JOURNAL OFVIROLOGY,Jan.1981,p.336-342 Vol. 37,No. 1 0022-538X/81/010336-07$02.00/0
Nucleotide
Sequence of the Chi Recombinational
Hot
Spot
X+D
in
Bacteriophage
Lambda
GERALD R. SMITH,`* MICHAELCOMB,' DENNIS W. SCHULTZ,' DONNA L. DANIELS,2 AND FREDERICKR. BLATTNER2
Institute of MolecularBiology, University of Oregon, Eugene, Oregon97403,'and Laboratory of Genetics, McArdle Laboratory, University of Wisconsin, Madison, Wisconsin537062
Chisites inbacteriophageAstimulate recombination promotedby the RecBC
pathway of Escherichia coli. Mutations which create these sites occur at four widelyseparated loci inA. We report here the nucleotidesequence surrounding the site ofoneoftheseloci,
XD,
locatednearthe S gene. The mutationscreating the activeChi site, designatedX+D,
aretransversionsfromG
toT.
Thismutation, like theX+B
andX+C
mutations previously analyzed, leads to a nucleotidesequence common toallthree activeChi sites.
Generalized recombination in
bacteriophage
A isstimulated
by
special sites,
calledChi,
when recombinationproceedsvia theRecBCpathwayof the host ofA, Escherichia coli
(see
reference24for a review).Onehypothesis ofthe
mecha-nism ofthisstimulationsupposes thatChisites are
special
nucleotide sequencesrecognized by
oneof theenzymes intheRecBC
pathway.
To substantiate thishypothesis,
wehaveexamined thenucleotidesequencessurrounding
Chi sites todetermine whether Chiisaunique
nucleotidesequence(22, 23). Extensivesequencehomology
wasfound in thesequences of twowidely
sepa-ratedChi sitesinA(22).Fromthoseanalyseswe
could not concludehowmuchofthehomology
wasessential for Chi activity. Acomparison of
other sequences containing Chi would aid in
revealing
the extent ofthe common sequencenecessary forChi. Inthis paper we report the
nucleotidesequence of a third ChisiteinA and
compare that sequence with the two Chi
se-quencespreviouslydetermined.
Wild-type A is devoid of Chi,but mutations
which create Chi occur at the four
well-sepa-rated loci shown in Fig. 1 (10, 25). Sequence
analysis of the
XB
and xC loci was possiblebecause of theavailability of deletionmutations
with endpointsnear the Chi sites. These
dele-tions allowed thegenetic and physical mapping
essential for locating an interval smallenough
for convenient nucleotide sequence analysis.
Themutations creating Chi at the
XB
andXC
lociwere found to be single-base-pair changes (22, 23).
Asimilaranalysisofthe
XA
andXD
loci wasnotimmediately possible since these loci are in
regions of genes essential for A growth. The
availabilityof A DNAfragmentswithendpoints
near
XD
andinserted into self-replicatingplas-midsallowedthegeneticandphysical mapping
and thenucleotidesequenceanalysisofxD
re-portedhere.
(Weuseherethenomenclature of Maloneet al.[12].
X+
designatesthegenotype of theactive formof Chi sitesin A.X°designates theinactive form.XA, xB,
xC, andXD
designate the lociinAwhere the
X'
mutationsmay arise. Chi referstothephenotype ofthesites.)
MATERIALS
AND METHODSBacterial strains. E. colistrainswith Aprophage
deletions extending from galthrough attL and into theQtoSregionwereisolated and characterizedby Shapiro andAdhya (19). More detailed mappingwas
described by Herskowitz and Signer (11). Strains MS5061 and MS509wereobtained from thecollection of Frank Stahl (University of Oregon), and strains MS1118 and MS1153wereobtainedfrom the collec-tion ofJeffrey Roberts (Cornell University). Strain
594 (28) is lacgal Strr and doesnotsuppressamber (sus) mutations.
