Copyright 01973 AmericanSocietyforMicrobiology Printed inU.SA.
Prophage
Induction
by High Temperature
in
Thermosensitive dna
Mutants
Lysogenic
for
Bacteriophage Lambda
H. SCHUSTER, D. BEYERSMANN, M. MIKOLAJCZYK, AND M. SCHLICHT Max-Planck-Institut fiirMolekulare Genetik, 1Berlin 33(Dahlem), Germany
Received forpublication 14December 1972
High-temperature treatment of thermosensitive dna mutants lysogenic for
phage A leads to prophage induction and release of phage (at the permissive
temperature) in elongation-defective mutantsof thegenotypesdnaB,dnaE, and
dnaG. In initiation-defective mutants no prophage inductionoccurs at42 C in mutants ofthe genotype dnaA, whereas with a dnaC mutant as well as with strain HfrH 252 (map positionnotyetknown) phagesarereleased at 42C. DNA degradation atthe replication fork at 42 C is observed in all dnaB(A) mutants
tested, but not in mutants ofthe genotypes dnaE(X) and dnaG(A). Therefore,
degradation of replication fork DNA is not a prerequisite for prophage induction.
In
X-lysogenic bacteria, phage development
canbe induced
by
a variety oftreatments thatdamage the bacterial chromosome and inhibit
DNAreplication. Since the bacterial recA
func-tionisrequired for induction (3), it issupposed
that induction results from recA-mediated
re-pair of
damaged
DNA. The mechanism ofinduction isstill obscure (5).
In thermosensitive dna mutants, DNA
re-plication can be interrupted at high
tempera-ture (42 C and
above),
and the mode ofcessa-tion of DNA synthesis varies markedly in the
different classesofdnamutants.In most ofthe
mutant types known to be defective in chain
elongation
(dnaB, dnaE, dnaG),
DNAsynthesisstops
immediately
after reaching the criticaltemperature (42 C; reference 23). A residual
DNA synthesis at the elevated temperature is
observedin mutant typesdefective inthe
proc-essof initiation (dnaA, dnaC, strain HfrH 252;
1, 4, 12). In dnaBmutantsDNAatthe
replica-tionfork is
degraded
extensively at 42C (19).It seemed worthwhileto us toexamine
X-lyso-genic dna mutants of different genotypes for
their ability to induce the prophage at high temperature inorder to find out whether or not
a damaged bacterial DNA is a necessary
pre-requisite for induction to occur. It hasalready
been reported that prophage A is not induced
when DNA synthesis ceases at 42 C in an
Escherichia colidnaA mutant. However,
induc-tionoccurredinthis mutant when the cellswere
UV-irradiated after
completion
of rounds ofreplication at 42 C (20). On the other
hand,
ithas been found that
prophage
X is induced athigh temperature in bacterial mutants with
temperature-sensitive
but unknowngenetic
de-fects (15,
22).
Induction ofprophage
Xhas alsobeen inferred from the loss of
colony-forming
ability
of a dnaB mutant after temperatureshifts to 42C (21).
MATERIALS AND METHODS
Radioisotopes. Radioisotopesusedwere 3H-thymi-dine,22 to 27Ci/mmol, and3H-thymine,5Ci/mmol,
from The Radiochemical Centre, Amersham,
Eng-land.
Bacteria. The E. coli strains listed in Table1 were used. All strains were lysogenized by phage A (wild
type) in our laboratory. In addition, strain HfrH
165/70 was lysogenized by phage Aind-. Strain K-12 C600wasusedas indicator for A.
