Vol. 46, No. 1 JOURNALOFVIROLOGY,Apr. 1983,p.177-186
0022-538X/83/040177-10$02.00/0
CopyrightC1983, American SocietyforMicrobiology
Structural and Antigenic Analysis of the Nucleic Acid-Binding
Proteins of Bovine and
Feline Leukemia Viruses
M.ALICEMORGAN, TERRY D. COPELAND,ANDS.OROSZLAN*
Biological Carcinogenesis Program, National Cancer Institute-Frederick CancerResearchFacility,
Frederick,
Maryland21701Received 9 September 1982/Accepted6January 1983
The nucleicacid-binding proteins of bovine leukemia virus (BLV) and feline leukemia virus (FeLV) were isolated in ahigh state of purity with chloroform-methanol extractionfollowed by reversed-phase liquid chromatography. Selective solubilization and purity of BLV p12 and FeLV plOwas confirmed by sodium dodecylsulfate-polyacrylamide gel electrophoresis. The compositions and molec-ular weightsweredetermined by amino acid analysis. An abundance of lysine and arginine residues along with their size identifies both BLV p12 and FeLV plOas small basic proteins similar to well-defined type C viral nucleoproteins. NH2-terminal degradation by the semiautomated Edman method provided the se-quence of the first 40 amino acids for both proteins. The putative nucleic acid binding site found in severaltypeC viralnucleoproteinswascontained within this
sequence, withthe most homology centered aroundan eight-amino acid region involving sevenidentical residues andonesubstitution. Antiseraweredeveloped in rabbits, and specificity and titers were determined by electroblotting and immunoautoradiography. By this technique,animmunological cross-reactionwas foundbetween BLV p12 and FeLVplO. The sharedantigenic determinantmost
likely exists in the highly conserved eight-amino acid region. Although this sequenceis alsohighly conserved in the nucleic acid-binding proteins of murine leukemiaviruses, the shared antigenic determinant isnotfound in these orany other type C viruses tested. It is suggested that substitution of arginine (BLV pl2/FeLV p10)tolysine (murine leukemia virus p10) is sufficienttoelicitachange inantibody specificity.
Bovine leukemia virus (BLV) and feline
leu-kemia virus (FeLV) are non-genetically
trans-mittedexogenousretroviruses showntobe
asso-ciated withdiseasein theirrespective hosts (4,
5).
BLV is regarded as the causative agent of
enzootic bovine leukosis, a lymphosarcoma,
found in domesticated cattlewidespreadin both
North America and Europe.Itinfects20%of all
dairy cattle and isfound in 60%o of all herds in
theUnited States.It canbetransmitted
horizon-tally through cell-to-cell contact or insect
vec-tors and vertically by congenital means (6).
Recently infectious BLV was demonstrated in
themilkofdairycows(7).Itisalso infectiousin
otherspecies includingsheep, goat,pig (18), and
chimpanzee(32).The gag geneproductsarethe
internal structuralproteins;p24,themajor
struc-tural protein (8); p15, probably a major
phos-phorylatedcomponent(31);p12,thenucleic
acid-binding protein (NBP) (17); and p1O, which is
less well characterized. The precursor to gag
geneproductsisa-65,000-daltonprotein(Pr65)
whichwasshownto containp24, p15, p12,and
plOby trypticpeptide mapping (9).BLV has not
been readily classified into one of the
well-defined groups of retroviruses. For example,
antiserum to BLV p24 failed to detect
cross-reactivity with the internal protein of FeLV,
Rauscher murine leukemia virus (R-MuLV),
foamy-likebovine syncytiavirus, Mason-Pfizer
monkey virus (8, 20), oravian oncornaviruses
(19).
FeLV is alsoanaturally occurring infectious
retrovirus which causes leukemia and
lympho-sarcomaincats and isresponsible for one-third ofallcancerdeaths. Theprimarymodeof virus
transmission is horizontalthrough saliva,
infect-ing most free-roaming pet cats. Pathogenic
forms of leukemiacanbeinducedby inoculation
ofcats with FeLV (5). FeLV canalso grow in human embryonic lungcells, dog kidney cells,
and pig embryo cells (15) and can produce
lymphosarcoma indogs (28). FeLV strainscan
bedivided intosubgroups A, B,andCbased on
theirpatterns of attachment toreceptorsatthe
hostcell membrane. The gaggene-encoded
in-ternal proteins are p15, p12, p27, andplO(15),
177
on November 10, 2019 by guest
http://jvi.asm.org/
178 MORGAN, COPELAND, AND OROSZLAN
whicharesimilarin propertiestotheproteins of
MuLV. Based on protein sequence homology
and immunological relatedness with MuLV,
FeLVis classifiedas typeC, subgroup I (23).
The only BLV protein partially sequenced is
p24,which is foundtocontainsequence
homolo-gy with type C virus p30 structural proteins,
especially with FeLVp27, suggesting thatBLV
p24 and FeLV p27 are evolutionarily related
proteins (22). BLV p24 has recently been found
(25)tohavestructural homology with the
analo-gous24,000-molecular-weight proteinfroma
vi-rus associated with human adult T-cell
leuke-mia-lymphoma and designated HTLV (26).
Several linesof evidence indicate that the NBPs
of the virionmaybeamongthemostconserved
gaggeneproducts ofretroviruses. Asthe
RNA-associated internal core proteins of the virus,
theymayhaveanimportant functionin
replica-tion and assembly and theymaybeunder high
evolutionary constraint. As a continuation of
ourstudies onthe BLV-FeLVrelationship, we
sequenced and immunologically characterized
the NBPs of these viruses. Theprimary
struc-ture data of BLV p12 and FeLVplO show the
presence ofahighly conserved segment in the
putative nucleic acid binding site region. This
andthe results of immunologicalanalysiswhich
indicate cross-reaction permit the delineation of
the shared antigenic determinant.
