-stitutes of Health collection
orwereconstructed by
recom-bination betweenappropriate A mutants, using selection procedures described in the text.
plates without selector phageandindividually testing theresulting coloniesfortemperaturesensitivity.The results obtainedbythe two methodswere identical.
RESULTS
Isolation ofpuqmutants.Weisolated
mu-tationsthatspecificallyaffect either the synthe-sis of theQgeneproductoritsactivityasfollows.
AQam mutants fail to form plaques on ahost
containing a supC ochre suppressor (strain CA169). Since these same phage form normal
plaquesonahostcontaininganamber suppres-sor (YMCsupF) that inserts the same amino
acid (tyrosine) at the amber site as the supC
suppressor,thegrowth defect of XQammutants
in strain CA169 is probably a consequence of theirinabilitytomakesufficientQgeneproduct. The levels of amber suppression in strains CA169and YMCare,respectively, about15and 50%(15).
WhenXQam2l isassayedonCA169, the
effi-ciency of plaque formation is about 6 x
1O-'
relativetothatonYMC. Of theserareplaques,
three classescanbe distinguished onthe basis
ofplaquesize onCA169. The first class, repre-senting about 10% of the plaque forners,
con-tainsphagethat formnormal-sizeplaqueswith
thesameefficiencyonstrainsCA169, YMC,and
594
(sup').
Thesephagearepresumablyamber'revertants. Asecondclass ofphage,also
repre-Bacterial YMC
NS62 NS617 NS61
594 W33S0
C600 CA169 NS377 NS577 NS460
N205
SA268 SA431 SA613 SA741 SA307
DC401
DC402
VOL. 30,1979 3
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.61.464.64.524.2]sentingabout 10%of theplaques, forms pinpoint
plaques on strain CA169 and 594, but normal-size plaqueson strain YMC. The efficiency of plaqueformationis about the same inall cases.
Thesemutants areprobablythe same as the "Q-independent(qin)"phage studied by Herskowitz and Signer (17) that exhibit an increased level of Q-independent transcription of late phage genes. These were not further characterized. The thirdclassofphage mutants, representing the remaining 80% of the plaque formers on strain CA169, made normal-size plaques on YMC, small plaques with unit efficiency on
CA169, andplaques with lessthan
10'5
efficiency on 594. Our hypothesis was that these phageacquired a second-sitepuq mutation, enabling themtogrow better when eitherthe synthesis oractivity ofthe Q gene product waslimiting. SinceApuqQam2l phagedonotformplaqueson
the
&itp+
host, we suggest that at least some Q gene product is essential for improvedphage growth. Three representative class 3
Qam2l pseudorevertants of independent
origin
(Qpuq3Qam21,
Apuq4Qam21,
andApuq16Qam21)werechosen forfurther analysis.
Specificity ofpuqmutations.The puq
mu-tationsmight allow AQam2l to grow better in
strainCA169because thesemutationsalterthe
Q-gene protein so thatits activity is increased orbecause these mutations increase thelevelof
Q gene expression. If the first explanation is correct, wemightexpect thatsuppression of the
Qam growth defect byaparticularpuqmutation
would bespecificforthemutationschosen;that
is, the protein alteration produced bythe puq mutationmight onlyincrease the specific
activ-ityof the protein whentyrosine is inserted by
an amber-suppressingtRNA at a particular
lo-cationin theprotein. If the secondexplanation
is correct, puq suppression of the Qam defect
should be independent of the particular Qam
mutation used.
To test these two possibilities, we constructed phage by recombination, carrying each of three puqmutations with three differentQam
muta-tions. We firstselectedQam+revertants ofeach
ApuqQam2l phage by selecting
normal-size
plaquesonstrain 594(sup+). The Qam+puq
mu-tants were then crossed with
XQam501
orXQam57
in strainYMC(supF).Theprogeny ofthecrosses wereplated on CA169. About 5% of the plaques were small (characteristic of Qam
pseudorevertants)and, of these, about 20% were indistinguishable in their properties from
ApuqQam(see above). The Qam mutation could
alwaysbe recoveredfrom theseApuqQamphage
in backcrosses with wild-type X by selecting
phagethatfailed to formplaquesonCA169 and 594butplated nornallyonYMC. We obtained the same resultswith all three puq
mutations,
soninecombinationsofApuq
Qam phage
(three
puqmutationsand three
Qam mutations)
have the same properties. Wesuggest that the puqmutationincreases thesynthesisof the Qgene product andnotthespecific
activity
of the pro-teinitself. Mappingdata described in thenext sectionplacethepuqmutationsat aposition
totheleftoftheQgene(Fig. 1),moredefinitively ruling out a direct alteration of the Q gene product by puq.
Mapping puq mutations. (i) Phage
crosses between
AimmAcIpuqQam21
and Aimm21cItsSam7. We identified Aimm-21cItsSam+ recombinantsamongthe progeny ofthiscross asclear-plaque fornersonstrainC600 at420C (Table 2A).Atotal of75 to90%ofthese
recombinantscarried theQam21 mutation and,
amongthese,60 to70% carriedthepuqmutation
(recombinant classa). We conclude thatthepuq
mutations tested (puq3, puq4, andpuql6) lie betweentherightend of theimm2l substitution and theQam21mutations. Because most recom-binants thatcarried theQammutation also car-ried the puqmutation,puqmustbe closetoQ. (ii) CrossesbetweenXimmAcI+puqQam21 andXcIts857
(Q-attR)A prophage
deletions.Wetestedabatteryofprophage deletions that
enterthe Xprophagefromtherightandremove
the Qgene, yet endwithin the region between genes 0 and Q (Table 2B). We infected these
lysogens withtheAimmAcI+puqQam21mutants
and selectedphagethatrescuedthecIts857 im-munitymarker from theprophage. Such phage form clear plaques on YMC at 42°C. If the
prophage deletion contains thewild-typeallele of puq, then someof the cIts857 recombinants
shouldhavelost the puqphenotype. Sincenone ofthe lysogenscarrythe prophageQgene, the
infecting phagecannot lose theQam mutation.