E. coli strains withplasmidsinto whichfragments
ofA DNA were inserted were obtained from other
investigatorsasfollows. From David Wilson (Cornell University) weobtained strainD264, which contains plasmidpDW1,aderivative ofplasmid pBR322 (26)
carrying EcoRI fragment F ofA (27) with a BamHI
linker attachedtotheright cohesive end.Inaddition tobearingthisplasmid,strain D264 isHfrC thi-1trpR rbs-115 leu. From Jeffrey Roberts and Michael Schechtman (CornellUniversity) we obtainedstrain
Q,which containsplasmidpOPII-APQ,aderivative of
plasmid pMB9 (3) carrying a fragment of the lac operon withUV5 mutation (21) and EcoRIfragment Eof A(27)bearingthe nin-5 mutation(5).Inaddition tobearingthisplasmid, strainQ carriesadeletionof
the lac-proregionofthechromosomeand anepisome
336
on November 10, 2019 by guest
http://jvi.asm.org/
SEQUENCE OF
X'D
RECOMBINATIONAL HOT SPOT 337Xloci
?genome
Right end of
XA XB XC XD
A t t , Jt t ~~~~~~cIt~ t R,
0.0 0.2 OA :0.6 5.8 1*i
I"MS"deletions
Eco8[ / / Hindm EcoRI H
A P. S
R0
0.8 0.9 1.0\'
ALDNA Tragments L !I\
inserted into plosmids / \
pRK112
PDW1
XD
/HaM A/UIl HhaI
15 AATTCATTAGTAATAGTTACGCTGCGGCCTTTTACACATGACCTTCGTGAAA TGGCAGGAGGTCGCGCTAACAACCTCCT 3
A3'CT TAAGTAATCATTATCAATGCGACGCCGGAAAATGTGTACTGGAAGCACTTTCc CCACCGTCCTCCAGCGCGATTGTTGGAGGA 5
EcoRI m
;inX+Dmutants
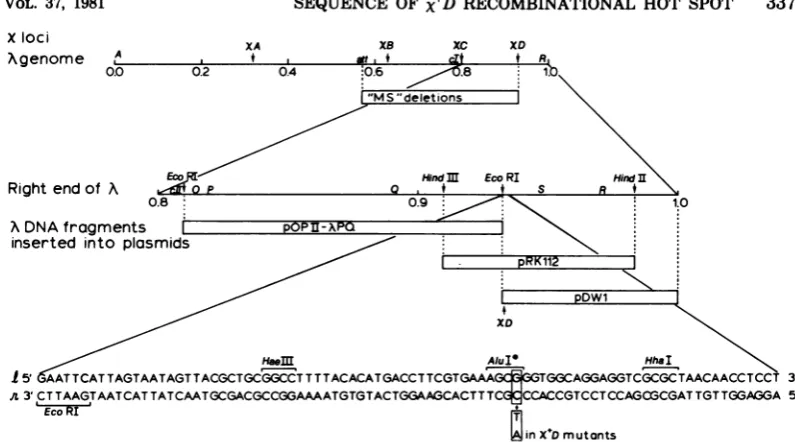
FIG. 1. Genetic andphysical map of phage A, showing the location and nucleotide sequence of theXD
locus. Oneach line,physicaldistances aredrawnto scale. The top line shows thefour lociat which X+
mutations arise, genes A, J, cI, and R, and theprophage attachment site att atthe indicatedfractional
distances from the left end of the A chromosome (7). Theextentsof prophage deletions in strains MS509,
MS1118,MS1153, and MS5061, usedtomapXD(Table1),aredrawnrelativetothepositions indicatedonthe
top line. The second line shows therightendofA, includingtheXD locus, genes cIIthrough R, and the positions of certain endonuclease cleavage sites. The extents ofA DNA fragments generated by these endonucleasesareshown relativetothe second line. Thesefragmentsarepresent in the indicatedplasmids,
whichwereusedtomapXD(Table2). The bottom line shows the nucleotide sequence surrounding the
X'D
mutations, which alter the basepairoutlined(Fig.2).Hyphensareomittedforclarity. Brackets above and below the line indicaterecognitionsequencesforthe indicated endonucleases.AluI* isacleavage site present only in DNAfromX Dmutants(Fig. 3).1andrrefertotheconventional stranddesignationsinA.
with thelacIQmutation (15). FromWilliamReznikoff
(University of Wisconsin) we obtained plasmid
pRK112A, aderivative ofplasmid ColElbearing the kanamycin and neomycinresistance determinants of Tn5 (1) and HindIII-HindII fragment A2 of A (18) bearing the susS7 mutation(9). Strain C600(thr-1 leu-6thi-1 lacYl supE44 tonA21) wastransformed with
thisplasmidDNAtogeneratestrain FB1034.