MATERIALS AND METHODS
Virus. BLVwas grownin fetal lamb kidney cells (33),and theRickard strain(A, B)of FeLVwasgrown in feline lymphoblasts (27). Viruses were purified by sucrose density gradient centrifugation according to standardproceduresandobtainedfrom theViral Re-sources Laboratoryof theNational Cancer Institute-Frederick Cancer ResearchFacility, Frederick,Md.
Chloroform-methanol extraction. NBPs of BLV and FeLVwerepartiallypurified bychloroform-methanol
extraction (21). Chloroform-methanol (2:1, vol/vol) wasaddedtothevirussuspensioninthepresenceof low-ionic-strength buffer, 0.01 MTris-hydrochloride, pH 7.4)-0.001 MEDTA-0.05 M NaCl. Aftervigorous agitation for 2 min, the phases were separated by
centrifugation at 2,000 rpm for 20 min at 4°C. The phosphoproteinand RNApartitionedtotheaqueous
phase,thevirallipidspartitionedtotheorganic phase, and theremainingviralproteinswerecontained in the interphase.To solubilize BLVp12and FeLVplO,the interphase was resuspended in high-ionic-strength buffer, 0.01 M Tris-hydrochloride (pH 7.4)-0.001 M EDTA-1.0 MNaCl,and theextractionwasrepeated
with the NBP contained in the high-ionic-strength
aqueousphase.
RPLC. Further purification was performed by re-versed-phase liquid chromatography(RPLC)on a Wa-ters> BondapakC18 column(13).The NBPcontained in theaqueousphasefromhigh-ionic-strength chloro-form-methanol extraction was lyophilized and then resuspended in 6 Mguanidine-HClandadjustedtoa pH of2.0withl1otrifluoroacetic acid. A 0to 30%o
gradientof acetonitrile in 0.05% trifluoroacetic acidat pH 2.0 wasusedtoelute the NBP from the columnby reduction of the hydrophobic interactions. Proteins wererecovered bylyophilization.
Reductionandcarboxanidomethylation. Proteinwas reduced in the presence of6Mguanidine-HCI-0.1 M NaHCO3 (pH 8.5) buffer with 0.17 M dithiothreitol.
Sampleswereflushed with N2 and incubatedatroom temperature for 3 h. Reducedproteinswere
carbox-amidomethylated with 0.4 M iodoacetamide (room temperature;13to14h).
SDS-PAGE.Sodiumdodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE), according to the formulation of Laemmli(17), wasusedtodetermine
purity of the NBPs and approximate molecular weights.
Amino acid analysis.Thecompositionand molecular weightof the NBPs weredetermined by amino acid
analysis on a DurrumD500 amino acid analyzeras
previouslydescribed (11). Therawdatawereanalyzed byaprogramrunontheNational Institutes of Health DEC 10 computer which selectsamolecularweightfor the protein based in suchaway as to minimize the departure fromintegralvaluesforallresidues (3).
NH2-terminal sequence analysis. NH2-terminal se-quence analysis by semiautomated Edman degrada-tionwasperformedwith thespinning-cupliquid phase system (24) on a Beckman 890C sequencer in the presenceof Polybrene (30). RPLC on a Waters
phenyl-alkylcolumnwas usedtoidentify andquantitate the PTH derivativesofaminoacids(10).
Antisera.AntiseratothepurifiedNBPs were devel-oped in rabbits. The initialinjectionwasmade
intra-dermallyin thebackfootpadsand subcutaneously in four sitesonthehindof the rabbit with 150 ,ug of NBP in phosphate-buffered saline mixed with an equal volume of Freund complete adjuvant. The rabbits wereboostedsubcutaneouslyapproximatelyevery10
days with 50 Fg of purified protein each time in
phosphate-bufferedsalinemixed withanequalvolume
of Freund incomplete adjuvant. Ten days after the secondboost,thefirsttestbleed of 10 ml from theear veinwasobtained. The rabbitswererepeatedly boost-ed and then blboost-edaccordingtothis schedule. Animals immunized with BLVp12andFeLV plOwerebledout
byheart punctureafter 130 and 182days, respectively.
Anti-BLV p24guinea pig serum andanti-FeLV p27 rabbitserum wereavailable frompreviousstudies(8). Electroblotting and lmmunoautoradlography. The
specificities of the antiseraweredeterminedby elec-troblotting and immunoautoradiography (29). BLV and FeLV wereelectrophoresed on7.5to20%o
SDS-polyacrylamideslabgels.Afterelectrophoresisthegel
was equilibrated in transfer buffer (0.04 M sodium phosphate, pH 6.5). Aminobenzyloxymethyl paper was converted to the active diazobenzyloxymethyl form(DBM paper)bytreatmentwithfreshly prepared
4mMNaNO2 in 1.17MHCI for 30 minoniceandthen washed with water and equilibrated with transfer buffer. Virus was transferred from the gel to DBM paperelectrophoreticallyfor 2h at25 V.Remaining active groupsonthepaperwereblockedafter incuba-tionat roomtemperature for2h with1% bovine serum albumin in transferbuffer. The paper was rinsed in waterandcutvertically into 0.5-cm-wide strips (repre-sentingalane ofseparated viral proteins from SDS-PAGE) whichwererotatedat37°C in rabbit antiserum J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
ANALYSIS OF NBPs OF BLV AND FeLV 179
2.0
A B
1.0!-C
A30
-0 10 20 30 40 0 10 20 30 40 0 40 50 60
[image:3.490.46.447.62.377.2]Time (Min)
FIG. 1. RPLC of(A)BLVp12, (B) FeLVplO, and (C)reduced andcarboxamidomethylatedBLVp12.