Media. Tryptone broth: 1% tryptone (Difco) and 0.5% NaCl. K medium: 1: 1 mixture of
double-strength M9 medium and 3% Difco Casamino Acids (27); A medium (7); M9 medium (27). All synthetic mediaweresupplementedwith20
Mig
ofthymine and thiamineperml.Inthe case of strain HfrH252, the media contained 20Mlg
ofproline per ml in addition.Measurement of induction of prophage X by temperature shift.Lambda-lysogenic bacteria grown
overnightintryptone broth at room temperature were usually diluted1:1,000 into K medium and grown at 30Ctoabout1 x 101cells per ml. The temperature of theculturewasthen raisedto 42C (or46Cinthe case of dnaE mutants) and incubation was continued. 879
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Escherichia coli strainsa
E. coli strain dna Remarks Reference
locus
HfrH 252 ? 1,17
HfrH 252rev dna+ Selected for growth This paper at 42C, our
lab-oratory
PC5 A 4,23
E508 A 23
CRT46 A 13, 23
HfrH 165 dna+ Parental strain of 2, 19 HfrH165/70
HfrH165/70rev dna+ Selected for growth Thispaper at 42C, our
lab-oratory
HfrH165/70 B 2,19
Hfr 100 B 19
PC6 B 4, 11
PC8 B 4,11
PC2 C 4,23
PC7 D 4,23
BT1126 E 24
E486 E 23
E101 F 9,23
JW185 G dna allelenumber 11
308
PC3 G 4,23
aSource of dna mutants: HfrH 252, HfrH 165, HfrH
165/70, Hfr 100, BT1126 (H. Schaller and F. Bonhoeffer);
PC2, 3, 5, 6, 7, 8 (J. Zeuthen); E486, E101, JW185 (J.
Wechsler).
Since the time period for optimal induction at ele-vated temperatures might be different for different dna mutants,ananalysisasshowninFig.1and3 was carriedoutwithatleastone mutantofeach genotype. For this purposesampleswerewithdrawnatintervals and (i) assayed immediately for surviving bacteria (colony-forming ability at 30 C), induced prophage,
and free phage (after treatment with CHCl3); (ii) diluted 1: 100 into K medium and incubation
con-tinuedfor 2 to 3hat30C,after which theculturewas
againassayed for freephage.
In someexperiments, K medium was replaced by
tryptonebroth,M9or Amedium.Thenumber of cells containingan inducedprophageweremeasuredbya
spottestin ordernot tooverlook induction processes yielding only low phage bursts. A serial dilution of bacteria(in the rangeof 50to2,000 cells perspot)was
spottedonto alawnofK-12 C600 indicator bacteria. Plates were subsequently incubated at 27 to 30 C. Control cells(grownat30C)wereincludedinthespot test to correct for the spontaneously induced pro-phages.
Measurement of the stability of radioactively
labeled chromosomal DNA at elevated tempera-ture:(i)pulselabeling(replicationforkDNA).To1
volume oflog-phase grownbacteria in K medium at
30C,0.03volume of3H-thymidine(1mCiperml)was
addedfor50s,after which theculturewastransferred
to 42 C (or 46C in thecase ofdnaEmutants) with
simultaneous addition of 0.05 volume ofthymidine
(10 mg/ml).
(ii) Uniform labeling (preexisting DNA).
Bac-teriawere grownfor sixtosevengenerationsat30C in
K medium supplemented with 3H-thymine (10 uCi and 20mg perml). Ata titer of108perml, the cells
were harvested by filtration, washed, and resus-pended in the same volume of K medium
supple-mented with 200 ug ofunlabeled thymine. After an additional 15 minofincubationat30 C,the cultures
weredivided. Onepartwasshiftedto42C (or46C in
thecaseofdnaE mutants), and anotherpartwaskept at30C.
Samples (0.1 ml) wereremoved successively from
theradioactive cultures.Incorporationofradioactive DNA precursors wasterminated, and trichloroacetic acid-insoluble radioactivity was determined as de-scribed elsewhere (19).
RESULTS
Induction bytemperatureshift of prophage
X in lysogenic dna mutants: (i) induction of
X-lysogenic initiation-defective mutants. In initiation-defective mutants, X is able to repli-cate at temperatures at which host DNA syn-thesis hascome toahalt (13, 17). Here, induc-tion of prophage and release of phages can be followed directly at the elevated temperature.