None of the recombinants tested (at least 100 from each cross) lost the puq mutation. As a
control, we used
XcI+Pam80
to infect eitherSA268 orSA431 andfound that about 10% of the AcIts857 recombinantprogenywere Pam'.
We suggest that the wild-type allele(s) of the puq mutations is removedbytheprophage dele-tions. These results and those shown in Table
2Aplace thepuqmutations between the left end oftheSA268 and SA431 deletions and the wild-type allele of the Qam21 mutation. We cannot
rule out,however, that the puq mutations map outside and just to the left of the SA268 and
SA431deletions.
(iii) Crosses between XcIts857puqQam21
on November 10, 2019 by guest
http://jvi.asm.org/
PHAGE A MUTATIONS AFFECTING Q GENE ACTIVITY TABLE 2. Mapping ofpuqmutatl
immn2l marker, the Q gene, andpi
A
imm,Acr
I
imm2lclt-pug mutation puq recombinant frc
14
16
B Infecting
pfropa
0.7 0.6 0.7 0.6 0.7
clt
-
c1tsa857-Prophage
Aclts8570am29+(Pam3.attR)A=SA439
Iclts857Pam3 +(Pam8OatR)A=SA444 Acils8S7PamO+(Qam2t-1tfR)6=SA268
Acits8S7Pam8O+(Qam2t1atIR)6=SA431
a
m(A)
Anm21cItsSam'
recombi asclear-plaque-formingphageonj(the Sam7 mutation isnotsuppr suppressor in strain C600). Eighl
recombinantscarrytheQam2lm
unable to form plaques on str
ionsrelative to the nants would be a measure of the relative dis-rophage deletions0 tances between imm434 and either puq or byp. Incontrast,ifpuqwerebetween byp andQam2l pvq
am21
+ (Fig. 2, order 2), we would expect all of the-J
Sam?Ximm434Qam
recombinants to carry thepuqmutation since the formation of puq+
re-combinants would require a triple recombi-tion[(a)/(a)+(b) nation event. For
puq3
andpuql6,
respec-tively, 12% (16/132) and 21% (22/106) of the
'o
Aimm434byp+Qam21
recombinants were puq+.so Therefore, both puq mutations are to the leftof ,,
Ibyp,
puq3
atafractional distance 0.88andpuql6
57 atafractional distance 0.79 of the distance
be-tweenthe
right
end of theimm434
substitution '° andthe byp mutation. Ifwe assumethatthese geneticdistances aredirectlyproportionaltothe puqGam21
physical
distancesbetweenthesemarkers,
then the puq mutationscanbelocated on the Aphys-,__,,,,,,,,,
ical map.Sincethe A mapcoordinate of therightend oftheimm434 substitution is79.1 (11) and
the right-most coordinate of the byp mutation q+phagt
anong
hekldt,57
mustbe90.6(see
below),
wecanlocatethepuq3
Q#m2t
recombinants and puql6 mutations on the A map betweenpuq-3 puq-16 coordinates 88 and 89. This would
put
thesepuqmutationsin the DNAsequences deletedbythe 0/120 0/123 nin5 deletion (DNA deleted between 83.6 and o/laO 0/110 89.2 on the X map) or just to the right of that
0/211 0/143 deletion
(Fig.
1).0/140 0/126 (iv) Crosses with
APam3nin5Qam21
andApuqQam67.
To determine whether the puqnantswere selected mutationsareindeed removed
by
thenin5dele-strainC600 at42°C tion, we crossed
APam3ninNQam2l
with ressed by the supEApuqQam57
(Table 3). The Qam57 mutation is ty percent of these located to the right ofQam2l
(18), and phageutation as
they
are with this mutationwill
form plaques on am594(sup').
WeYMC(supF)
butnot onC600(supE).Phagewithscreeneca tnes AunmMLcLUswamZI recomUmants ror
the puq mutation based on their ability to form
plaquesonstrainCA169.(B)We selectedphagethat
had rescued the clts857allele from theprophageas
clear-plaque-forming phage onYMCat42°C. These werescreened forthe puq mutationonstrain CA169.
andXimm434bypQ+. Topositionthe puq mu-tations with respect to the
byp
mutation,
we selectedXimm434Qam21recombinantphageas turbid plaque formers onYMC(supF)
at42°C
(phage carrying the byp mutation form clear
plaques [5]) (Fig. 2). The absence of the byp mutation among these recombinants was
con-firned by their inability to form plaques on
strainNS377(see above and reference 33). Since the byp mutation is located between phage
genes P and Q (seenextsection), thepuq
mu-tation might be located either to the right or
totheleft ofbyp. If puqwerebetweenimm434
and byp
(Fig.
2, order 1), the fraction ofXimm434puqQam21 phage amongthe
recombi-orderI
order 2
imm434 + byp +
cItsg5T puq + I m21
imm434 byp + +
[image:5.501.62.254.74.386.2]cJfts57 + P 0 1
FIG. 2. Turbid-plaque-forming recombinant phages(recombinants between theimm4andbyp
markers) were selected on strain YMC at 42C. Ninety-eight percent of these phage caried the
Qam21mutationasthey were unabletoform plaques
onstrain594(sup+).Thepresenceofthepuqmutation amongtheseXimm434byp'Qam2lrecombinantswas
assessed bytheir ability toformplaquesonstrain CA169.