diluted in 0.05 MTris-hydrochloride(pH7.4)-0.005M EDTA-0.15 M NaCl-0.25%gelatin-0.05% Nonidet P-40(TENG-N) for15 to16 h. Thestripswererinsed in water and incubated first at 37°C for 2 h in four changes ofTENG-Non ashakingwaterbathand then for 2h in125I-labeledproteinAat40,000to50,000cpm perml of TENG-N.Labelwasremovedandthestrips
werewashed with four changes of 0.05 M Tris-hydro-chloride(pH7.4)-0.005M EDTA-1.0 M NaCl-0.25% gelatin-0.4% Sarkosyl (TENG-S)for 2 hat37C. The stripswererinsed in water, airdried,andexposedto flashedX-rayfilm withanintensifyingscreenfor 15to 16h.
DOTtests.Antibody titersweredeterminedby direct-ly spotting DBM paper strips with 1 Fg ofpurified
NBP in0.04M sodiumphosphate buffer, pH6.5. With this methodanexact amountofproteinwasboundto DBMpaper.Reaction with serial dilutions ofantisera,
2'"I-labeledprotein A, and autoradiographywere
car-riedoutasdescribedabove.
RESULTS
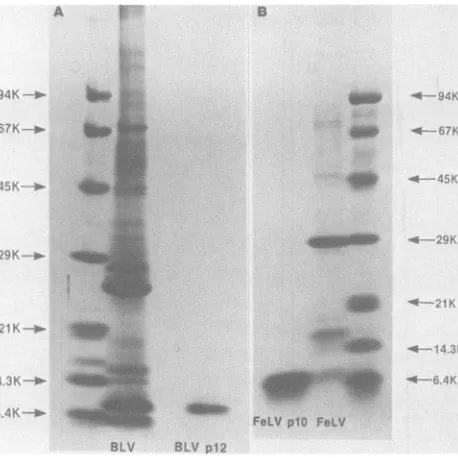
Purification of BLV and FeLV NBPs. The
NBPs of BLV and FeLV were purified by chloroform-methanolextraction andRPLC. The proteins partitioned to the high-ionic-strength aqueous phase during extraction with chloro-form-methanol.Anapproximate purity of >95%
afterchloroform-methanolextractionwas deter-mined by SDS-PAGE. RPLC was used to re-move possible contaminating protein
compo-nentsand RNA andallow for desalting and rapid concentration ofprotein bylyophilizationfroma volatile solvent. RPLC profiles of the extracted proteins from BLV, FeLV, and reduced and carboxamidomethylated protein from BLV are shown inFig. 1A, B, and C, respectively. Both nucleoproteins elutedat25%acetonitrileasthe majorcomponentof theextraction. The elution
patterns of BLV and FeLV NBPs indicated multiple protein peaks (three) eluting within a narrow range of acetonitrile gradient. When BLV p12 was reduced and carboxamidometh-ylated the elutionpatternshowedasingle major
peak, suggesting that the apparent chromato-graphic heterogeneitymayhavebeen duetothe formationof inter-orintramolecular S-S bonds.
The purity of the proteins and approximate molecular weights were determined by SDS-PAGE.Purified NBP fromBLVis demonstrated inFig. 2A. InoursystemithasanMrof-8,000 comparedwith standard proteins. Previous no-menclature has identified this protein as BLV p12 (4). We will use the same designation. Figure 2B confirms the purity of FeLVp1O, the U
e
0
a A0 4c
20 ' c 00
00
l0
VOL.46,1983
/
I
on November 10, 2019 by guest
http://jvi.asm.org/
180 MORGAN, COPELAND, AND OROSZLAN
-4 ..-
-.; ,_, * t W .S,,.1
...r b _Lb b * E
* E
_ r
*- Fo *9 E [ 's... .FR
.. s. ::.
*'- 1!o _
7 _L
*
* s
[image:4.490.137.366.77.306.2]-.o. _
... ...
...,,X,4.
_
< . ... i:
.L
--@- _
FeLVp10 FeLV
FIG. 2. SDS-PAGE of BLV virus and purified BLV p12 (A) and of FeLV virus and purified FeLV plO (B).
Standard proteins (molecular weights in parentheses) are phosphorylase b (94,000), bovine serum albumin (67,000), ovalbumin(45,000),carbonicanhydrase(29,000), soybean trypsininhibitor(21,000),lysozyme (14,300), andaprotinin (6,400).
NBPfrom FeLV, which hasamolecularweight
of-6,000.
Amino acid composition. The aminoacid
com-positional data of BLV p12 and FeLV p1O,
based on 24-,48-, and 72-hhydrolysis,aregiven
in Table 1. The total number of amino acids
TABLE 1. Aminoacidcomposition ofBLV
p12
and FeLV plOBLVp12 FeLVplO
Aminoacid Residues Nearest
ReiusNearest
per
ineger
per integer protein tgr protein inerAsp 3.72 4 6.09 6
Thr 2.90 3 1.81 2
Ser 2.33 2 1.93 2
Glu (2.1) 2 5.29 5
Pro 14.83 15 3.70 4
Gly 5.82 6 3.23 3
Ala 3.08 3 3.25 3
Val 1.26 1 1.99 2
Met 0.71 1 0.00 0
Ile 1.03 1 0.94 1
Leu 2.04 2 3.59 4
Tyr 0.82 1 1.02 1
Phe 0.00 0 0.00 0
His 2.18 2 0.99 1
Lys 7.36 7 8.78 9
Arg 4.82 5 5.76 6
Cys (not determined) Trp (notdetermined)
Molwt 6,034 5,652
minus cysteine and tryptophan is 55 for BLV
p12 compared with 49 forFeLV p1O. Note the
abundance of basic amino acids, lysine and
arginine, in bothproteins. The numberof
resi-dues ofserine,alanine, isoleucine, andtyrosine
areidentical forbothproteinsas istheabsence
ofphenylalanine. BLV p12 has a high proline
content notsharedbyFeLVp1O.The
computer-assisted molecularweightsfrom thecomposition
(withoutcysteineand tryptophan) are 6,034for
BLVp12 and5,652for FeLVp1O.
