0022-538X/79/06-0674/09$02.00/0
Nucleotide Sequence
Deletions Within the Coding Region for
Small-t Antigen
of
Simian
Virus 40
GUIDO VOLCKAERT,'JEANFEUNTEUN,2 LIONEL V.CRAWFORD,3 PAULBERG,4 AND
WALTERFIERSI*
Laboratoryof Molecular Biology, StateUniversity of Ghent, B-9000 Ghent,Belgium';Institut deRecherches
ScientifiquessurkCancer,94800Villejuif,France2;Department of Molecular Virology, Imperial Cancer
Research Fund, London WC2A 3PX,England";andDepartment of Biochemistry, Stanford University
Medical Center, Stanford, California 94305w
Received for publication 28 November 1978
Simian virus40earlymutants withdeletions mapping in the 0.53-0.60 region
havebeen sequenced by the Maxam and Gilbert approach. All these deletions
affect the small-t gene. The size of the shortened small-t-related polypeptides
produced by several of themutantshasbeen compared with the molecular weight
as deduced from the nucleotide sequence. There was good agreement for the
mutants dl890, d1891, and d12102. For d12121 and dl2122 the small-t-related
proteinwasconsiderably larger thanexpected. It ispossible to explain this result
onthe basis of thenucleotidesequence:thenormalsplicingeventofthesmall-t
mRNA stilloccurs,butasthedeletion shifts the reading frame, translationofthe
small-t-relatedpolypeptidecontinues beyond thesmall-tsplice,butinadifferent
reading frame thanlarge-T. Mutants d1883, d1884, and d12112have lost one of
the small-t splicing boundaries, and no (or minute amounts of) small-t-related
protein has been observed in mutant-infected cells. The possible relationship
between splicingandtransportofpolyadenylic acid-containing mRNAfromthe
nucleustothecytoplasm in vertebratecellsisdiscussed.
Theearly regionof the simianvirus 40(SV40)
genome extends counter-clockwise from about
position 0.66 to 0.17 on the standard
physical
mapofSV40DNA(13,23).It codes foratleast
twoproteins,small-t and
large-T
antigens (4,
18,21), whicharetranslatedfromtwo
different,
butoverlapping,mRNA's(1).
The early region has been characterized by only one complementation group of
tempera-ture-sensitive mutants: tsA (14).
However,
Shenketal. (24)described another class of
early
regionmutants, whichwereconstructed by
in-troducing deletions in the 0.60-0.53 segment of
theviral DNA. Thesemutantsresemble inmany
aspects thepolyomahr-tmutantsisolatedbyT. Benjamin and co-workers(7,9,28).Themutants
still producenormallarge-T,butthesizeofthe
small-tprotein is affectedbythedeletion(4).In
permissive cellstheygrowonlyslightlylesswell
thanwildtype, but whentestedunder
appropri-ate conditions their transforming ability is
greatly reduced (2, 3, 6,25).
Examination of thenucleotidesequence of the
beginning of theearlyregion and theN-terminal
amino acid oflarge-Tandsmall-t showed that both proteins initiate at the same AUG start
codon,correspondingtomapposition0.649
(nu-cleotidesE80-82,numberingsystemasinref.8),
and have a common amino-terminal sequence
(19, 30).An openreadingframe extendingtothe
information for a UAA termination signal at
0.547(E602-604) allows the deduction ofa
174-amino-acids-long polypeptide. Considering the universality of the genetic code and the
alloca-tion ofthe firstboundary of the splice for
small-tmRNAtoE 606 (22),wehave concluded that
this deduced polypeptide sequence witha
mo-lecularweight of 20,503 does indeed represent
the small-tantigen (30).