Phage strains. Aint4 red3 gam210X+D123susS7 and XtsJ15 red3gam210 c126X+D124wereobtained from JeanCrasemann(University ofOregon). Ab1453
X+D123 susR5 was obtained from Frank and Mary
Stahl(UniversityofOregon).Otherphageswerefrom
ourcollectionor werefromcrossesbetween themand those listed above. susP and susR mutationsare de-scribedbyCampbell (4).
Phage crosses. Cellscontainingplasmidsor
par-tially deleted prophages were grown at 37°C in a
mediumcontaining1%(wt/vol)tryptone (Difco Lab-oratories, Detroit, Mich.) and0.5%(wt/vol) NaClin
water,supplementedwith 0.1%(wt/vol)maltose.Ata celldensity ofabout 3x 108/nl,fivephagespercell
wereadded.After 20 minat37°Cwithoutshaking,to allow phage adsorption, theinfectedcellswerediluted 1:1,000 inthe above medium.Afteraeration at37°C for 2h,CHC13wasaddedtokillremainingcells.Sus+
recombinants were selected on strain 594 on plates
containing1%(wt/vol) Trypticase (BBL Microbiology
Systems,Cockeysville,Md.), 0.5% NaCl, and1%agar
(Difco). Sus+ recombinants were scored for Chi as
describedinfootnoteaof Tables1and2.
Enzymes. Sources of some of the enzymes have been describedpreviously (22, 23). In addition, endo-nuclease MspI was purchased from New England BioLabs,Beverly,Mass.
Preparation of DNA and DNA sequence
anal-ysis.Wild-type
(k0D)
DNAwaspreparedfromAcI60susS7 (22). Mutant
(W+D)
DNA wasprepared fromAcI857X+D123susS7 and fromAb2red3 gam210 cI26
X+D124susS7.Samples ofapproximately 100 agwere
digestedfor18 h at37°Cwith20Uof endonuclease
EcoRIinareaction volume of1.0 ml. Afterphenol
extraction and ethanol precipitation, the DNA was
labeled withpolynucleotide kinase by the exchange
procedure(2), usingapproximately5Uof kinasein a
reactionvolume of100pl.Reactionwasfor90minat 37°C. Afterprecipitation ofthe DNA withethanolin
thepresence of ammoniumacetate(14),the DNAwas digested for 12 hat 37°Cwith 8U of endonuclease
MspIinareaction volumeof 50pl. Aftertheaddition ofsucroseand markerdyes,the DNAwassubjected
to electrophoresis in a 7% polyacrylamide gel
(Bio-RadLaboratories,Richmond,Calif.)in Ebuffer (20) untilxylenecyanolFF hadmigratedabout7 cm.The third mostrapidlymigratingband,at apositionabout
two-thirdsof thewayfrom theorigintoxylenecyanol VOL. 37, 1981
'I rNKIA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.496.52.450.61.282.2]338 SMITH ET AL.
FF, was detected byautoradiography, electroeluted,
andsubjectedtonucleotidesequenceanalysis (13, 14).
RESULTS
Geneticandphysicallocation ofXDlocus.
Theprecise location of the
XD
locus wasessen-tial toidentifyaregionof Asufficientlysmall for
convenient DNA sequenceanalysis. Stahletal.
(25) mapped the A+D123 allele to a position in
orbetweengenesQ and S.Wemapped thisand
another
X+D
allele(X+D124),
both ofsponta-neous origin (J. Crasemann, personal
commu-nication), between the endpoints of two dele-tionsby usingavariation oftheprocedure de-scribed by Stahl et al. (25).