The initiation-defective mutants dnaA E508(X) and dnaA CRT46(X) do not undergo
spontaneous prophage induction after cessation of DNA synthesis at 42 C (20). Another dnaA
mutant (PC5), isolated by Carl (4), also does
not induce X prophage efficiently. When this
strain is incubated for2to3hat42C, at most
4% ofthe cellsare induced(Table 2).
We would conclude from theseresults that in initiation-defective mutants cessation of host DNA synthesis perse is not a sufficient condi-tion for X induction to occur. However, this is
not true for an initiation-defective mutant of the genotype dnaC PC2 (4). With the nonlyso-genic strain, thecell numberstillincreasesbya factorof3.3reachingaplateau after2to3 hat 42C. Under thesameconditionstheX-lysogenic mutantinduces the prophageand the cells die
(Table 2).
Still another initiation-defective mutant,
strain HfrH 252(X) (whose map location is not yet known), behaves similarly to the strain
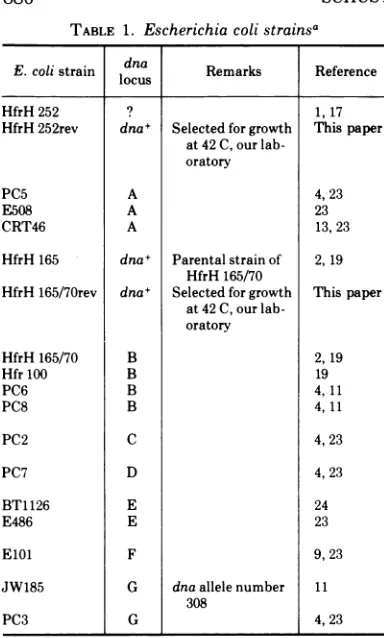
dnaC PC2. In Fig. 1 phage release at 42 C is
followedover aperiodof 4 h in A medium for the
mutantaswellasforarevertantof thismutant.
In both strains, the number of spontaneously induced phages increases as a consequence of
thetemperatureshiftto42C.Theeffect ismore pronounced with the mutant than with the revertant strain. However, no cell death is
880
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 2. Induction of prophage Xindna(X) mutants byhigh temperaturea dna(X)mutant dna locus Induction Fractionofcells induced Remarks
HfrH 252 ? + In A medium:seeFig.1
In K medium: phage bursts >2 after 3 h at 42 C (surviving cell fraction = 0.15)
PC5 A _ <0.04(2-3 h42C)
E508 _ } Measured by Monk and Gross (20)
CRT46
HfrH165/70 B + 0.4-0.7(2-3h42C)
Hfr 100 + Maximal phage release after 1-2 h
PC6 + at42C+3hat 30C (seeFig. 2)
PC8 +
PC2 C + Phage bursts >2after 3h at42C
(surviving cell fraction = 0.03)
PC7 D _ Slight increase of spontaneously
induced prophages at 42 C
BT1126 E + 0.1-0.4(1.5 h 46C) Phage bursts usually <1
E486 + 0.7-0.9(1.5-2h46C) Maximal phage release after 1.5 h at46C + 3hat 30C
Ell F _
JW185 G + 0.3(3 h42C) Maximalphage release after3h
at42C+3hat30C
PC3 0.005(2 h42C)
aInduction isregardedaspositive,whenatleast10%ofthemutantcellsyieldinfectivecenters.The fraction ofcellsinduced is difficult to determineexactlywith the strains dnaCPC2(X) and HfrH252(X),sinceinthese mutants A isable to replicate at42C. Here phage burstsareexpressedasthe number ofphagereleasedat42C perviable cell at time -zero, 42 C.
102 [
101I
1.