The isolation of the aforementioned viable
deletionmutants andthe occurrenceoftriplets
correspondingtoterminationcodons in all three
reading frames in the region 0.53-0.55 of the
DNA message strand suggested that the seg-ment 0.60-0.53doesnotencode information for
large-T. Since neither of theuninterrupted
cod-ing regionsflankingthissegmentwould suffice
to specify a94,000-dalton large-T protein, and
considering the existenceofpeptidescommon to
large-T and small-tantigen, anintragenic
splic-ingevent washypothesizedfor thelarge-Tgene
(4).Direct characterization oftwoearlymRNA
species (1) in addition to nucleotide sequence
information in theearly region (30)clarified the
splicingoflarge-TmRNA andestablishedmore
precise boundariesfor theinterveningsequence.
674
on November 10, 2019 by guest
http://jvi.asm.org/
NUCLEOTIDE SEQUENCE OF SV40 SMALL-t MUTANTS 675
Large-T antigenis coded forbytwononadjacent
DNA segments: onefrom 0.65 (E 80) to about
0.60 andasecond from about 0.53 to the
infor-mation foraterminationcodon at 0.17(E 2550).
Analysis of the small-t mRNA showed that it
also has aspliced structure, lacking onlya
rela-tively short sequencearoundmap position0.54
(1). Considering the characterization of the
ge-nomesof small-t deletionmutantsby digestion
with restrictionenzymes(6, 24),andconsidering
also the terminationsignalsin the different
po-tential reading frames (8,30),it can be concluded
that the intervening sequence in the large-T
genecan start no further than nucleotide E 380
and endno sooner than E 670 (Fig. 3). As
dis-cussed before (30), the real boundaries cannot
be much removed from these limits. Indeed,
according to more recent results, the large-T
spliceextends from E 326 to E 671(22).
SV40DNA contains aunique TaqI cleavage siteat 0.565mapunits.Taking advantageofthis
feature,several laboratories have isolated
addi-tional mutants with deletions in the 0.60-0.53
region by selection forTaqI-resistantderivatives
(6, 25). These mutants were characterized by
restrictionenzymemappingandbytheir biolog-ical properties. We have sequenced the DNA
region spanning the deletion in mutants from
the800series(24)andfrom the 2100 series (6),
which were derived from SV40 wild-type 830
and fromlarge-plaque (LP)strain SV-1 ofSV40,
respectively. Thimmappaya and Shenk (29)
have obtained identical nucleotide sequence
re-sults for the series d1800mutants.
MATERIALS AND METHODS
Preparation of viral DNA. DNA from the 800
series of dlmutants waspreparedasdescribed
previ-ously (31). Extraction of viral DNA from the 2100
seriesof dlmutantsis describedby Feunteunet al.
(6).
Degradationof DNA withrestrictionenzymes.
Hinfl and TaqIrestriction endonucleaseswere
puri-fied in the laboratory accordingto standard
proce-dures. HaeIII and MboII endonucleases were
pur-chased from NewEngland Biolabs (Beverly,Mass.).
Digestionof DNA andseparationof restriction
frag-ments was asdescribedpreviously (31).
Labelingof DNAfragments.The restriction
di-gestof viral DNAwastreated with 1yg ofbacterial
alkaline phosphatase for45min at37°Cin the
pres-enceof 0.1% sodiumdodecylsulfate, extracted twice
withphenol, andprecipitated with ethanol. A
three-fold molarexcessof[y-YP]ATP, prepared according
tothe method ofGlynn andChappell(12),wasadded,
andtheprecipitate wasdried down. The residuewas
redissolved in20
Ad
of0.02MTris-hydrochloride (pH7.6)-0.01 M MgCl2-0.025 M 8-mercaptoethanol, and
0.5,l-(1.5 U) of T4polynucleotide kinase (P-L
Bio-chemicals,Milwaukee,Wis.)wasadded.Thereaction
mixture was incubated for 25 min at 37°Cand was
then loadedonto a 5% polyacrylamide slab gel
con-taining 0.04 M Tris-hydrochloride, 0.02 M sodium
acetate, and 0.002 M EDTA (pH 7.8). After
electro-phoresis, the bands were located by autoradiography
and eluted. Thelabeled DNA fragments could then be
cleaved with another restriction enzyme.