NH2-terminal sequence. TheNH2-terminal
se-quenceasdeterminedbysemiautomated Edman
degradation of BLV p12 is shown in Fig. 3A.
Assignmentswerebased on theoccurrenceof a
newlyarising peakateachcycle compared with
standardPTH amino acids. In asingle
microse-quenceanalysis of4nmolofBLVp12,
unambig-uousassignmentsweremadefor residues1to 40
except cysteines in positions 24, 27, and 37,
leucineresidue 28(which was lost), and
threo-nine 39. Cysteine residues cannot be identified
after Edmandegradationoftheunmodified
pro-tein duetotheirinstability. Thecysteines were
determined by degrading purified peptides
de-rived by endoproteinase
Lys-C
cleavage ofre-duced and carboxamidomethylated BLV p12.
These two peptides, one representing the
frag-ment of residues 18to 29 and theother
repre-sentingthatofresidues 30 to40,wereavailable
from other studies. Sequence analysis of the
peptides confirmed the initialresults of Edman
J. VIROL.
., :.-..>.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 46, 1983
BLV p12
Val-His-Thr-Pro-Gly-Pro-Lys-Met-Pro-Gly-Pro-Arg-Gl n-Pro-Al a-Pro-Lys-Arg-Pro-Pro a 2.9 1.5 3.6 3.2 1.9 2.0 3.1 1.4 2.7 1.8 1.3 3.0 1.6 1.7 1.5 1.3 2.1 2.1 0.9 1.9
b 4. 3 4. 7 5.3
ANALYSIS OF NBPs OF BLV AND FeLV 181
10 15 20
21 25 30 35 40
Pro-Gly-Pro-Cys-Tyr-Arg-Cys-Leu-Lys-Gl u-Gly-Hi s-Trp-Al a-Arg-As p-Cys-Pro-Thr-Lys
a 0.2 0.3 0.4 0.2 0.2 0.1 0.2 0.1
b 6.8 6.2 4.5 2.9 7.5 6.2 2.3 4.1 1.1 17 14 17 8.9 19 17 16 6.2 3.1 2.9 2.5
FeLV plO
1 5 10 15 20
Ala-Thr-Val-Val-Al a-Gln-Asn-Arg-Asp-Lys-Asp-Arg-Gl u-Gl u-Lys-Lys-Leu-Gly-Asp-Gln a 6.8 4.7 9.0 5.6 5.7 3.4 3.0 3.4 3.3 4.8 2.9 2.4 3.7 3.3 3.2 5.1 2.1 1.3 2.1 2.0
21 25 30 35 40
Arg-Lys-Ile-Pro-Leu-Gly-Lys-Asp-Gln-Cys-Ala-Tyr-Cys-Lys-Glu-Lys-Gly-His-Trp-Val a 2.2 4.1 1.6 1.9 1.0 1.3 1.1 1.1 1.0 0.7 1.5 0.7 1.3 0.5 0.2 0.2 0.5
[image:5.490.56.427.53.398.2]b 25 10 16 9.8 8.9 8.6 11 15 4.2 4.4
FIG. 3. NH2-terminalamino acid sequence of BLVpl2and FeLVplO. Numbers belowresiduesareyieldsin nanomoles ofPIHamino acid derivatives as obtained by degrading the unmodified proteins (rows a) and
fragments (rows b). Inthe initialanalysis ofBLV p12the assignmentsfor residues 21 to 30werebased on qualitative data due to instrument malfunction which didnotallow accurate quantitation. Inputs for Edman
degradationwere asfollows: BLVp12-intactprotein,4nmol;fragment18to29,21nmol; fragment30to40,24 nmol; FeLVp10-intact protein,12nmol;fragment23to27,33nmol;fragnent28to34, 18nmol.
degradationandalso allowedassignments to be
made atpositions 28 and 39 asshown.
Figure3B shows thesequencefor FeLV p1O.
The first40 amino acidresidues, except cysteine
residues 30 and 33 and lysine 27, were
deter-mined by a single microsequencing analysis of
12nmol ofprotein. Assignmentsof the cysteines
and lysine residue 27 were made as previously
discussed for BLV p12, using Lys-C cleavage
fragments
(T. D.Copeland, M.A. Morgan, andS. Oroszlan, manuscript in preparation). BLV
p12and FeLVplONH2-terminal sequences are
comparedforhomologywith each other and the
aminoacid sequenceofthe NBPfromR-MuLV
(12) in Fig. 4.
Immunological
characterization of BLV p12and FeLV p1O. Antisera specific for the NBPs
were developed in rabbits. The immunization
5 10 15 20 25 30 35 40
FeLV plO A T V V A Q N R D K D R E E K K L G D Q R K I P L G K )Q C A Y C K E K G H W V R D C P K BLV p12 V H T P G P K M P G P R Q P A P * * * K R P P P G P * * * C Y R C L K E G H W A R D C P T R-MuLV plO A T V V S G Q R Q * D
R.Q
G G E * * * R R R P Q L D R D Q C A Y C K E K G H W A K D C P KFIG. 4. NH2-terminal sequences of BLVp12, FeLVp1O, and R-MuLVplOarecompared insingle-letter aminoacid code:A,alanine;T,threonine;V, valine; Q, glutamine;N,asparagine; R,arginine; D,aspartic acid; K,lysine; E, glutamic acid; L, leucine; G, glycine, I, isoleucine; P, proline;Y, tyrosine; H, histidine; W,
tryptophan; M, methionine; S, serine; C, cysteine. Any residue identical witharesidue fromeither of the other
proteinsis underlined. Gaps are indicated by an asterisk. The sequenceof FeLV plObeyondresidue 40 istaken fromotherstudies(Copelandetal.,unpublisheddata). The sequence of R-MuLV plO is takenfrom Hendersonet al.(12).