VoL 30,1979 5
3
PU4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.266.460.452.656.2]6 STERNBERG AND ENQUIST
the Pam3 mutation will not form plaques on
C600(supE) at420C. Thus, onlyQam57+
Pam'
recombinant phage will form plaques on C600 (supE) at420C. These recombinantphagewere
purifiedandtested fornin5,puq, andQam2las
described in footnote a of Table3andabove. An
ambiguityarises in thisanalysisbecause it isnot possible to distinguish between Xnin5Qam2l
andXnin5puqQam2l because nin5isapuq
mu-tation (see next section). In addition, itis not
possible to distinguish between Xam+puq and
Xam+puq+.In any case, we wereunabletodetect anyQam21puq+nin+ phage,suggestingthat the
nin5 deletionremoves the wild-type alleles of puq3 andpuql6. Wecannotruleoutthe possi-bilitythat puq maps very closetobutjust out-sideofthe nindeletion.
byp
andnin5arepuq mutations. The puq,nin5,andbypmutationsarelocatedinthesame regionoftheX genome. Thisproximity encour-agedusto testnin5andbypfor the puq
pheno-type.To dothis,theQam21 mutationwas intro-duced into Xnin5 and Xbyp. Xnin5 and Xbyp phage were crossed with XQam21, and phage
[image:6.501.54.241.366.539.2]thatformed smallplaquesonCA169 (about2% of the yield from the cross) were selected for
TABLE 3. Mappingofpuq mutations relative to the
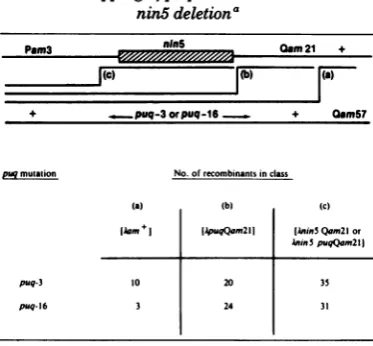
nin5deletiona
PsWnm3A Oem21 +
I(c) (b '(a)
+ ..puq-3orpuq-16 + 0am57
puqmuaation No. of recombinants in class
(a) (b) (c)
Aam+ (ApuqQam2lI (Auin Qam2lor
AinSpuqQam2l1
puq-3 10 20 35
puq-16 3 24 31
0The
level ofwild-type (am+) recombinants was measured as plaque formers on strain 594(sup+).AQam21recombinants were assayed on strain C600 at
420C
(neither AQam57 norXPam3 form plaques on C600 atthis temperature) and represent 10% of the totalphage burst. We determined whether the nin5 mutation was present intheseAQam2lrecombinants by measuring either their ability to form plaques on strain NS577ortheir EDTAsensitivity. Among theAQam21recombinants without the nin5 mutation, the presence or absence of the puq mutation could be assessedbytheability of these phage toformplaques onstrainCA169(supC).
further analysis. The small-plaque phenotype itself strongly suggested that nin5 and bypwere
puqmutations. We demonstrated that these Q pseudorevertants were either XbypQam21 or Xnin5Qam2l bythefollowingcriteria: like both the nin5 and byp parents, the recombinants formedplaquesonNS577 (33); thepresumptive
nin5 recombinant isasresistanttoEDTA treat-ment as is thenin5parent(wild-typeX is EDTA sensitive; deletions are EDTA resistant); and both phage do not form plaques on 594(sup+) but do plaque on YMC(supF). Furthermore, they both fail to complement XQam2l in 594(sup+). Because these phage are able to grow
on CA169, both byp and nin5 must be puq mutations.XbypQam2lfonnslargerplaques and Xnin5Qam2l forms smaller plaques than does ApuqQam2l on CA169. The kinetics of phage production by these Qam2l phage inCA169(see
below) confirm thisincreasing ability to express thepuqphenotype (byp>puq >nin5).
Mapping nin5 and byp mutations. Al-though both nin5 and byp mutations have been located in the interval between phage genes P and Q, their location relative to each other is unclear. (Are they at the same site? Is bypto the left orright ofnin5?) The following three
crosses locate more precisely these two muta-tions.
(i)
Crosses between either Pam8Onin5or XOam29byp andXcIts857 (Q-attR)A
pro-phage deletions. We infected each prophage deletion with XPam8Onin5 or
AOam29byp
andisolated
amber'
recombinantsfrom the progenybyplatingthe phage yield onstrain
594(sup')
(Fig. 3A). These
amber'
plaques were thenscreenedfor loss of nin5orbyp.Wedetectedno nin+ or byp+ plaques among several hundred
amber'recombinants. We conclude that the two
prophage deletions contained instrains SA268
and SA431 carryneitherthewild-typeallele of the byp mutation nor DNA to the right of the nin5 deletion.
(ii) Crosses between Xnin5Qam2l or
XbypQam2l and (attL-P)A prophage dele-tions. We used another set ofprophage dele-tions that enter the P-Q region from the left
(Fig. 3B). These strains were infected with
Xnin5Qam2l orXbypQam21. Amber'
recombi-nantswere selectedbyplatingthe phage yield
on594(sup'),andphage in the resulting plaques werethen tested for the loss ofthe nin5 or byp mutations.Among several hundred recombinant plaques obtained from the three crosses, none contained nin+ phage. Similarly, byp+ phage
were not found in crosses involving SA613 or
SA741.However,20% of theamber'phage from
on November 10, 2019 by guest
http://jvi.asm.org/
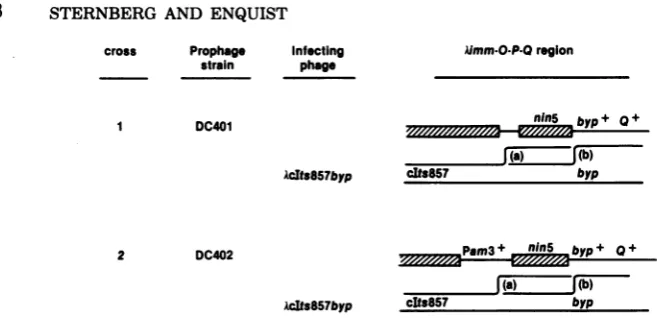
PHAGE A MUTATIONS AFFECTING Q GENE ACTIVITY
Cross Prophag Infecting
Strain Phage A) I SA268or
SA431
APamSOnin5
SA2S8or
SA431
10am29byp
8) 1 SA613or
SA741
AbypQam2l
SA307
Abyp0am2l
lo-P-Qregion
0+_P+________
Pam8O 0 +
o+P+ ,,aS,...