DNA sequencing. The sequencing technique of
Maxam andGilbert (16) was used with thefollowing
modifications: only 1
iLg
of carrier DNA was usedduring thechemicaldegradation reactions, and 4 ug of
tRNAwasadded after the chemical degradation. The
digestswerefractionated on 10%polyacrylamidegels
with dimensions of 90by30by0.2 cmand
autoradi-ographed at -70°C with intensifying screens from
CAWO(Schrobenhausen, F.R.G.) (15).
RESULTS
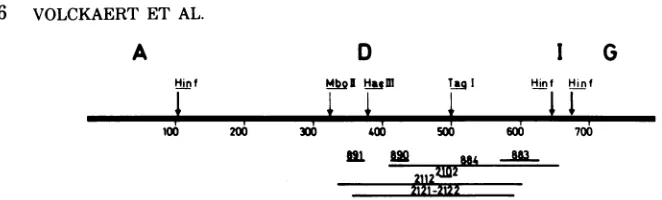
Strategy for sequence analysis. The Hinfl fragment D encompasses the 0.53-0.60 region (Fig. 1), and the following approach was used to
sequence the segmentof this fragment relevant
to eachmutant DNA. The wild-type or mutant
Hinfl fragment D was labeled at both 5' ends
with T4polynucleotide kinase and
[y-32P]ATP
and cleaved with a suitable endonuclease to
generate a singly labeled subfragment which
contained the deletion and which could be
se-quenced by the Maxam and Gilbert approach.
The particular enzyme used for this second
cleavagewasHaeIII for d1884 and d12102, MboII
for dl2112, dl2121, and dl2122, and TaqI for dl883, d1890, and wild-type large plaque. For analysis of d1891 the viral DNA was cleaved
with TaqI, 32P-labeled, and subsequently
di-gested with Hinfl. The resulting subfragments
weresequenced by the procedure of Maxam and
Gilbert (16).
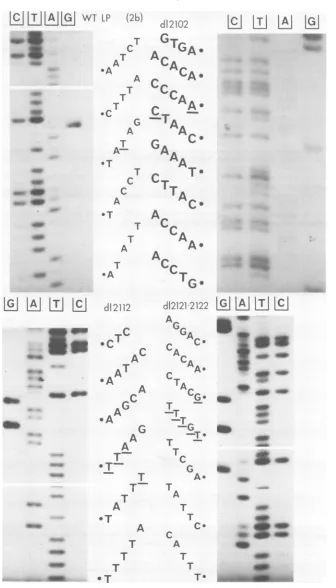
Mutantsof the800series. Figure 2a shows
the chemical degradation pattem of the DNA
region encompassing the deletion in mutants
d1883, dl884, dlI90, and dl891. The number of base pairs deleted from each mutant DNA is compiledinTable 1. The deletion indl890 and
in dl891 (27 and 25 residues, respectively), is
located closer to theorigin or5'-proximal part
of the early mRNA, whereas the deletions in
dl883 and dl884 are more counterclockwise and
include the termination triplet of the small-t
gene. dl884 hasby far the largest deletion: 247
base pairs. The deletion ofd1883 comprises 57
basepairs. Because the original d1883 stock was
suspected of being contaminated by wild-type
virus(see Discussion),the sequence analysis was
repeated on a
dl83
stock which was againplaque purified and which no longer gave any
small-t-relatedpolypeptide; the results were the
same. Adl883 stock obtained by rescue from a
transformedcelllinealso gave an identical
pat-tern.
Mutants ofthe2100series.Figure 2b shows
VOL. 30,1979
on November 10, 2019 by guest
http://jvi.asm.org/
676
A
Hinf
I
D
bl
1
I
G
Hinf Hinf
11
100 20 300 T00 6 700
89 mS
Uw
8832112~21
FIG. 1. Schematic representation of the location of deletions in the 0.53-0.60 region. The heavy horizontal
line representswild-type SV40 DNA and is calibrated in units of 100 base pairs. Restriction cleavage sites
thatwereused in the sequence determinations (see Results) are indicated by vertical arrows above the DNA
line;Hinfrestriction fragments are noted above this line by capital letters. Deletions are represented by light
horizontal lines.
thegel pattern of wild-typeSV-1 (LP) and
mu-tantfragmentsofd12102,-2112, -2121, and -2122.