1 5
on November 10, 2019 by guest
http://jvi.asm.org/
182 MORGAN, COPELAND, AND OROSZLAN
16
12
0
x
4
6
X, 4
0
x
-F
2
BLV p12 Immunization
20 60 100
Time (days)
140 180
FeLV plO Immunization
20 60 100 140 180
Time (days)
FIG. 5. Immunization schedules and titers of anti-BLVp12rabbit serum andanti-FeLV plO rabbit serum.
Symbols:0,testbleed; l , injection.
schedules with the resulting antibody titers for
BLV p12 and FeLV plO are shown in Fig. 5. Arrowsindicatedayswheninjections(a total of
nine for each rabbit) were made, and circles
indicate the titers oftest bleed and bleed-out
sera.Eachserum wastitratedby
immunoautora-diography, usingtheDOTtest. Thehighesttiters
obtainedfor anti-BLVp12serum, and FeLVplO
antiserumwere16,200and5,800. Atotalof550
jig
of each purified protein was used for eachrabbit. Immunization with BLV p12 was
com-pletedin 130daysand thatwithFeLVplOwas
completedin 182days. Inspiteoftherelatively
large doses ofantigensandlong periodof
immu-nization. theresultingantiserawerehighly
spe-cific. This reflects onthehigh stateofantigenic
purity oftheRPLC-purifiedproteins.
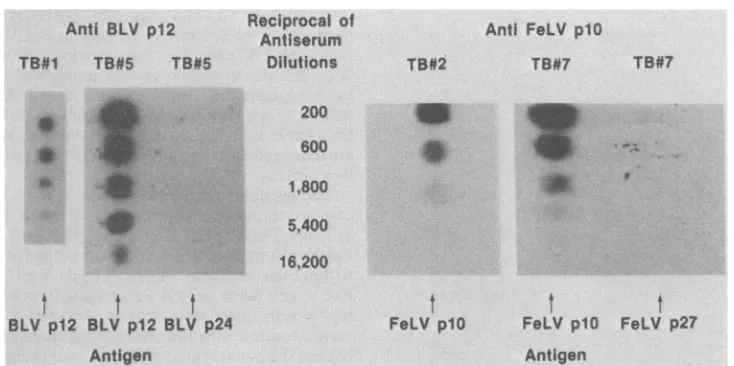
Figure 6 shows an actual titrationpattern of
anti-BLVp12 and anti-FeLV plO rabbit seraas
obtained by the DOT test. All test bleeds and
bleed-out serawere titrated by this procedure,
butonlyresultsoftwobleeds(oneearlyand one
later) from each rabbit immunized with the
NBPs areshown. Test bleed1of anti-BLVp12
rabbitserumtitratedat1,800 withanincreaseto
16,200at testbleed5, which didnotreactwhen
tested against BLV p24. Test bleed 2 of
anti-FeLVplO rabbitserumtitratedat1,800 withan
increase to 5,400 in test bleed 7, which was
negative when spotting FeLV p27. Prebleeds
J.VIROL.
i i i i i i i
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.490.99.398.67.482.2]ANALYSIS OF NBPs OF BLV AND FeLV 183
Anti BLV p12
TB#1 TB#5 TB#5
Reciprocal of
Antiserum Dilutions
200
600
1,800
4:
0:
.0
A*
.w
TB#2
Anti FeLV p10
TB#7 TB#7
I
*_
5,400
16,200
t
t
1
BLV p12 BLV p12 BLV p24
Antigen
t
t
t
FeLV p10 FeLV p10 FeLV p27
Antigen
FIG. 6. Titration of anti-BLVp12rabbitserum and anti-FeLV plOrabbit serum by NBPspottingof DBM paper. TB, Test bleed.
taken before immunization of eitherrabbitwere
negative.
Specificity of antiseratoBLVp12 and FeLV
plO wasfurther studied by
immunoautoradiog-raphy involving the electrotransfer of
SDS-PAGE-separated viral proteins to DBM paper
(Fig. 7). In lanes 1 to 3 FeLV proteins were
transferred and stained with appropriate
anti-sera.Lane1shows thereaction with anti-FeLV
p27 rabbit serum. Lane 2 wasstained with
anti-FeLVplOrabbitserumwhich reacted with plO
only. Thesameprotein wasalsorecognized by
antiserum toBLVp12asshown in lane 3. BLV
proteins were transferred in lanes 4 to 6 and
stained withappropriate antisera. Lane 4shows
the reaction of BLV with anti-BLV p24guinea
pig serum. Lane 5 was stained with anti-BLV
p12rabbitserum.Rabbit antiserumtoFeLVplO
also cross-reacted withBLVp12. Both antisera
to BLV and FeLV NBPs reacted specifically
withBLV, recognizingp12only. Various other
retrovirusesweretestedwith anti-BLVp12and
anti-FeLV plO rabbit sera after virus proteins
weretransferredto DBM paper. Table 2
summa-rizes these results.Asexpected, anti-FeLVplO
serum reacted in an interspecies fashion with
other mammalian type C virus NPBs but not
with Rous avian sarcoma virus p12. The anti-BLVp12serum,however, didnotrecognizeany
type C virus testedexcept FeLV. Homologous
andheterologous titers (numbers in parentheses)
for both seraarealsogivenin Table 2.