Oam29 byp Q +
byp 0am21
_____________bp _0+
byp ORM21
3 SA307
AninS5Om21
byp + a+ I
FIG. 3. Recombinantphage in these crosses between AOam, APam, and AQam phage and deleted A
prophages were selected asplaque formers on strain 594(sup+). Recombinants lacking thenin5 deletion
(crosses Al and B3)weredetectedby theirinabilitytoform plaquesonstrainNS377, and those lackingthe bypmutation(crosses A2,Bl,andB2)weredetectedby their turbid plaquephenotypeand confirmed by their inabilitytoform plaquesonstrain NS377.
the
AbypQam2l
xSA307cross werebyp'
(cross2 recombinant classa). SA307, therefore,
con-tains thewild-type allele of
byp
butnotDNAtothe leftonthe nin5deletion. We conclude that the
byp
mutation liesbetween the left endpointof the nin5 deletion andgeneQ.
(iii) Cro88e8 with Aclts857byp and
(attl-Oam29)A or (attL-Pam8O)A cryptic
pro-phage deletions derivedfromaAnin5
pro-phage.Todeterminewhether the nin5 deletion
removesthewild-type allele of
byp,
wecrossedXcIts57byp with prophage deletions derived froma?nin5 prophage. (Fig. 4). In bothcases, byp+ recombinants were detected. Two of four
byp+ recombinantsfrom the DC401 cross and three of eight byp+ recombinants from the
DC402cross weremoresensitivetoEDTA treat-ment than werenin5phage and failedtoform
plaquesonstrainNS577,bothexpected
charac-teristics ofbyp+ nin+ phage (recombinantclass
b). If thebyp+allelewerelocatedinaregionof
DNA removed by the nin5 deletion, then all
byp+ recombinants in these crosses should be nin5. As this is notthe case, the byp+ allele is
notcoveredby the nin5 deletion. Basedonthese data and those presented above, we conclude that the
byp
mutation is located between theright end of the nin5 deletion and the Qgene. IsolationofXnin5bypdouble mutants. If
nin5andbyp areatdifferentsites,it should be
possible toisolate the
nin5byp
double mutant. We did this by treating the progeny of the crossesshown inFig.4 with EDTAtoinactivatenin+ phage. The surviving phage were then
platedon
594(sup').
Ten to twenty percentof theresulting plaques wereclear and thuswerepotentialAnin5byp phage.Toverifytheir
geno-type, weisolated both nin5 and bypsingle
mu-tantsfromthe double mutant. Theexperiments were asfollows. We first crossed either Pam8O
or Qam2l mutations into the presumptive
Xnin5byp
phage and then crossed theXPam80nin5byp phage with XQam2l and the
Anin5bypQam2l phagewith XPam8O (Table 4).
In both cases amber+ recombinants were
se-lected and clear
(byp)
orturbid(byp')
plaques were tested for the nin5 mutation. We foundthat: (i) in cross 1, 30% of the turbid plaques
VOL. 30, 1979 7
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.115.417.60.350.2]8
cross Prophage Infecting strain phage
I DC401
2 DC402
Aimm-OP-Q region
nin5 byp+ °+
=
_
I )
J5b)
ArIts857byp c1tS857 byp
Pam3~+ WOl byp~ Q+
Acls857byp cUts857 byp
FIG. 4. Recombinantphageswith thebyp+allele were selected as turbid plaqueformers on strain 594 at
42°C. The EDTA sensitivity ofthephage contained in these plaques was tested under conditions in which
nin5phage are completely resistant and nin+phage are inactivated to a level of10-2 survival (EDTA treatmentfor10min at32°C).
containednin5byp' phage, and in cross 2, 35% ofthe clearplaques contained nin+ byp phage (in both cases, recombination occurred in seg-mentb;therefore, theoriginal phagemusthave carriedthenin5and thebypmutations); (ii)the location of byp between nin5 and Qam21 is
confirmed because all of the clear-plaque
for-mersin cross1werenin5,and all of the turbid-plaque
forners
incross 2 werenin+(recombi-nation in both crosses is in segment c);(iii) the relative recombination frequencies in intervals
a,b,andcare ameasureof the relativelengths of the intervals (based on the results given in Table 4, thevaluesfor these distancesexpressed
asafraction ofthe total recombination distance
betweenPam8O andQam2lare:segment a, 0.44
to0.49; segment b,0.21 to0.24; and segment c, 0.30to0.32).
Mapping
byprelativetotheA-480recom-bination site in gene
Q.
A unique site forrecombination betweenphagesA and
480
islo-cated within the Q gene of these phages at position 90.6ontheA map (11, 14). Asthissite isimmediatelytotherightofthe nin5deletion,
we decidedto use a
X-+80
hybridphage with A DNAtotheleft of this recombination site and480
DNAtotherightof thissitetolocalizemorespecifically the byp mutation. We infectedstrain
YMC with this hybrid phage and with
AbypQam2l (Fig. 5) and selected hAQam+
re-combinantphage from the phageyieldasplaque
forners on a +80-resistant derivitive ofstrain
N215. Three of 1,120 recombinantplaques were clearat
320C.
Presumably,they contained phage withthe byp mutation andwereformed byre-combination event c. The
bypQ480
genotype of theserecombinantphagewasconfirmedby thefollowing tests: (i) they forned plaques on NS577,aproperty of byp mutants; and (ii) when
anN205(recA) lysogen containing the presump-tive AbypQ4)80 prophage was infected with
Xred3imm434Ram5, plaquesappeared at an
ef-ficiency of< 10-4 relative to that found after infection of an N205(recA) lysogen carrying a
AbypQA prophage. Plaque formation by heter-oimmune R- phage on lambda lysogens of
N205(recA) is anindication of the transactiva-tion of the prophage R gene by gpQ of the infecting phage (36). Thefailure to transactivate the R gene from thepresumptive bypQ480
pro-phageimpliesthat itindeed carried
480
DNA to therightof the A-480recombination site. Trans-activation of the )80R gene isnotstimulatedbythe Q gene product of phage A (10, 36). We conclude that
byp
is located between therightend of thenin5 deletion (89.2onthe map) and the QX-Q480recombination site (90.6on the A map).