* 10
@ 10 2[
A
bacteria
/ phage
A
B
102
-1o1l bacteria _._
-,/ phage
0-3-~ -0_ _
_I I
0 1 2 3 4 0 1 2 3 4
hours at 42 °C
FIG. 1. Phage release at 42 C of the strains HfrH252(X) and HfrH252rev(X). Bacteria were grown in A medium at 30 C andincubated in the same medium at 42 C. Cell concentration at time zero, 42 C: A = HfrH
252(A),5 x 107per ml;B = HfrH252rev(A),1 x 108 perml. .1_
0o-3
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.106.399.433.638.2]observed with strain HfrH 252 afterDNA
syn-thesis and cell divisions have come to a halt,
indicating that only a minor fraction of cells
have been induced. Only the subsequent
ir-radiation with UV light or treatment with
mitomycin (both at 42 C) leads to induction
(data not shown). However, when K medium
wasused instead of Amedium, the cells diedat
42 C and a larger number of phages were
released (Table 2).
(ii) Induction of X-lysogenic
elongation-defectivemutants. In the
elongation-defective
mutants investigated, X DNA is not
replicated
atthe restrictivetemperature (6, 13, 17).
There-fore, induction of
prophage,
provoked
by
theinterruption of DNA
synthesis,
issubsequently
measuredattemperatures
permissive
for DNAsynthesis. Phage bursts of
prophage-induced
bacteria are
determined
at 30 C after theinduction
period
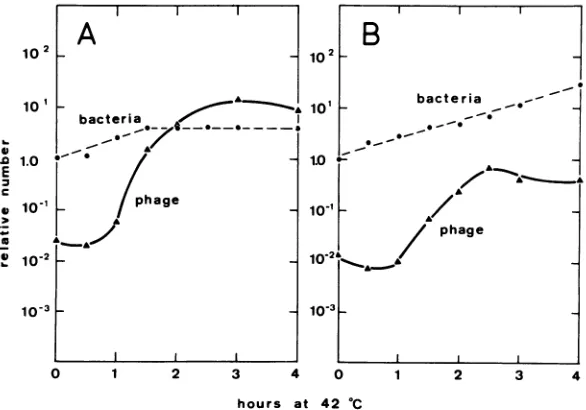
at 42 C.dnaBmutants. In
Fig.
2phage
release at 30C of strain HfrH
165/70(X)
after an inductionperiod
of 100 min at 42C is shown. Amaximalburstisobtained after3to4hofincubation at
30 C. The
phage
yield (after
3 h at 30C)
inrelationtothe
preceding
inductionperiod
at42108
io7
._
106
105
0 1 2 3 4
[image:4.493.59.247.347.590.2]hours at 30 °C
FIG. 2. Phage release at 30 C of strain HfrH 165/70(X) after 100 min of preincubation at 42 C. Bacteria were grown in K medium at 30 C and incubated in thesamemediumat42C. Cell concen-trationattimezero,42 C: 9x101perml.The culture
was diluted1:100 into K mediumafter100minand incubationwascontinuedat30 C.
C is shown in Fig. 3A. Largest bursts are
obtainedafter 1 to 2 h ofincubation at 42 C, and
induction is accompanied by a decrease in the
numberofsurviving bacteria. On the contrary,
intherevertant strain, growth at 42 C only leads
to aslight increase in the numberof
spontane-ously induced phages (Fig. 3B).
According to the experimental conditions
outlined in Fig. 3, all dnaB mutants examined
induce X prophage very efficiently following an
incubation periodat 42 C of 1 to 2 h (Table 2).
However,different dnaBmutants differ slightly
with respect to the length of time at 42 C
required for optimal induction. It is also
com-mon tothe (lysogenicaswell as the nonlysogen-ic) dnaB mutants tested that at 42 C they
degrade their DNAatthereplication fork
exten-sively (see below andreference 19).
Prophage induction studies were done in
moredetail withstrainHfrH165/70(X).Usually
40 to70% of thecells induced the prophageat 42
C. Induction and phage release in K medium
werealsoveryefficientathighconcentrations of
bacteria (omitting the 1: 100 dilution step, see
Materialsand Methods). By this means phage
titerslarger than1 x 1010permlwereobtained.