DISCUSSION
Chloroform-methanol extraction and RPLC in
combination are effective methods in which to
purifyto ahighstateof puritythe NBPsofBLV
and FeLV. The purity of BLV p12 and FeLV
plO was confirmed by SDS-PAGE. In spite of theapparenthomogeneity in thissystem,RPLC
patterns indicatedmultipleprotein peakseluting
withina narrow rangeofanacetonitrilegradient.
This heterogeneity of
chloroform-methanol-ex-tracted BLV p12was showntobe dueto
disul-fide cross-links between cysteine residues. The
reduced and carboxamidomethylated protein
elutedas asingle peak.Itis assumed that similar
intra- or intermolecular cysteine interactions
may be responsible for the chromatographic
heterogeneity of FeLVp1O. Itis clear from the
partial sequence data that both proteins have
morethanonecysteine.
Theaminoacidcompositiondataindicate that
both NBPs containanabundanceofbasic amino
acids. Noaminoacidappeared withhigher
resi-due number than lysine plus arginine in either
proteinexceptthe 15residues ofprolinefound in
BLVp12. This unusually highfigureforproline
in BLVp12 isnotshared in thecompositionsof
FeLVplOandof other known NBPsoftypeC
viruses. Recently, the primary structure of the
NBP (p1O) of R-MuLV was reported (12).
R-MuLV plOisabasicproteincontaining5lysine,
9arginine, and 2 histidine residues,atotalof 16
basic amino acid residues of 56 in thecomplete
protein. Gel filtration andelectrophoretic
analy-sis hadoriginally estimatedthemolecularweight
to be between 7,000 and 10,000. The exact
molecularweight is reportedas6,347. The
mo-lecular weights calculated from composition
(without cysteineand tryptophan) forBLV p12
(6,034) andforFeLVplO(5,652)arealso smaller
thanmolecularweights determinedby gel
elec-trophoresis. Results indicatethat BLVp12and
FeLV plO are similar in size and composition, VOL.46,1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.490.63.430.73.256.2]184 MORGAN, COPELAND, AND OROSZLAN
FeLV LV
po27.
;F
F
FIG. 7. Specificity of anti-BLV p12and anti plO rabbit sera by immunoautoradiography. was transferred to DBM paper (lanes 1 to
stained with anti-FeLV p27 rabbitserumataI
dilution (lane 1), anti-FeLV plO rabbit serur
1:6,000 dilution (lane 2), and anti-BLV p12
serumata1:100 dilution(lane 3).BLVwastran! toDBMpaper (lanes4to6)and stained withant p24 guinea pigserumata1:2,000dilution(lane4 BLVp12 rabbitserumata1:10,000dilution(I
and anti-FeLV plO rabbit serum ata 1:100 d
(lane6).Arrowsindicate positionofproteinafte PAGE.
exceptfor proline in BLV p12 (see above), structurally defined MuLV viral NBP.
The alignments of FeLV p1O, BLV pl
R-MuLVplOareshowninFig.4.BLVpl
FeLVplONH2-terminalsequences dono
tain much homology in the first 40 residi
the proteins. The most homology betwee
proteins centers around a common regioi
contained in R-MuLV p1O. This conserv
gion has been expressed as aset of thre
residuesspacedatN, N + 3,andN+ 13
Gly-HissequenceatN+ 7andN+8, wh
isthepositionin theamino acidsequence
firstCys residue ofthe set(cysteineinpc 30 in the alignment). Involvement of ty:
andlysine residues in nucleic acidbinding;
tyofproteins has been indicated with stud
the chemical modification of R-MuL%
(L. E. Henderson, C. W. Long, and S. (
lan, Fed. Proc. 39:1606, 1980) and the
binding protein coded for by gene 5 of
riophage fd(2).Tyrosinehas also beenimj
ed intheability ofgene32protein from bac
phage T4 to bind to single-stranded DN
The conserved region of FeLV plO co
bothtyrosineandlysineresidues andaG
sequenceattheappropriate positions. Th
teines and theGly-His sequenceof BLVp12 in
positions 31 and 32 align with that in FeLVplO
and R-MuLV p1O. The homologous region of
BLV p12 alsocontains tyrosine and lysine. Due
to their sequence homology with a
well-identi-fied NBP inaregion consideredtobeimportant
fornucleic acid binding, BLVp12 and FeLVplO
aredeterminedtobe the NBPs from the
respec-tive viruses.
The results from immunological studies
indi-cate an authentic cross-reaction between BLV
p2;4
p12
and FeLVp1O.
Therefore,
BLVp12
andFeLV plO mustshare acommonantigenic site
within theirstructure. AntiseratoBLV p12 and
FeLV plO were tested in
immunoautoradiog-_p% raphy with other retroviruses. Anti-FeLV plO
serumreactedwith the NBPs ofall mammalian
butnottheaviantypeC viruses tested (Table 2).
Anti-BLV p12 serum does not react and thus
doesnotshare antigenic determinantswith any
v-FeLV of the other viruses testedexceptFeLV. These
FeLV results also show that antigenic determinants
3) and shared between feline plO andmurine plOare
1:6,000 not present in BLV p12. The comparison of
[image:8.490.51.243.63.272.2]m at a NH2-terminal sequences (first 40 residues) of
febrred
theseproteins
(see
Fig.