Single-step growth experiments. Up to
now,wehave discussedthepuqphenotype only
intermsofplaque sizeandplating efficiency.In thissection,wequantitatively analyzetheeffect
of various puq mutationson kinetics ofphage development andyield ofphage carrying Qam mutations. We used strains 594(sup+), CA169(supC), and YMC(supF) and infected
them with wild-type A, AQam21, AbypQam2l,
Anin5Qam2l, Apuq3Qam21, and
Anin5-bypQam2l (Fig. 6). In YMC(supF), all phages
grewwell,althoughAnin5Qam2lseemedtogive aslightlyreducedphageburst. The puq
pheno-type isexpressedin strainCA169(supC).Inthis host, AbypQam2l andAnin5bypQam21 grew
al-most aswellasAwild type.However, the growth of Apuq3Qam21, Anin5Qam2l, and XQam2l
phageswasincreasinglymore defective both in the kinetics ofappearance and in the yield of phage. In the
sup'
host, only the wild-type J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.62.391.58.217.2]PHAGE A MUTATIONS AFFECTING Q GENE ACTIVITY
TABLE 4. Orderingof Pam80, nin5, byp, and Qam2l mutations byfour-factor crossesa
cross1 a b c
+
n1nS
byp Oam2lPamSO + +
Recombinant Segmentin which amber+recombinant No. isolated frequency recombination event occured
bypnin+ 0 --
-bypnin 17 0.30 (c) byp +nin+ 28 0.49 (a) byp+nins 12 0.21 (b)
cross 2 a b c
+ + Oam2l
Recombinant Segmentin which amber + recombinant No. isolated frequency recombination eventoccured
bypnin+ 24 0.24 (b) bypnin
byp +ni + byp +ninS
44 0.44 32 0.32 0
(a) (c)
a(Cross1)Wild-type(am')recombinants were
as-sayed byplaqueformation on strain 594 and repre-sented2% ofthephageyield. Thirtypercentof these
plaqueswereclear(byp),andof 17 clearplaquestested
all contained phagewith the nin5 mutationbyvirtue of their increased EDTA resistance. Among the
re-maining 70% turbid plaques, 12 of 40 tested were
judgedtobe nin5 byvirtue ofboth their increased EDTAresistance and theirabilitytoform plaqueson strainNS377.(Cross2) In this cross 2.8%ofthephage yieldweream'recombinantsand of these, 32% were
turbid(byp')and 68% were clear(byp).The presence of the ninallele among these recombinants was as-sessedasincross 1.
phage grew normally. Anin5bypQam21 and
AbypQam2lproduced onlytwo- tofourfoldmore
phage than did otherAQam2l phageinthis host. Whereas AbypQam2l and Anin5Qam2l phage did not forn plaques on 594(sup+) [less than
10'
relative to plaque formation on YMC(supF)], Xnin5bypQam21 phage fonned pinpointplaqueson594(sup+) withalmost wild-typeefficiency.Kineticsofendolysinproduction.The
ef-ficiencyof activation of latephagegenesbythe Q geneproductisgenerally assayedby
measure-0
a
I
a.
ment of phage endolysin (the phage R gene product) (9, 22, 33). We measured endolysin
after infection of strains YMC(supF),
CA169(supC), and 594(sup+) by XQam2l and
XQam+ phages (Fig. 7). In YMC(supF), all phages produced normal or near-normal amounts ofendolysin activity, ranging from 800 to1,000 unitsby 40 min after infection (Fig. 7a). After infection of CA169(supC) with XQam2l, endolysinproductionwasmuch reduced relative
h8O att8O Imm434' 080
(a) (b) (c)
hA attA cItS857 byp Qam2l
FIG. 5. AbypQp8O recombinant phage were de-tected as clear-plaque-forming phage on strain
NS460 at 32°C. Wecould confirm that these clear
plaquescontainedphage with the bypmutation as thesephageformedplaquesonstrain NS377.
TimeafterInfection(min)
FIG. 6. These single-step growth experiments were
performed aspreviously described (33). Thephage
yieldwassampledatdesignated times after infection
byblendingsamplesoftheinfectedcellculture ina Vortex mixerwith several dropsof chloroform. The bacterialstrainsusedwereYMC(a), CA169 (b), and 594 (c). The phage used were XcIts857Qam+ (0), AcIts857Qam21 (A), AcIts857nin5Qam21 (A),
Ac1ts857puq-3Qam21(0),AcIts857bypQam21(),and
AcIts857nin5bypQam21 (0).
9 VOL. 30,1979
on November 10, 2019 by guest
http://jvi.asm.org/
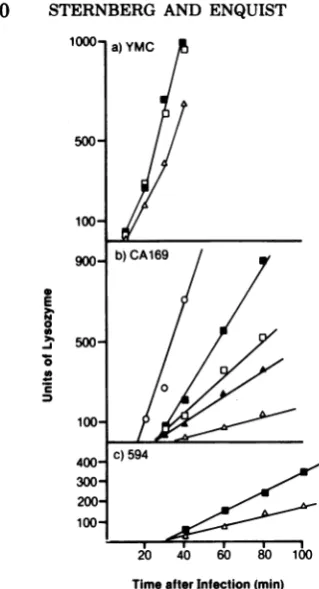
[image:9.501.52.448.70.559.2] [image:9.501.265.446.302.567.2]E
000
400- c)594
300-200
100
20 40 60 80 100
[image:10.501.60.220.62.357.2]Time after Infection (min)
FIG. 7. Lysozyme assays were perforned as
de-scribedin the text. Thebacterial strainsusedwere
YMC (a), CA169 (b), and 594 (c). The symbols for phage usedwerethesame asthosegiven in the legend toFig.6.