X+D
phages
withsusS or susR mutations were grown in cells
containing partially deleted
prophages
fromwhich
S'
or R+ could be rescued (Fig. 1). Thesimultaneousrescueof the
X°
allele fromagiven
prophage indicated that the deletion in that
prophage had not removed the
XD
locus.Table 1 shows that the
XD
locus is locatedbetweenthe endpointsofdeletions MS1153 (or MS1118)andMS509.X°wasrescuedfrom
dele-tions MS5061, MS1118, and MS1153, but not
fromMS509, whether
S'
or R+ was the selectedrescuedallele. Similar results wereobtained in
crosseswith
X+D123
andX+D124,
whichsuggestthatthese two
X+
mutations arelocatedatthesame site or neareach other. (The nucleotide
sequenceanalysis reported below demonstrated
that these two mutations occurred at the same basepair.) These results did not,however, allow
us toconclude whether
XD
islocatedinQ, in S,orbetweenthe two genes.
Tolocate the
XD
locuson thephysical map ofA,
weutilizedplasmidscontaining fragments
ofTABLE 1. Rescue ofXDfrom partially deleted prophagesa
Pro- Fraction of
Chi'
among follow-phagepaeDeletion
Deletion endsends ingselectedrecombinants:dele-
betweenb:
S+tion bewen
strain
X+D123
X+D124
WM=MS5061 Q21andQts7 74/141 317/469 189/347
MS1118 Qts7and S3 176/310 280/419 72/119 MS1153 Qts7 and S3 67/117 409/592 116/182 MS509 S3and S2 0/370 0/448 0/111
aX+DsusSorX+DsusRphagesweregrown intheindicated bacterialstrains asdescribedinthetext. Sus+recombinants wereselectedon strain 594 andscoredbyaninspectionof the plaquesize for the Chi phenotype (10). The data indicate the number ofChio(small-plaque phenotype) recombinantsout of thetotalnumber ofrecombinantsinspected. Representative plaques from eachcrosswererestreakedonfreshplates to verifythe Chiphenotype.Complete genotypes of the infecting phagewereint4red3gam210X+D123susS7,b1453X+D123 susR5,and b2red3gam210 cI857X+D124susS7.
'Fromreference11.
A DNA from the Q-S region (Fig. 1). The
end-points of these fragments were generated by
endonucleases whose cleavage sites have been
located on thephysical map (6, 16, 18, 27). The
rescue of the X° allele from these fragments
wouldlocate
XD
relative to thesecleavage sites.This rescue was accomplished similarly to the
rescue from theprophages, via linkage to a
se-lectable marker (susP+, susS+,orsusR+).
Table 2 showsthat
XD
is located very nearEcoRI site 5 and to therightof it.AmongSusR+
phages generated by recombination with
plas-mid pRK112, 12% have rescued
X0.
This resultshows that
XD
is located on the3,600-base-pairfragment A2 (18) generated by endonucleases
HindIII and HindII. This fragment contains
EcoRI site 5 (18, 27). We crossed
X+D
Sus-phages with the twoEcoRI-generated fragments
extending to the left (pOPII-APQ) and to the
right (pDW1) of this site. No X° recombinants
were found among 5,000 P+ recombinants
gen-erated by recombination with plasmid
pOPII-APQ. On the otherhand, asmall number ofX°
recombinants were found withplasmid pDW1.
Theseresults show that
XD
islocated on EcoRIfragmentF(27), whichextendstotherightfrom
EcoRI site 5. Taken together, the results in
Table 2 show that
XD
isbetweenthe EcoRI siteand the HindII site located about 2,800 base
pairs to its right. From the lowfrequency of X°
rescue from plasmid pDW1 and the high
fre-quency of
X0
rescuefrom plasmid pRK112, weconcludedthat
XD
islocated very near therightof EcoRI site5.