4)
doesnotdemonstratesi-BLV
ahomologous
region
between FeLVplO
and1), anti- BLV p12 not shared by R-MuLV
p1O,
whichane5), couldreadilyaccountfor the
FeLV/BLV-specif-lilution ic antigenic cross-reaction(seeabove).
rSDS- The amino acid sequence of BLV p12 and
FeLV plO shows a highly homologous region
beginning with Gly-His, positions 37 and 38 in
tothe the alignment ofFig. 6. Within this
eight-resi-due-long homologous region there is only a
2,and
12and
t con- TABLE 2. Reaction of anti-FeLVplOand anti-BLV
ues of p12rabbitserawith the NBPs of various retroviruses
,n the aftertransferof separated viral proteins from
SDS-n also PAGEtoDBMpaper
ed
re-e Cys
anda
ereN of the
sition rosine
activi-lieson
plO
)rosz- DNA-
bacte- plicat-
cterio-A (1). ntains ly-His
e
cys-Reaction with antiserumto:
Virus' FeLV
plOb BLVpl2C
BLV +(400)d + (16,200)
FeLV +(5,400) + (1,600)
M-MuLV +(1,600)
-RD-114 +
R-ASV
-a Besides BLV and FeLV, the following viruses weretested: Moloney-MuLV, catendogenous type C virus (RD-114), and Rous avian sarcoma virus (R-ASV).
bAntiserum was used at 1:6,000 dilution in the homologous system and 1:100 in the heterologous systems.
c Antiserum dilutionswere1:10,000in the homolo-gous systemand1:100in theheterologous systems.
d The numbers in parentheses represent the titers of bleed-outseraasdeterminedbytheDOTtest.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 46,1983
single amino acid difference between FeLV and
BLV. Alanine of BLV p12 is substituted for
valine in FeLV p10. In this position R-MuLV plO has alanine, like BLV p12. C-terminal to this
residue, both bovine p12 and feline plO have
arginine whereas the murine plO has lysine.
Since the complete amino acid
sequence
of BLVp12 (Copeland et al., manuscript inpreparation)
and the nearly complete sequence of FeLV plO
(T.
D.Copeland
etal., unpublished data)
donotshow ahigher degree of homology in the other
parts of the molecules, the above eight-residue
segment is the mostlikely candidate to represent
the shared antigenic site responsible for BLV
pl2/FeLVplO cross-reaction readily detectable
on denatured proteins. The Ala-Val exchange
may not be considered significant enough to
altertheimmunogenic response and subsequent
antigen-antibody complexing. Although arginine
andlysine are both basic amino acids, theyare
stericallydifferent. This difference could explain
why anti-BLV p12 serum reacts with FeLVplO
but not with the murinep10.Chemical
modifica-tion of arginine andlysine residues and the use
ofsyntheticpeptideswillaidthe accurate
defini-tion of the immunogenic determinant shared by
BLV andFeLV.
ACKNOWLEDGMENTS
Wegratefullyacknowledge theexcellent technical assist-anceofCatherine V.HixsonandYoung Kim. We also thank Donald C. Fish andcolleaguesforassistance with immuniza-tions.
This research was sponsored by thePublicHealthService National CancerInstitute under contractN01-CO-75380with LittonBionetics.
LITERATURECITED
1. Anderson, R. A., andJ.E.Coleman.1975. Physiochemi-calpropertiesof DNAbinding proteins:gene32proteinof T4andEscherichiacoliunwinding protein.Biochemistry 14:5485-5491.
2.Anderson,R.A., Y.Nakashima,andJ. E.Colemn.1975. Chemical modifications of functionalresidues of fd gene 5 DNA-binding protein. Biochemistry14:907-917. 3.Boyer,S.H., A. N.Noyes, M. L.Boyer,andK.MUT.
1973. Hemoglobina chains in apes. Primarystructures
and thepresumptivenatureof back mutation inanormally silent gene. J. Biol. Chem. 248:992-1003.
4. Burny, A., C. Bruck, H. Chatrenne, Y. Cleuter, D.
Dekegel, J. Ghysdael, R. Kettmann, M. Ledercq, J. Leonen, M.Mammerickx,and D.Portetefle. 1980. Bovine leukemia virus: molecularbiologyand epidemiology,p. 231-289. InG.Klein(ed.), Viraloncology.RavenPress,
NewYork.
5. Essex, M. 1980.Felineleukemia and sarcomaviruses,p. 205-229.InG. Klein(ed.), Viral oncology. RavenPress,
NewYork.
6. Ferrer, J.F.1979. Bovineleukosis: natural transmission and principles of control. J. Am. Vet. Med. Assoc.
175:1281-1286.
7. Ferrer, J. F., S. J. Kenyon, and P. Gupta. 1981. Milk of dairy cows frequently contains a leukemogenic virus. Science213:1014-1016.
8. Gilden,R. V., C. W.Long,M.Hanson,R. Toni,H. P.
Charman,S.Orozlan,J.M.Miller,and M.J. VanDer
Maten.1975.Characteristics of the majorinternalprotein
ANALYSIS OF NBPs OF BLV AND FeLV 185
andRNA-dependent DNApolymerase of bovine leuke-miavirus.J.Gen. Virol. 29-.305-314.
9. Gupta,P.,adJ. F. Ferrer. 1980. Detection of a precur-sor-likeproteinofbovineleukemia virus structural poly-peptides in purified virions. J. Gen. Virol. 47:311-322. 10. Headerso, L.E., T. D. Copend, and S. Oroszlan. 1980.
Separationof aminoacidphenylthiohydantoinsby high-performance liquid chromatographyonphenylalkyl sup-port. Anal.Biochem. 102:1-7.
11. Henderson, L. E., T. D. Copeand,G. W. Smythers,H. Marquardt,andS.Oroszian. 1978.Amino-terminal amino acid sequenceand carboxyl-terminal analysis of Rauscher murine leukemia virus glycoproteins. Virology 85:319-322.