to the YMC(supF) infection. The nin5, puq3, and byp derivatives ofAQam2l produced,
re-spectively, increasingamountsofendolysinafter
infectionofCA169(supC) (Fig. 7b).In contrast,
endolysin production by both XQam2l and
AbypQam2lin594(sup+)wasmuchreduced;the
byp mutation enhanced enzyme production by onlyafactor of 2(Fig. 7c). Comparingtheresults shown inFig.7b andc, weconcludethat thebyp mutation produces atwofoldincrease in
endo-lysin synthesis in the absence of the Q gene
product (XQam2linstrain594)anda morethan
sixfold increase inendolysin synthesiswhen syn-thesis of theQgeneproductislimiting(AQam2l instrain CA169).
Clear-plaque phenotype of Abyp phage. Phage that carry the byp mutation form clear
plaquesandareimpairedintheirabilitytoform lysogens. The frequency of lysogen formation
was generally three- to sixfold less than that found forbyp+ phage (Table5). Wehave shown
inthetwoprevioussectionsthatthe byp
muta-tion increasestheexpressionoftheQgeneand latephage functions. This increasedexpression
could channel a phage infection into a lytic
rather thanlysogenic response. We would expect that if this were so, XbypQam2l phage would
form lysogens with a higher frequency than )bypQ+ phage inYMC(supF) and with even a
higher frequency in 594(sup+). This is, in fact,
what weobserved(Table 5). InYMC(supF)and 594(sup+) hosts, the Qam21mutation increased the frequency of lysogen formation by Abyp phage, respectively, two- andfourfold. Indeed,
XbypQam2l
forms a more turbid plaque onYMC(supF)than does
Xbyp.
DISCUSSION
Inthis paperwehavedescribedthe isolation and characterization of mutations (designated puq)inphageA thatincreasesynthesisof theQ
gene product. Normally, these mutations have no phenotype because the level of the Q gene productsynthesized by wild-typeAorby
XQam
phage grown in strainYMC(supF) ismorethan sufficient for phage production. However, if
XQam
phage are grown in the weakam-sup-pressing strain CA169 (under these conditions, synthesis of the Qgene product islimiting for phagegrowth), thenthe puq mutation helpsto
alleviate
thephage growthdefect. Thus, whereasXQam21
fails to plaque on strain CA169,ApuqQam2l
plaqueswith unitefficiencyonthishost.SincepuqQam phagedo not grow in strains
lacking either am or oc suppressors, at least somesynthesis of the
Q
geneproductis neces-saryfor puqtoexertanyaffectonXQam
plaqueformation;
puqdoesnotcreateaQ-independentphenotype,asdomutationspreviouslydescribed
by Herskowitz and Signer (17). Various puq
mutations have been mapped between phage genesP andQ,aregion ofthe genome knownto containtwomutations(nin5andbyp)that allow A to grow better in the absence of the phage
gene Nproduct.Bothbypand nin5 are also puq
mutations. Based on plaque size, phage burst, and phage endolysin synthesis, byp is a better
TABLE 5. LysogenizationfrequencyofAbypphagea
Lysogenization
Bacterial frequency at:
Infecting phage strainI
=5 =0.2
AcIts857 594(sup+) 0.4 0.5
XcIts857Qam21 594(sup+) 0.25 0.36
AcIts857byp 594(sup+) 0.08 0.12
XcIts857bypQam21 594(sup+) 0.30 0.40
AcIts857 YMC(supF) 0.5 0.25
AcIts857Qam21 YMC(supF) 0.25 0.30
AcIts857byp YMC(supF) 0.10 0.06
AcIts857bypQam21 YMC(supF) 0.17 0.12
aLysogenizationwasmeasuredasdescribed in the text.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.501.255.447.527.651.2]PHAGE A MUTATIONS AFFECTING Q GENE ACTIVITY
puq mutationand nin5 isaworsepuqmutation than are the mutations (puq-3 and puq-16)
whoseisolationsaredescribedin this paper. A mapof theAgenome,withparticularfocus ontheregion between phage genes P and Q, is shown inFig. 1. The map coordinates of the nin deletion are well documented (11). Crosses
de-scribed in Results located the puq mutations
either underthenindeletion or veryclosetothe
leftorrightendofthisdeletion.Thebyp
muta-tion waslocalizedtotherightof thepuq muta-tions andthe nin deletion(rightmapcoordinate
89.2)andtotheleft of thecrossoverpointin the Q genes of phageAand phage80 (map
coordi-nate 90.6). We have isolated Anin5byp double
mutants,confirmingthe locationof byp.
Based on ouranalysis of the properties of nin,
byp,and puqmutations,we candraw the
follow-ing conclusions about theregulation ofQgene
expression andtheactivityof the Qgene
prod-uct.
(i) The byp mutation increases the level of phageendolysin synthesis byafactorofnomore than2when the Qgene productis not synthe-sized (Fig. 7). Transcription ofthe phage
endo-lysingene(geneR)intheabsenceof theQgene
product initiates exclusively at pL and reads
throughinto genes Qand R (9). Since R gene
transcription is enhanced twofold by the byp mutation, Q gene transcription should be
en-hanced to the sameextent.Yet, whenalimiting amount ofQ gene product issynthesized (Fig.
7b), theenhancement ofendolysinsynthesis by bypisinexcessof sixfold.Theseresultsindicate that theconcentrationdependenceof the activ-ity of the Q gene product is not linear. This might be the case if the product of gene Q is
requiredas adimeror atrimer. In sucha circum-stance,atwofold reductioninmonomer
synthe-sismightmeanafourfoldreductionor even an eightfold reductioninactivity.