Nucleotide sequence of the
XD
locus. To determine the nucleotide sequence of theXD
locus, weprepared DNA labeledatEcoRIsite 5andextending rightwardabout 350 basepairsto an MspI site. This fragment contains the
regiondeducedabovetocontainthe
XD
locus.Figure 2 shows the results of nucleotide
se-quenceanalysis from
wild-type
(X°),X+D123,
andX+D124
DNAs. The sequences obtained are identicalfor all threeDNAs,except forthe 55th nucleotide to the right of the EcoRI cleavageI
site(5'G-A-A-T-T-C 3'; measured on the I
strand). In wild-type DNA (AcI60 susS7), this
nucleotide isG, whereas in
X+D123
andX+D124
DNAs, thisnucleotide is T. Sequence analysis
of another wild-type
(X0)
strain (AcI72) showeda Gatnucleotide 55 (data not shown). In
addi-tion, ourwild-type sequence in this region agreed
with thatdetermined by F. Sanger and A.
Coul-son for their strain of AcI857 susS7 (personal
communication). We concluded that the X+D
mutation is the
change
G -- T 55 nucleotidesfrom the EcoRI site on the Istrand.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SEQUENCE OF x+D RECOMBINATIONAL HOT SPOT 339
A G C+T C A G C+T C
6-a
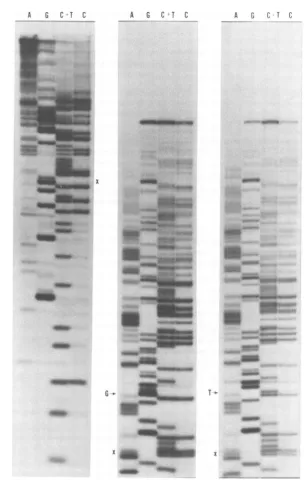
FIG. 2. Nucleotidesequencedetermination. (Left)
X'D124
sequenceextendingabout40nucleotidesright-wardfromtheEcoRI site shown in the bottom lineofFig.1.Identicalsequenceswereobtainedwithwild-type (X°)and
X'D123
DNAs(datanotshown).(Middle)Wild-type (X°) sequence extending rightward fromnucleotide 38toabout nucleotide85, relativetothe EcoRIcleavagesite.(Right)X+D124
sequencecorrespondingtothewild-typesequence in the middlepanel.Anidenticalsequencewasobtained with
X+D123
DNA(datanot shown). Arrows indicate thepointoftheX+Dmutation. ThesequencedeterminationwasbythemethodofMaxam andGilbert(13, 14).Cleavageatspecificbases isindicatedatthetopofeachlane. Fortheleft panel,
cleavedsamplesweresubjectedtoelectrophoresis througha20%polyacrylamide geluntilxylene
cyanol
FF(designatedX) hadmigrated11.5cm.Forthe middle andrightpanels,electrophoresis wasthrougha12% polyacrylamidegeluntilxylenecyanolFF hadmigrated34cm.Migrationwasfromtoptobottom. Further detailsonthepreparationofthelabeled DNA andonthesequencedeterminationaregiveninthetextand
references cited there.
x
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.496.84.393.68.549.2]340 SMITH ET AL.
TABLE 2. RescueofXDfrom plasmids carrying fragmentsof ADNAa
Re-Bacte- Extent of A bin- Fraction rial Plasmid DNAfragment' ants ofChi°
strain
se-lected FB1034 pRK112 HindIII-HindII R+ 51/432 Q pOPII-XPQ EcoRI-EcoRI P+ 0/5,000
D264 pDW1 EcoRI-cosc S+ 3/500
D264 pDW1 EcoRI-cos R+ 4/4,000
aX+D123phagewithsusP, susS,orsusR mutationswere
[image:5.496.64.250.74.190.2]grown in theindicatedbacteria, andSus+ recombinants were selected and scored for the Chiphenotypeas described in Table 1, footnote a. Complete genotypes of the infecting phage wereint4 red3 gam210X+D123susS7,b1453X+D123susR5, and red3 gam210 cI26susP80X+D123.Incontrolcrosseswith
Xe
phage (red3 gam210 cI26 susS7 or int4 red3 gam210susP80), noChi+ (large-plaque)Sus+ recombinants were found (data not shown),indicating that the X DNA fragments in the plasmids did not contain aX+Dmutation.
'SeeFig. 1.
cTherightcohesive end of X.