Prophage induction also occurred when K
me-diumwasreplaced bytryptonebroth. However,
forunknownreasonsphageyieldwasextremely
low if the 1:100 dilution step (into tryptone
broth)
was omitted. In M9 medium, phagebursts lower than0.1 wereobtained after50 min
at 42C and2h at 30C. ProphageXind-, which
is not inducible by UV (14), also cannot be
induced by high temperature in
165/70(Xind-).
dnaE mutants. In mutants having a
ther-mosensitive lesion at the dnaE locus, DNA
polymerase III activity is thermosensitive (10).
Inthe strains E486 andBT1126, DNAsynthesis
stops at 42 C and 46 C, respectively. No DNA
degradation at the replication fork is observed
at these temperatures (see below). Both
lyso-genicstrains induce their prophage (Table 2).
dnaG mutants. In the strains JW185 and
PC3,
the synthesis of short deoxynucleotidechains (Okazaki DNA) is inhibited at 42 C (18). At that temperature nodegradationofDNA at
thereplicationfork isobserved in strain JW185
(see belowandFig.4).Strain
JW185(X)
inducesthe prophage at 42 C; strain PC3(X) does not
(Table2). UV irradiation of the bacteria at
30
Cleads to induction of
JW185(X),
butPC3(X)
isonly poorly induced. As it was found later,
phageXisonly weakly reproducedinPC3at 30
C (in comparison to growth in C600). In this
mutant, either the mutation in dnaG affects
growthof X at 30C orthepoorgrowthisdue to
someotherproperty ofthe strain.
io4
103,
on November 10, 2019 by guest
http://jvi.asm.org/
102 1
10'
1.0 1
10o-lo03
0 1 2 3 4 0 1 2 3 4
hours at 42 C
FIG. 3. Phagereleaseat30 Caftervariouspreincubation periodsat42C. BacteriaweregrowninKmedium
at30C and incubatedin thesamemediumat42C. Cell concentrationattimezero,42C. A = HfrH 165/70(X),8
x 107per ml;B = HfrH 165/70rev(X), 7x 105per ml. Relative numberofbacteria (----0)andrespective
phage (A A) afterthe indicated timeat42C. Relative numberof phage(A A)afterthe indicated time
at42Cplusanadditional timeof3 hat30 C in eachcase.
Stabilityof chromosomal DNA and X
pro-phage induction in dna mutants: (i) fate of
30 C pulse-labeled DNA atelevated
temper-atures. With X-lysogenic as well as with some
nonlysogenic elongation-defective mutants, the
effectofhightemperatureon(i) thereplication
fork DNA(the mostrecently made DNA at 30
C) and (ii) the uniformly labeled chromosomal
DNAwas studied. In the formercase theDNA
was pulse-labeled with 3H-thymidine
immedi-ately before the temperature shift (see
Mate-rials and Methods). Aswas already mentioned
above, all the dnaB mutantstested (see Table
2) degrade replication fork DNA. As an
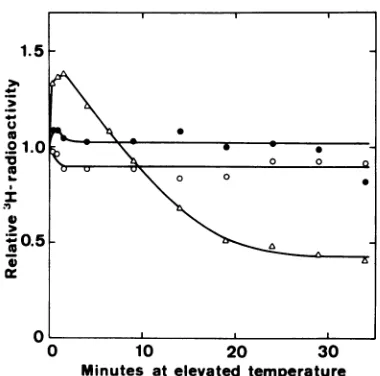
exam-ple, the kinetics of DNA degradation in strain
dnaB PC8(X) are shown in Fig. 4. However,
when mutants ofthe genotype dnaE [E486(X)
and BT1126] and dnaG [JW185(X) and PC3]
were tested in the same way, no significant
degradation was observed. The stability of
pulse-labeledDNAat42C is showninFig.4for
the strains E486(A) andJW185(A). Both these
mutants, aswellasstrainBT1126(X), do induce theirprophage by high temperature treatment
(Table 2).Obviously degradationof the DNAat
the replication fork is not a prerequisite for
inductiontooccur.