(ii) Abyp phage form clearplaques and lyso-genize standard bacterial hosts withathree-to sixfold-reduced frequency relative to byp+ phage. In contrast,
XbypQam
phage lysogenize strain594(sup')
at the same frequency as doXbyp+
phageand lysogenizestrain YMC(supF)at a frequency internediate between
Xbyp
andXbyp+
phage (Table 5). These results suggestthat afterinfection thelysogenization defectof
Abyp phage reflects an increased expression of
the Q gene and late phage functions, with a
consequent channeling of the infection into a
lytic rather than the lysogenicresponse. If the
synthesisofQgeneproductisreducedor
elimi-nated, thenthelysogenizationfrequency ofbyp
phage is thesame asthat ofbyp' phages.
(iii)
How do nin5, byp, and puq mutationsexerttheir affectsongpQactivity?We can rule outthepossibilitythat thesemutationsdirectly alter the Q geneproteinbecausenin5 is a
dele-tionthat maps to theleft of the Q gene, and the puqphenotype exhibitedby puq-3,puq-16, and byp is independent of the Qam mutation used (see Results; data for byp is not shown). More likely, these mutations act by increasing Q gene
transcription. As we cannot detect endolysin production from Apuq-3, Apuq-16, Anin5, and
Xbyp lysogens(data notshown; also see reference
5), these mutations mustnot create a constitu-tive Q and R gene promoter like the qinlOl mutation (10). Since theregion between phage genes P and Q contains a transcription
termi-nation site
(tR)
that is sensitive to the N gene product,thepuq, nin5, and byp mutations might alteroreliminate this sitesothattranscriptionbeyond itbecomes Nindependent. If
transcrip-tion antitermination mediated by the N gene product wereless than 100% efficient attR2,then thesemutations would enhance transcription of downstream genes (Qand R) even in the pres-enceof theNgeneproduct. Indeed, transcription stimulated bythis productdecreasesasthe num-berofterminationsites traversed by the
polym-erase increases (1, 2), suggesting that N-me-diatedtranscriptionantitermination is not 100% efficientateach terminationsite.
Althoughthismodelfitswellwiththe nature of the nin mutation, a deletion which could
eliminate the tR2 termination site, it does not
readilyexplainsomeof the properties of thepuq andbypmutations. First,whereasnin5andbyp mutationsare N-independentmutations, puq-3 andpuq-16 are not
[XNam7Nam53cl7byp
andXNam7Nam53nin5
plaque onstrain 594(sup+),but ANam7Nam53cl7puq does not (data not
shown)].
Yet,puq-3andpuq-16 mutationsshowa stronger puq phenotype than does the nin5
mutation(Fig.6and7). Thesediscrepanciesare
mosteasilyreconciledifthe puq mutation does
notaffectatermination site but rather creates
apromoter upstream from tR2. Thus, itsability
to stimulate Q gene transcription would still dependonNproductactivity.Second,sincebyp is not covered by the nin5 deletion, if nin5 deletestR2,then byp must either affect a second
transcription termination site in thisregion or must create a new promoter capable of tran-scribing theQ gene in theabsence of the N gene product.Assuchapromoter is not active in the prophage state (5), wesuggest that it requires upstream transcription (e.g., from pR) for its activation.Inthe absence of theNgeneproduct, the lowlevel oftranscription that might proceed through thetR2sitecouldactivate this new pro-moter, resulting in an increased Q gene
tran-VOL. 30,1979 11
on November 10, 2019 by guest
http://jvi.asm.org/
scription.
In additionto the nin5, byp, and puq muta-tions, a fourth mutation which maps in thetR2
region of the A map is pasB (31). This mutation enables a Xred-gam- phage togrow better in strainscontainingapolA mutation (thefeb phe-notype [38]). Preliminary experiments indicate that nin5 and puq-3 are pasB mutations but that byp is not. As the pasB phenotype is not
well understood, thesignificance of these find-ings must await further experimentation.
An analysis of the transcription patterns in the P-Qregion of the A map produced by the various mutant phages shouldhelp to elucidate their mode ofaction.
ACKNOWLEDGMENTS
We thankR.Weisberg for his support and encouragement during thecourseof thiswork and L.Stockmanforcarefully typing thismanuscript.
LITERATURE CITED
1. Adhya, S.,and M.Gottesman. 1978.Control of tran-scription termination. Annu. Rev. Biochem. 47:967-996.
2. Adhya, S.,M.Gottesman,B. deCromburgghe, and D. Court. 1976. Transcription termination regulates gene expression,p. 719-730. InR. Losick and M. Cham-berlin(ed.), RNA polymerase Cold Spring Harbor Lab-oratory,ColdSpringHarbor, N.Y.
3. Appleyard, R. K. 1954. Segregation ofnew lysogenic typesduring growthof adoubly lysogenicstrainderived from Escherichiacoli K12.Genetics39:440-454. 4. Brenner,S., and J. R. Beckwith.1965.Ochre mutants,
a newclassofsuppressiblenonsensemutants.J. Mol. Biol.13:629-637.
5. Butler, B., and H.Echols. 1970.Regulationof bacterio-phage Xdevelopmentby gene N:properties of a muta-tion thatbypassesNcontrol of lateprotein synthesis. Virology40:212-222.
6. Campbell,A. 1965. The steric effect inlysogenizationby bacteriophage A. II. Chromosomalattachment of a b2 mutant.Virology27:340-345.
7. Campbell,A., and E.Balbinder. 1958. Propertiesof transducingphage. CarnegieInst.WashingtonYearb. 57:386-389.
8. Court, D., and K. Sato.1969.Studies on novel transduc-ing variantsoflambda:dispensibilityof genes Nand Q. Virology39:231-252.
9. Couturier,M., C.Dambly,and R.Thomas.1973. Con-trol ofdevelopment in temperate bacteriophage. V. Sequential activation of the viral functions. Mol. Gen. Genet. 113:244-250.
10. Dambly, C.,M.Delstanche,and A. M.Gathoye.1979.
qinlOl:promoter mutation which allows the constitu-tive expression of the late genes. J. Virol. 30:14-20.