Thewild-typeLPpattern showsonlyone
differ-ence in the sequence of thisregion when
com-pared with wild-type strain776: atnucleotideE
601 anA.T basepairoccurs, which isaC.Gin
the776sequence. Thismutation,whichdoes not
change the primary amino acid sequence of
small-t antigen,mayonly representadifference
between virus stocks. d12102 ismissing 15base
pairs, from E 488 to E 502. Remarkably, this
deletion is asymmetric: all residues removed
duringconstruction of thismutant werelocated
atthe left of the TaqI site (alternatively, it is
notexcluded that themutant was notgenerated
by theexonuclease treatment, but wasalready
present in thestock).
Bothd12121 and d12122 have lost thesegment
from E 361 to E 594. However, restriction
en-zyme analysis showed that d12122 gave a less
shortened Hinfl fragmentD. A closerinspection
of the DNAsequence revealedthatdl2122 has
an additional
point
mutation (G C -* T-A) in theHinflrecognition site between fragmentsDandI,resultingin alarger(fused) fragment.The
possibility that Hinfl fragment I is duplicated
canbe ruledout,because theAluI fragment C,
whichincludes theHinflfragment I,isidentical
inboth mutants (data notshown). Another
in-terestingpointisthattheligationof the mutant
DNAhasnotbeenstraightforward: there is an
inserted A-A-A-Csequence intheDNA message
strand. It isunclear how thissequencearose. The mutantd12112 has lost the segment be-tween E 339and E 607 butcontains an insertion of two A-residues. Hence a series of three A-residues hasbeenelongatedto five. The addition couldpossiblybecaused bysomeslippageof the
polymeraseduring therecircularizationevent. It
should be noted that thesephenomenawere not observed in the deletion mutants of the 800 series.
DISCUSSION
Thedeletions in the 800 series and 2100 series
mutantDNAs aresummarizedinFig. 3.
Knowl-edge of theexactposition andextent of the lost
segment allows us to compare the size of the
small-tfragmentsproducedin vivoand the
trun-catedproteins predicted from themutant DNA
sequences;the differencesaretabulatedinTable
1.The sizeof these small-t-related polypeptides
hasbeen determinedbyelectrophoresisin
poly-acrylamide gels after immunoprecipitationwith
serum from animals bearing SV40-induced tu-mors (5, 6, 17, 20, 21, 25). Since about the first
half of thesmall-tproteinisalsopresent in
large-T antigen, both proteins may share one (or
more) antigenic determinants. Paucha and
Smith (20) have also shown that analysis via
immunoprecipitation does notleadto selective
loss of small-t-related polypeptides. Based on
theresultsinTable 1,itispossibletoassign the
mutants to different classes. Sequence data
about thecorresponding regioninpolyoma virus
DNAandamutantwith adeletion thereinhave
also beenreported recently (10, 11,26).
Mutantswithanin-phasedeletion in the
small-t gene. The deletionindl890andd12102
comprises 27 and 15 base pairs, respectively,
within the small-t coding sequence; hence the
respectivemutantsmall-tproteinswilllacknine
andfiveamino acids.This is in excellent
agree-ment with the decrease in size of the
small-t-related fragment actually observed (4-6, 25)
(Table 1). It is noteworthy that a deletion of
only five amino acids reduces drastically the
transforming ability; in bothcases the deletion includes one of thecysteine-rich clusters which occurtwice inSV40 and also inpolyoma
small-tantigen (10). Sleighet al. (25) presumed that
all the deletions in mutants they had isolated were"in phase", becausetheyobserved agood
correlation of deletion size and decrease in
mo-lecular weight of the resulting small-t-related
polypeptide. However, the fact that we found
onlytwoin-phasedeletionsoutofeightmutants
sequencedindicates that suchin-phasedeletions
offer noselectivebiological advantage.