(ii) Fate of 30 Cuniformly labeledDNAat 42 C. Elongation-defective mutants with
3H-thymine uniformly labeled DNA were
in-cubated in K medium at30C and42C for 3h,
and trichloroacetic acid-insoluble radioactivity
was determined at 30-min time intervals (see 1.
I
.21.
0
cc
0 10 20 30
Minutes atelevated temperature
FIG. 4. Fate of 30 C pulse-labeled DNA in dna
mutants at elevated temperatures. Bacteria were
grownin Kmedium at 30 C and pulse labeled with 3H-thymidineat30C, and trichloroacetic acid-insolu-bleradioactivity wasfollowed aftertemperatureshift (see Materials andMethods).A,Strain dnaBPC8(A), 42 C;0, strain dnaEE486(X),46C; 0,strain dnaG
JW185(G),42 C.
also Materials and Methods). With the strains
dnaBHfrH165/70(X), dnaEE486(X), and dnaG
JW185(X), between 15 and20% of the 3H label
became acid-soluble after 3 h of incubation at
bothtemperatures. However, degradation at42
C wassomewhat faster thanat30C in all three
mutants. Under the same conditions in strain
4) .0
0
B
,
bacteria ,
_~~~~~~~~~.
0~0
4
F ,phage,
742°C.3h30°C/'
A
hage
/I
.5
0
.0 0
.0 0~
~~~~~
0o L
1.5
o
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.99.397.47.269.2] [image:5.493.254.444.323.511.2]dna+165(X) the acid-insoluble 3H label
in-creased by about5% at 30 C (probably due to a
depletion of the residual 3H-labeled precursor
pool).Theseresults indicate that in the mutant
strainsthednadefectalreadyleads to alimited
turnover of the chromosomal DNA at 30 C.
Surprisingly, in strain dna+165(X) 13% of the
DNA was also made trichloroaceticacidsoluble after 3 h at 42 C. Therefore, it is unlikely that
the DNA turnoverat 42C in thedna mutantsis
thereason forinduction.
DISCUSSION
Lambda-lysogenic elongation-defective
mu-tants of the genotype dnaB, dnaE, and dnaG
induce their prophage at high temperature.
Thus, the inactivationof afunctionresponsible
forthe elongationof areplicating DNA has the
same effect as the variety ofother treatments
whichall leadtoprophageinduction by
damag-ing the DNA and inhibiting DNA replication.
Consistent with this are the findings that dna
mutants with onlyareducedDNAsynthesisat
elevated temperature
(dnaD
PC7, residualDNA synthesis withoutreaching a plateau [4];
dnaF E101, DNA
synthesis
with animmediatereductioninrate [23] dueto adefective
ribonu-cleoside
diphosphate
reductase[9])
do notin-ducetheprophage (Table 2).
It is striking that all dnaB mutants which
degrade their DNAatthe
replication
forkat42C also induce their
prophage
andyield
largephage bursts. (In
comparison
to the inductionby UV irradiation this "temperature induction"
iseasier tohandle, andwe areusingit
routinely
for the preparation ofphage
stocks.)
However,these DNA
degradation
products
areprobably
not the reason for the release of
prophage
re-pression since other
elongation-defective
mu-tants
(dnaE,
dnaG)
which donotdegrade
theirreplication fork DNA arealso inducible
by high
temperature. It is also
unlikely
that a moregeneral
degradation or abreakdown andresyn-thesis ofpreexisting DNAat42C isthereason
forinduction.
Although
aturnoverofthis DNAisobserved at 42 C in dna mutants, it also takes
place in a lysogenic wild type which is not
induced atthistemperature.
When DNA
synthesis
isstopped by thymine
starvation in E. coli strainsauxotrophic.
forthymine, an endonuclease activity appears
which introduces nicks into the DNA(8). Since
X prophage can also be induced
by thymine
starvation (16),nickingofthe DNAmaytrigger
in some way the induction process. Such an
eventcould also be thereasonfortheinduction of X in dna mutants. However,
preliminary
experiments with a (nonlysogenic) dnaB
mu-tantruled out an extensive nicking of chromoso-mal DNA (19). A more careful analysis of all
lysogenic dna mutants tested is in progress.