12. Henen, L. E., T. D. Copeland, R. C. Sowder, G. W. Smythers,andS.Oroszlan. 1981.Primarystructureof the low molecular weight nucleic acid-binding proteins of murine leukemiaviruses. J. Biol. Chem. 256:8400-8406. 13. Henderson, L. E., R. C. Sowder, and S. Oronzlan. 1981. Protein andpeptide purification byreversed-phase high pressurechromatography using volatile solvents, p. 251-260. InD. T.Liu, A. N. Schechter, R. Heinrikson, and P. G.Condliffe (ed.), Chemical synthesis and sequencing of peptides and proteins.Elsevier/North-Holland, Inc., New York.
14. Jarrett, 0. 1971.Virology and host range of feline leuke-miavirus. J. Am. Vet. Med. Assoc. 158:1032-1039. 15. Khan, A. S., andJ.R.Stephe. 1977. Feline leukemia
virus:biochemical andimmunological characterization of gaggene-coded structural proteins. J. Virol.23:599-607. 16.Laeminmli, U. K. 1970. Cleavage ofstructural proteins during the assembly of the head of bacteriophage T4. Nature(London)227:680-685.
17. Long, C. W., L. E. Hendeson, and S. Oroszlan. 1980. Isolation and characterization of low-molecular-weight DNA-binding proteins from retroviruses. Virology 104:491-496.
18. Mammerl_kx, M., D. Portetelle, and A. Burny. 1981. Experimental cross-transmissions of bovine leukemia vi-rus (BLV) between several animal species. Zentralbl. Veterinaermed. 28:69-81.
19. McDonald, H. C., and J. F. Ferrer. 1976. Detection, quantitation and characterization of the major internal virion antigen of the bovine leukemia virus by radio-immunoassay.J.Natl.Cancer Inst. 57:875-82. 20. Mdbonald, H. C., D. C. Graves, and J. F. Ferrer. 1976.
Isolation andcharacterizationof anantigen of the bovine C-type virus.Cancer Res. 36:1251-1257.
21. Olpin, J. L., and S. Oroszlan. 1980. Rapid stepwise solubilizationandpurificationof typeC retrovirus struc-turalproteinsbyextractionwith organic solvent. Anal. Biochem. 103:331-336.
22. Oroszlan, S., T. D. Copeand, L. E. Henderson, J. R. Stephenson, and R. V. Gilden. 1979. Amino-terminal se-quenceof bovine leukemia virus majorinternal protein: homology with mammalian type C virus p30 structural proteins.Proc. Natl.Acad.Sci. U.S.A. 76:2996-3000. 23. Oroszlan, S., and R. V. Gilden.1980. Primarystructure
analysis ofretrovirus proteins,p. 299-344. InJ. R. Ste-phenson (ed.), Molecularbiology of RNA tumor viruses. AcademicPress, Inc., New York.
24. Oroaz_an, S., L. E.Henderson, J. R.Stephenson, T.D. Copeland, C. W. Long, J. N. Ible, and R.V.GUiden. 1978. Amino- and carboxyl-terminal amino acid sequences of proteins coded by gag gene of murine leukemia virus. Proc. Natl. Acad.Sci. U.S.A. 75:1404-1408.
25. Orszlan,S., M. G. Su dharn,T. D.Copeland,V.S. Kalyanaraman, R. V.Gilden,andR.C.Gallo. 1982. Pri-mary structureanalysis of the major internalproteinp24 of human type C T-cell leukemia virus. Proc. Natl. Acad. Sci. U.S.A. 79:1291-1294.
26. Polesz,B. J., F. W.Ruscettl,A. F.Gazdar, P. A. Bunn, J.D.Minna, and R. C.Galb.1980.Detection and isola-tionoftype C retrovirus particles from fresh and cultured lymphocytesofapatientwithcutaneousT-cell
on November 10, 2019 by guest
http://jvi.asm.org/
186 MORGAN, COPELAND, AND OROSZLAN
ma.Proc. Natl. Acad.Sci. U.S.A. 77:7415-7419. 27. Rickard, C. G., J. E. Post, F. Noronha, and L. M. Barr.
1969. Atransmissable virus-induced lymphocytic leuke-mia of thecat.J.Natl.Cancer Inst. 42:987-1014. 28. Rickard,C. G., J.E. Post, F.Noronha,andL.M. Barr.
1973. Interspecies infection by feline leukemia virus: serialcell-free transmission in dogs of malignant
lympho-mainduced by feline leukemia virus,p.102-112. In R. M. Dutcher and L. Chieco-Bianchi (ed.), Unifyingconcepts of leukemia. Karger, Basel.
29. Symlngton, J., M. Green,andK. Brakmann.1981. Im-munoautoradiographic detection of proteins after electro-phoretic transfer from gels to diazo-paper: analysis of adenovirus encoded proteins. Proc. Natl. Acad. Sci.
U.S.A.78:177-181.
30. Tarr, G. E., J. F. Beecber, M. Bell, and D. J. McKean. 1978.Polyquarternary aminespreventpeptide loss from
sequenators.Anal.Biochem. 84:622-627.
31. Uckert, W., and V.WunderHch. 1979. Proteins of bovine leukemiavirus: p15 is the major phosphoprotein. Acta Biol. Med.Germ. 38:35-42.
32. Van Der Maaten, M. J., and J. M. Milher. 1976. Serologi-calevidence of transmission of bovine leukemia virusto
chimpanzees. Vet. Microbiol. 1:351-357.
33. Van Der Maaten, M. J., J. M. MfLwer, and A. D. Bootbe. 1974.ReplicatingtypeC virus particles in monolayer cell cultures of tissues from cattle with lymphosarcoma. J. Nail. Cancer Inst. S2:491-494.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/