CreationofanAluI
cleavage
siteby
theX+D
mutation. The nucleotide sequencesre-ported above predict that the
X+D
mutationcreates arecognitionsequenceforendonuclease
AluI, d(5' A-G-C-T 3') (17). We verified this prediction bydigestion oftheend-labeledDNA fragments used for sequence
analysis.
Figure 3shows that digestion of the
X+D123
fragmenrtwith AluI produces a labeled
fragment
of the sizeexpected fromtheposition
oftheAluI rec-ognitionsequencerelative tothe other endonu-cleaserecognitionsites inFig. 1. Thewild-type(X0)
fragment
is resistanttodigestion
with AluIunder the sameconditions. We concluded that
the
X+D
mutations, like theX+C
mutations andsome
X+B
mutations, create an AluI recognitionsite (22, 23).
DISCUSSION
The resultspresented heredemonstrate that
X+D
mutationsaresingle-base-pair changes,
likethe
X+B
andX+C
mutations analyzed previously.The active
X+D
hot spot is created by thetrans-version G -- T (Fig. 1).
X+B
can be createdeitherby the tranversion C-+ G or bydeletion
of this C (22), whereas
X+C
is created by thetransversion A T
(23).
TheX'
mutationschange sequences in A which are almost the Chi
sequence into the correct, active Chisequence.
Acomparison of the nucleotidesequences at
the
X+B,
X+C,
andX+D
loci sheds somelight onthe sequence which defines Chi. As shown in
Fig. 4, the octamer 5' G-C-T-G-G-T-G-G 3' is
present at all three loci.When the twopossible
activeChi sequences at the
XB
locus are takenintoaccount, this octamer is the longest
contin-uous nucleotide sequence present at all three loci.At eachlocus, the
X+
mutation changesabaseto create this octamer(Fig. 4). If Chi isa
unique nucleotide sequence, this octamer may well be Chi. Two other
possibilities
must be considered, however. Perhaps Chi is a uniquesequence, but onlya portion ofthe octamer is
Chi;orperhaps Chiis avariablesequencewhich includes the octamer.
The extent and precise nucleotide sequence
requirement of Chi can be determined via an
examination of mutations which create or
de-stroy anactive Chisequence. Inthe
alignment
ofsequencesshown inFig.4, wecould conclude
thattwo nucleotides are essential to Chi.
X+B
mutationslead to aGatthe leftmostpositioninthe octamer underlined in Fig. 4. A C at this
position abolishes Chi activity (22).
X+C
andX+D
mutations leadtoaTatthethirdposition
in the octamer. An Aor a G at this
position
isFIG. 3. Creation ofanAluIcleavage site by
X'D
mutations. DNAfr-agmentslabeledatthe EcoRI site shown in the bottom line ofFig. 1 and extending
rightwardto thefirst MspI cleavagesite(about350
base pairs) were digested with endonuclease Alul.
Thedigestwassubjectedto electrophoresisthrough
an8%polyacrylamide gelinEbuffer (20). Migration
wasfr-om top to bottom. (a)
X'D123
DNA; (b)wild-type (X") DNA. Arrows indicate thepositionsofthe
origin (0), migration of xylene cyanolFF (X9, and
migration of bromophenolblue(B). Thepositionsof
labeled
fr-agments
produced by digestionwithendo-nuclease HhaI (arrow 1) and with endonuclease HaeIII(arrow 2)weredetermined ina separate
ex-periment underidentical conditions by noting their
migrationrelative to thedyes. The relative sizes of
thesefr-agmentsareshowninFig.1.Asmallamount
oflabeledDNAfr-om
X'D123
ramainedundigested. J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.331.391.297.486.2]X+D
X B121 5'ACAATGAGTGGCAGATATAGGCTGGTGGTTCAGGCGGCGCATTTTTAT 3' X B132 5'AACAATGAGTGGCAGATATAGCTGGTGGTTCAGGCGGCGCATTTTTAT 3' XC157 5'CGTTGATAAGTCGCAGATCAGCTGGTGGAAGAGGGACTGGATTCCAAA 3' XVD123 5'TACACATGACCTTCGTGAAAGCTGGTGGCAGGAGGTCGCGCTAACAAC 3'
FIG. 4. Comparison of nucleotide sequences at the X+B, x+C, and X+D loci. Portions of the I-strand
sequencessurrounding the indicatedX+mutations are shown. Two different sequences, exemplified by the
X+B121
andX+B132
mutations, are found at theX+Blocus(22). TheX+C
sequence isfromreference 23, and theX+Dsequence isfrom Fig.1and 2.Thesequences are aligned such that theunderlined octamer is coincident for the four sequences (seetext).Hyphens are omitted for clarity.
