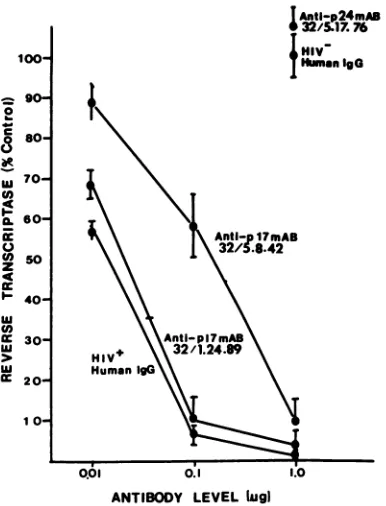
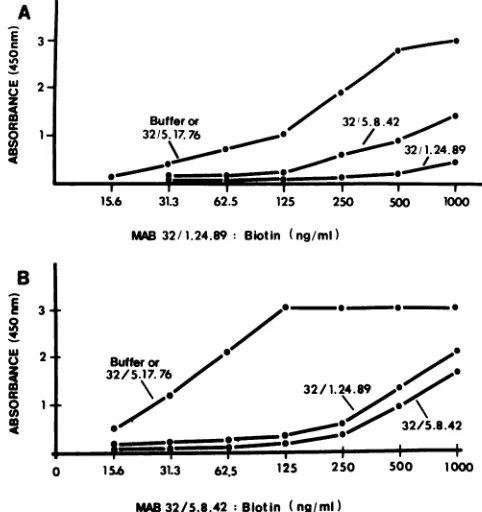
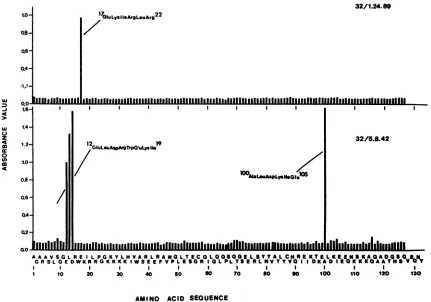
Human immunodeficiency virus type 1-neutralizing monoclonal antibodies which react with p17 core protein: characterization and epitope mapping.
Full text
Figure

Related documents
It is contended that, a general-purpose method of formulating and solving dynamic programming problems will help practically oriented students to learn, adopt and apply the technique
The five scales are the ones that Karavas-Doukas (1996) had used. Negatively-worded items have been recoded so that 6 is the positive end of the scale. The highest mean is for
The clinical study on Uragan Vatham was carried out at the post graduate Noi Naadal Department of Government Siddha Medical College Hospital, Palayamkottai. Case Selection
Numerous studies of human immunodeficiency virus type 1 (HIV-1)-specific cytotoxic T lymphocytes (CTL) have examined their ability to recognize B-cell lines expressing recombinant
At 127 weeks p.i., FIV-specific pCTL activity was detected with lymphocytes isolated from the lymph nodes of both FIV-infected cats.. In cat F1, this lymph node pCTL activity was
For most of the glycoproteins expressed so far in insect cells, the oligosaccharide side chains have been reported to differ markedly from their vertebrate counterparts in that
At various times following HSV-1 corneal infection, the eyes of RB6-8C5- treated and control SCID mice were collected, and the infec- tious virus titer in each eye was
Plasmids pSlX (expressing L, M, and S HBsAg) and pS2X (expressing M and S HBsAg) can help to package the HDV genome (Fig. 2A, lanes 2 and 3) as efficiently as the plasmid containing