11.Davidson,N., and W. Szybalski. 1971. Physicaland chemicalcharacteristics of lambdaDNA, p.45-82. In A. D. Hershey (ed.), The bacteriophage lambda. Cold SpringHarborLaboratory,ColdSpringHarbor,N.Y. 12. Dennert, G.,and V.Henning.1968. Tyrosine-incorpo-ratingambersuppressors in Escherichia coli K12. J. Mol.Biol. 33:322-329.
13.Dove, W. F., H. Inokuchi, and W. Stevens. 1971.
Replicationcontrol inphagelambda,p.747-771.InA.
D. Hershey (ed.), The bacteriophage lambda. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y. 14. Fiandt,M., Z. Hradecna, H. A. Lozeron, and W. Szy-balski. 1971. Electronmicrographic mapping of dele-tions, insertions, inversions, and homologies in the DNAs of coliphages lambda and phi 80, p. 329-354. In A. D.Hershey (ed.), The bacteriophage lambda. Cold Spring HarborLaboratory, ColdSpringHarbor,N.Y. 15.Gorini, L 1970. Information suppression. Annu. Rev.
Genet.4:107-134.
16.Herskowitz, L. 1973. Control of gene expression in bac-teriophage lambda. Annu. Rev. Genet. 7:289-324. 17.Herskowitz, I., and E.Signer.1970. Control of
tran-scription from the r strand ofbacteriophage A. Cold Spring Harbor Symp. Quant.Biol. 35:315-323. 18.Herskowitz,I., and E.Signer.1970.A siteessential for
expression of all late genes inbacteriophageA. J.Mol. Biol. 47:845-856.
19.Hopkins,N. 1970.Bypassingapositiveregulator: isola-tion of aA mutant that does notrequire N product to grow.Virology 40:223-229.
20. Kourilsky, P.,L.Marcaud,M. M.Portier,M. H. Za-mansky, and F. Gros. 1968. Studies on the messenger RNA ofbacteriophage A. I. Various species synthesized early after induction of theprophage. Proc. Natl. Acad. Sci. U.S.A.61:1013-1020.
21. Kumar, S., K.Bovre,A.Guha, Z.Hradecna,V. M. Maher, and W.Szybalski.1969.Orientationand con-trol of transcription of E. coli phage A. Nature (London) 221:823-824.
22. McMacken,R.,M.Mantei,B.Butler,A.Joyner,and H.Echols. 1970. The effect of mutations in the clI and cIII genes ofbacteriophageA onmacromolecular syn-thesis in infected cells. J. Mol. Biol. 49:634-654. 23. Packman,S.,and W. S.Sly.1968.Constitutive A DNA
replication byA c17, a regulatorymutant related to virulence.Virology34:778-781.
24. Parkinson, J.,and R.Huskey.1971.Deletionmutants
ofbacteriophagelambda. I. Isolation and initial char-acterization. J. Mol. Biol. 56:369-384.
25. Pereira DaSilva, L.H., and F. Jacob. 1968. Etude genetique d'une mutation modificant la sensibilitea l'immunite chez le bacteriophage lambda. Ann. Inst. Pasteur Paris 115:145-158.
26. Roberts, J. W.1969.Promoter mutation in vitro. Nature (London)223:480-482.
27. Roberts,J.W. 1969. Termination factor for RNA syn-thesis. Nature(London)224:1168-1174.
28. Roberts,J. W. 1975.Transcriptiontermination andlate controlinphagelambda. Proc. Natl. Acad. Sci. U.S.A. 72:3300-3304.
29. Schrenk,W.J.,and R. A.Weisberg.1975. Asimple method formakingnewtransducinglines ofcoliphage A. Mol.Gen. Genet.137:101-107.
30. Shimada, K., R. Weisberg, and M. E. Gottesman. 1972.Prophagelambdaatunusual chromosomal
loca-tions.I.Locationofthesecondary attachmentsites and
theproperties of thelysogens.J. Mol.Biol. 63:483-503.
31. Shizuya,H.,D.Brown, and A.Campbell.1974.DNA polymeraseI-dependentmutantsofcoliphagelambda. J.Virol. 13:947-952.
32. Sternberg,N.1976. Ageneticanalysisofbacteriophage A headassembly.Virology71:568-582.
33. Sternberg, N. 1976. A class ofrifR RNA polymerase mutations that interferes with theexpression of coli-phage A late genes.Virology73:139-154.
34. Sternberg,N.,D.Tiemeier,andL.Enquist.1977.In vitropackaging ofaADamvectorcontainingEcoRI DNAfragments of Escherichia coli andphageP1. Gene 1:255-280.
35. Sternberg,N.,and R. A.Weisberg.1975.Packagingof
on November 10, 2019 by guest
http://jvi.asm.org/
PHAGE A MUTATIONS AFFECTING Q GENE ACTIVITY prophage and host DNA by coliphageA.Nature
(Lon-don) 256:97-103.
36. Szpirer,J., and P. Brachet. 1970. Relation physiolo-giquesentreles phagestemperesAet N80.Mol.Gen.
Genet. 108:78-89.
37. Szybalski, W., K. Bovre, M. Fiandt, S. Hayes, Z. Hradecna, S. Kumar, H. A. Lozeron, H. J. J.
Nijk-amp,and W. F. Stevens.1970.Transcriptional units
and their controls in E. coliphage A: operons and
scrptons.Cold Spring Harbor Symp. Quant. Biol. 35: 341-353.
38. Zissler, J., E. Signer, and F. Schafer. 1971.Therole of recombination in growth of bacteriophage lambda.I. Thegammagene, p.455-468.InA.D.Hershey(ed.), The bacteriophage lambda. Cold Spring Harbor
Labo-ratory,Cold Spring Harbor, N.Y.
VOL. 30,1979 13
on November 10, 2019 by guest
http://jvi.asm.org/