ineffective (23; Fig. 2). These observations are
consistent withtheview thatChiis, orcontains,
the uniqueoctamer indicated in Fig. 4. Verifi-cation of thishypothesis could be achieved by analysis ofmoremutationswhich createChior by the isolation andanalysisofmutations which destroy Chi. If Chiislimitedtothe octamer,all ofthese base changes would occur within the octamer.
It is interesting that the octameris oriented in thesame directionatthe three
X+
loci pres-ently analyzed. That is, the octamer shown ispresent on the I strand of A atall three active
Chisites. Fauldsetal. (8) have shown thatthe
inversion of ChiwithinA inactivates Chi. Yagil et al. (E. Yagil, N. Dower, D.
Chattoraj,
M.Stahl,
C. Pierson, andF. W.Stahl, Genetics,
inpress)haveshown thatthe active orientation of
the Chi site that they used, resulting from a
spontaneousmutationin thetransposonTn5,is
the samethroughout the right 60%ofA.Ifthe
octamerisatleastpart of
Chi,
wewouldantici-patethat it would be present in the same ori-entation ateach
position
inA,
as wehave ob-served.ACKNOWVLEDGMENTS
We areparticularlyindebtedtoJeffrey Roberts,Michael
Schechtman,DavidWilson,and William Reznikoff forsending
ustheirplasmidscontainingfragmentsof ADNAand infor-mation about them beforepublication.Wealso thank Jean
Crasemann, JeffreyRoberts,MaryStahl,and Frank Stahl for phage and bacterial strains from their collectionsandFred
Sanger and Alan Coulson for unpublished sequences. We
gratefullyacknowledgeourmanycolleaguesintheInstitute ofMolecularBiologyfor manyhelpfuldiscussions and unpub-lished data.
This research was supported by Public Health Service grantsAI13079andGM26798from the National Institutes of HealthtoG.R.S. and Public Health Service grant GM21812 from theNational Institutes of HealthtoF.R.B.
LITERATURE CITED
1. Berg,D. E. 1977. Insertion and excision of the transpos-ablekanamycinresistance determinantTn5,p.205-212. In A.I.Bukhari,J. A.Shapiro,andS.L.Adhya (ed.),
DNAinsertionelements,plasmids,andepisomes.Cold SpringHarborLaboratory,ColdSpringHarbor, N.Y. 2. Berkner,K.L.,and W. R.Folk. 1977.Polynucleotide
kinase exchange reaction. J. Biol. Chem. 252:3176-3184.
3. Boyer,H.W.,M.C.Betlach, F. Bolivar,R.L.
Rodi-guez,H.L.Heyneker, J. Shine, and H. M. Good-man. 1977. Theconstruction ofmolecular cloning ve-hicles, p. 9. In R. F. Beer and E. G. Bassett (ed.), Recombinant molecules: impact on science and society. Raven Press, New York.
4.Campbell,A. 1961.Sensitivemutantsofbacteriophage A.Virology 14:22-32.
5. Court, D., and K. Sato. 1969. Studies of novel transduc-ing variants of lambda: dispensability of genes N and Q. Virology39:348-352.
6. Daniels, D. L., J. R. de Wet, and F. R. Blattner. 1980. New map ofbacteriophage lambda DNA. J. Virol. 33: 390-400.
7.Davidson, N., and W. Szybalski. 1971. Physical and chemicalcharacteristics of lambda DNA, p. 45-82. In A. D.Hershey (ed.), The bacteriophage lambda. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. 8. Faulds, D. H., N. Dower, M. M. Stahl, and F. W. Stahl.
1979.Orientation-dependent hotspot activity in phage A. J. Mol. Biol. 131:681-695.