Mutants with "out-of-phase" deletions
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.72.403.53.153.2]NUCLEOTIDE SEQUENCE OF SV40 SMALL-t MUTANTS 677
producing a small-t protein of a size pre-dictable onthe basisof the location of the deletionin DNA. The deletion of d1891 starts
at E 352 and comprses 25 base pairs. Hence,
after thedeletion thecoding sequence proceeds
inanotherreadingframe until it terminates with
aTGA tripletatE432-434.Theprotein specified
by this coding sequence would be 109residues
long and have amolecular massof 12,443, i.e.,
8,060 daltonsshorter than wild-type small-t. A
small-t-related polypeptide is present in cells
infected withd1891; its decrease in sizerelative
towild-typesmall-thas been estimatedas6,500
to7,000daltons(6, 17). The agreement between
thepredictedand observed decrease in size may beregardedasreasonably good, especiallysince
the loss of most of thecysteineresidues from the
mutant small-t may greatly alter its mobility
behavior on sodiumdodecylsulfategels.
Mutants without-of-phase deletions pro-ducing atruncated small-t with a greater thanexpectedchainlength.Thesmall-t
pro-teinsynthesized by d12121 and d12122 is
consid-erably larger than the expected size based on
the extent of the deletion. Both mutants have
an identical deletion, and nucleotide E 594
marks the end of themissingDNAsegment; this
is only seven nucleotides before the wild-type
small-t termination codon.Sincefouradditional
nucleotides have beeninserted, the deletion is
not amultipleofthree,andtherefore thereading
frame changesphase and the stop signal at E
602-604 no longerfunctions. However, another
TAAtripletoccursverysooninthe newreading frame, viz., at E 607-609. Based on the DNA sequence, the mutant small-t would have a
mo-lecular weight of 11,367, which is considerably
lower than the molecularweightestimate of the
small-t-related polypeptide actually observed
(6). However, the segment spliced out of the
small-t mRNA is 66 nucleotides long, from E
606 to E 671 (22); it starts just after thenormal
small-t TAA termination signal andbefore the
second TAA codon (at E 607), which would
terminate the small-t-related polypeptide.
Hence the splicing event eliminates a series of
terminationcodons from the mRNA andpermits
translationtoproceed into the regioncoding for
the second part of large-T antigen, but in a
differentreading frame, longenough to code for
about 5,900 daltons of protein. As aresult, the
small-t-related protein terminates with a TAG
codonatE821-823. It seems very likely,
there-fore, that the same splice which in wild type
occursafter thecoding region of the small-t gene
is actuallylocated inside the coding region for
the d12121-2122-directed, small-t-related
poly-peptide. On this basis, and considering the
boundaries of the deletion in the DNA and of
the splice in the RNA, one would predict a
fragment that is 3,395 daltons shorter than
wild-type small-t, in reasonable agreement with the
2,000 to 2,500 daltonsobserved directly (6, 17).
The explanation is further strengthened by the observation of May et al. (17), who found that
translation in vitro of the mutant cytoplasmic
early mRNA results in the synthesis of a 17K
protein (asobserved invivo), whereas a nuclear
mRNA extractgave rise to both a 17K andan
11K derivative. According to the model
pro-posed above, the latter would constitute the
translation product of the prespliced mutant
mRNA, and its size corresponds indeed with what one would expect from the nucleotide se-quence.
Mutantsproducing no small-t fragments
invivo. No small-t-related fragment has been
detected in vivo after infection with deletion
mutantsdl883 and d12112 (5, 6, 17). The
small-t that was previously found in d1883-infected
cells has now been shown toresult from
contam-ination with wild-type virus (5). The situation
withd1884 islessclear.Sleigh etal. (25) could
detect (a small amountof)asmall-t-related
poly-peptide that was 3,500 daltons shorter than
wild-type small-t. AlsoKhoury (personal
communi-cationcited in ref.29) observeda12,000-dalton
polypeptide in cells infected with d1884.