Thatinduction should result from the action
of therecA-mediatedrepair ofdamaged DNAis
difficult to explain in the case of such
elonga-tion-defective dnamutants which do not show a
noticeable
DNA breakdown but neverthelessinduce their prophage (dnaE E486, dnaG
JW185). However, the involvement ofthe recA
product in prophage induction may have noth-ing to do with DNA repair processes. In
(non-lysogenic) dnaB mutants asymmetric
chromo-some reinitiations can be triggered by
tempo-rary blocks in DNA chain elongation (25).
Double mutants of the genotype dnaB recA,
however, fail to reinitiate chromosome replica-tion after temperature shifts (A. Worcel, M.
Schwartz, personal communication). Such
ab-normal initiations may also trigger prophage
induction. They have in fact been observed
during inductionof aBacillus subtilisprophage
(26). It has to be shown whether abnormal
chromosomal initiationscanbe observed alsoin
all inducibleelongation-defective dnamutants.
In the initiation-defective dnaA mutant
PC5(X),
theprophage isnotinducedat 42C. Aswasobserved earlier, cessation of DNA
synthe-sis at 42 C in two other X-lysogenic dnaA
mutants also did not lead to induction (20).
Comparing the inducibilityof
elongation-defec-tive dna mutants with the non-inducibility of
initiation-defective dnaA mutantsby
tempera-tureshifts,onecouldassumethat the
induction.
process is onlytriggered in such cells in which
the chromosomal DNA is in the process of
replication. However,sincetwoother
initiation-defectivemutants,namely dnaC PC2 and HfrH
252
(unmapped),
can induce theirprophage
at42C, the explanationcannotbeso
simple.
Thefactthat thetwolatter strainsbehave
similarly
with regard to induction may indicate that
strain HfrH252isalso adnaC mutanttype.
ACKNOWLEDGMENTS
We aremuch indebtedto F.Bonhoeffer,H. Schaller, J.A.
Wechsler, and J. Zeuthen for providing us with the dna mutants. We would like to thank H. A. Rolton for her assistance inthe correctionofthemanuscript.
LITERATURE CITED
1. Beyersmann, D., M. Schlicht, and H. Schuster. 1971. Temperature-sensitiveinitiationofDNAreplicationin a mutant ofEscherichia coli K12. Mol. Gen. Genet.
111:145-158.
2. Bonhoeffer, F. 1966. DNAtransfer and DNAsynthesis during bacterial conjugation. Z. Vererbungsl.
98:141-149.
884
on November 10, 2019 by guest
http://jvi.asm.org/
3. Brooks, K., andA.J. Clark.1967. Behavior ofA
bacterio-phage in a recombination deficient strain of Esche-richia coli. J. Virol. 1:283-293.
4. Carl, P. L. 1970. Escherichia colimutantswith tempera-ture-sensitive synthesis of DNA. Mol. Gen. Genet.
109:107-122.
5. Echols, H. 1971. Lysogeny: viral repression and
site-specific recombination. Annu. Rev. Biochem.
40:827-854.
6. Fangman, W. L., and A. Novick. 1968. Characterization of two bacterial mutants withtemperature-sensitive synthesis of DNA. Genetics 60:1-17.
7. Fox, E., and M. Meselson. 1963. Unequal photosensitiv-ityofthetwostrands of DNA inbacteriophageA.J.
Mol. Biol. 7:583-589.
8. Freifelder, D., and E. Levine. 1972. Stimulation of
nuclease activity by thymine starvation. Biochem. Biophys. Res. Commun. 46:1782-1787.
9. Fuchs, J. A., H. 0. Karlstr6m, H. R. Warner, and P.
Reichard.1972.Defectivegeneproductin dnaF mutant
of Escherichia coli. Nature N. Biol. 238:69-71.