9. Goldberg,A.R., and M. Howe. 1969. New mutations in the S cistron ofbacteriophage A affecting hostcell lysis. Virology 38:200-202.
10. Henderson, D.,andJ.Weil. 1975. Recombination-defi-cient deletions inbacteriophageAandtheirinteraction withchi mutations. Genetics 79:143-174.
11.Herskowitz, I., and E. R. Signer. 1970. A site essential forexpression of all late genes inbacteriophage A. J. Mol. Biol. 47:545-556.
12.Malone, R. E., D. K. Chattoraj, D. H. Faulds, M. M. Stahl, and F. W. Stahl. 1978. Hotspots for generalized recombinationin theEscherichia coli chromosome. J. Mol. Biol.121:473-491.
13.Maxam, A. M., and W. Gilbert 1977.Anewmethod for sequencing DNA. Proc. Natl. Acad. Sci. U.S.A. 74:560-564.
14.Maxam,A.M.,and W.Gilbert.1980.Sequencing end-labeled DNA with base-specific chemical cleavages. MethodsEnzymol. 65:499-560.
15.Muller-Hill,B.,L.Crapo,andW.Gilbert. 1968. Mu-tantsthatmake morelacrepressor.Proc. Natl. Acad. Sci. U.S.A. 59:1259-1264.
16. Murray,K., and N.Murray.1975.Phagelambda recep-torchromosomes for DNAfragments made with restric-tionendonuclease III ofHaemophilusinfluenzaeand restriction endonuclease I of Escherichia coli.J.Mol. Biol. 98:551-564.
17. Roberts, R. J., P. A. Myers, A.Morrison, and K. Murray. 1976. Aspecific endonuclease from Arthro-bacter luteus. J. Mol.Biol.102:157-165.
18. Robinson,L H.,and A.Landy.1977.HindII, HindIII,
andHpaIrestrictionfragment mapsofbacteriophage A DNA. Gene 2:1-31.
19. Shapiro,J.,and S.Adhya.1969.Thegalactoseoperon ofE.coliK-12.II. A deletionanalysisofoperon struc-tureandpolarity. Genetics62:249-264.
20. Sharp, P. A., B. Sugden, and S. Sambrook. 1973. Detection of two restriction endonuclease activities in Haemophilusparainfluenzae using analyticalagarose
on November 10, 2019 by guest
http://jvi.asm.org/
342 SMITH ET AL.
ethidium bromide electrophoresis. Biochemistry 12: 3055-3063.
21. Silverstone,A.E.,R.R.Arditti,and B.Magasanik. 1970.Catabolite insensitiverevertantsofLacpromoter mutants.Proc.Natl. Acad. Sci. U.S.A.66:773-779. 22. Smith,G.R.,D.W.Schultz, and J.M.Crasemann.
1980.Generalizedrecombination: nucleotidesequence
homology between Chi recombinational hotspots.Cell 19:785-793.
23. Sprague, K. U., D. H. Faulds, and G. R. Smith.1978. Asingle base-pair changes creates aChi
recombina-tionalhotspotinbacteriophageA.Proc.Natl. Acad. Sci.
U.S.A. 75:6182-6186.
24. Stahl,F. W. 1979.Special sitesingeneralized
recombi-nation.Ann.Rev. Genet.13:7-24.
25. Stahl, F. W., J. M. Crasemann, andM. M. Stahl.1975. Rec-mediated recombinational hotspotactivity in bac-teriophagelambda. III. Chimutationsare
site-muta-tions stimulatingrec-mediated recombination.J. Mol. Biol. 94:203-212.
26. Sutcliffe, J. G. 1979.Completenucleotide sequenceof theEscherichia coli plasmid pBR322.ColdSpring Har-bor Symp. Quant.Biol.43:77-90.
27. Thomas, M.,and R. W. Davis. 1975. Studies onthe cleavage of bacteriophage lambda DNA with EcoRI restriction endonuclease. J. Mol. Biol.91:315-328. 28. Weigle, J.1966.Assembly of phage lambdain vitro. Proc.
Natl. Acad. Sci. U.S.A.55:1462-1466.
J. VIROL.