10. Gefter, M. Y. Hirota, T. Kornberg, J. A.Wechsler,and C.
Barnoux. 1971. Analysis of DNA polymerasesIIandIII
inmutantsof Escherichia coli thermosensitivefor DNA
synthesis. Proc. Nat. Acad. Sci. U.S.A. 68:3150-3153.
11. Gross, J. D. 1972. DNA replication in bacteria. Curr. Top. Microbiol. Immunol. 57:39-74.
12. Hirota, Y., J. Mordoh, and F. Jacob. 1970. On theprocess ofcellular divisioninEscherichia coli.III.
Thermosen-sitivemutantsofEscherichia coli alteredintheprocess of DNA initiation.J. Mol. Biol. 53:369-387. 13. Hirota, Y., A. Ryter, and F. Jacob. 1968. Thermosensitive
mutants of E. coli affected in the process of DNA
synthesis and cellular division. Cold Spring Harbor
Symp. Quant.Biol.33:677-693.
14. Jacob, F., and A. Campbell. 1959. Surle systemede
repressionassurantl'immunite chez les bact6ries lyso-genes. C. R. Acad. Sci. 248:3219.
15. Kirby, E. P., F. Jacob, and D. A. Goldthwait. 1967.
Prophage induction and filament formation in a
mu-tantstrain of Escherichia coli. Proc. Nat. Acad. Sci. U.S.A. 58:1903-1910.
16. Korn, D., and A. Weissbach. 1962.Thymineless induc-tion in Escherichia coli K12(X). Biochim. Biophys. Acta 61:775-790.
-17. Lanka, E., and H. Schuster.1970.Replication of bacteri-ophages inEscherichia colimutantsthermosensitive in DNA synthesis. Mol. Gen. Genet. 106:274-285. 18. Lark, K. G.1972.Genetic controloverthe initiation of the
synthesis of the shortdeoxynucleotide chainsinE. coli. Nature N. Biol.240:237-240.
19. Mikolajczyk, M., and H. Schuster. 1971. The fate of
newly made DNA in Escherichia colimutants ther-mosensitive in DNA synthesis. Mol. Gen. Genet. 110:272-288.
20. Monk, M., and J. Gross.1971.InductionofprophageAin
amutantofE.coli K12 defective in initiation of DNA replication at high temperature. Mol. Gen. Genet. 110:299-306.
21. Noack, D., and S. Klaus. 1972. Inactivation kinetics of lambdaphage repressors ina mutant of E. coli
tem-perature sensitive in DNA replication. Mol. Gen. Genet.115:216-224.
22. Sugino, Y., A. Sorai, T. Kogoma, Y. Hirota, and R. Okazaki. 1966. Inhibition of DNAsynthesis and induc-tion of Lambda phage in temperature-sensitive
mu-tantsofEscherichiacoli.Jap. J. Genet. 41:247-249.
23. Wechsler, J. A., and J. D. Gross. 1971. Escherichia coli
mutants temperature-sensitive for DNA synthesis. Mol.Gen. Genet. 113:273-284.
24. Wechsler, J. A., V. Niisslein, B. Otto, A. Klein, F. Bonhoeffer, R. Herrmann, L.Gloger, and H. Schaller. 1973. Isolationandcharacterization of thermosensitive Escherichia colimutantsdefective in deoxyribonucleic acidreplication. J. Bacteriol. 113:1381-1388.
25. Worcel, A. 1970. Induction ofchromosome re-initiations inathermosensitive DNAmutantof Escherichia coli. J. Mol. Biol. 52:371-386.
26. Yoshikawa, H., and M. Haas. 1968.On the regulation of the initiation of DNA replication in bacteria. Cold Spring Harbor Symp. Quant. Biol. 33:843-855. 27. Young, E. T., and R. L. Sinsheimer. 1967. Vegetative
bacteriophage A DNA.I. Infectivity ina spheroplast assay.J. Mol. Biol.30:147-164.