Craw-ford and O'Farrell (5) could detect a
small-t-relatedpolypeptideof 15,000daltonsonlyafter
alkylation. On the other hand, Paucha and
Smith (20) did notfind putative fragments
re-latedtosmall-t ind1884-infected cells,
irrespec-tive oflabeling conditions. The reason for the
different results isnotknown,butnevertheless,
wheneverasmall-t-relatedprotein ofa
molecu-larweightin agreement with thatpredicted from
the d1884 DNA sequence was detected, its
amount wasconsiderablyreduced.
Acommonfeature of this class of mutants is
that the deletion extends beyond the small-t
terminationtriplet. Assumingthat thesplice of
thesmall-t mRNA extends fromE 606toE671,
all threedeletion mutantseliminate one of the
boundaries of the spliced sequence. Mayet al.
(17) and Paucha and Smith (20) showed that
the absence of a truncated small-t protein in
cells infected with thesemutants is due tothe
lack of corresponding cytoplasmic mRNA.
Moreover, translation in vitro of a nuclear
ex-tractdidyield a small-t fragmentwhose
reduc-tion inmolecularweightwasingood agreement
with the decrease predictedfromthe DNA
se-quence (17; E. May, personal communication).
It therefore appears that mutant premessage
RNAispresentinthe nucleusbut is notspliced
and not transported to the cytoplasm. Hence
splicingcould be an obligatoryevent for
trans-VOL. 30,1979
on November 10, 2019 by guest
http://jvi.asm.org/
LT
.
L
_Gc
.j T
Ac
CG
cG
C
T,
LIl
L-r
*{.JW
I
:_:
-C..
GA
CA..
T~~~~~~~~~~~S
e_T~~~~~~~~~'
T
T~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~X.:i
Q~~~~~~~'
dl884
A
C
G A
T
T G A c
G
A c
A
C A
L!j
Lqj
LTJ
Lq
oMe ~ ~ le
*4.4
IW.
ap
F-p
T
A
FIG. 2. Chemicaldegradationpatternscoveringthe relevantarea of each deletionmutantDNA. (More extensivegelpatternswhich substantiatethesequencedeductionswerealsoobtained [datanotshownJ.)(a)
mutantsfrom the 800series;(b) wild-typeLPandmutantsfrom the 2100 series. TheDNA restriction fragment bearing the deletionwaslabeledatonlyone5' endanddegraded accordingtothe procedure ofMaxamand Gilbert(16). The base specificity of each reaction is indicated atthe top of each lane (note that the lane referredtoasTalsoshows Cbands; theAlanealsoweakly reveals the C residues). The nucleotidesequence
isshownalongside eachautoradiogram. Nucleotides that denote the boundaries of the deletion, the additional
nucleotides ind12112, d12121, andd12122, and thesingledifference between wild-type LP and wild-type 776 areallunderlined.
Wa') o1890
C
A.
AeGA
TT G
AcG
A -c
-G TA
C
A T
T T
T0
T
C
T
T T
A A
To;
d1883I9j
rGT
_sk
A
A
C A
T q
A C A
A
I A
T
A C
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.110.425.31.620.2]NUCLEOTIDE SEQUENCE OF SV40 SMALL-t MUTANTS 679
WWTJ[J
WT
LP
(2b)
U
_00 T
_*Ao.
A*A A
d12102
GTG
AcA.
A
-
CA
CA
CA.0
TT
a
*C
C-G
TA
A
G^
A
AA
_T r%
To
c c
A
.p.ap
S.:
TTA
C.
*T
AA
T C
T
A
A
aA
T A:
[GI [51
[T]
L9]
U;
4t,
a.w
a,
d12112
cTC
A
C
AT
9A
A
GC
*
AA
G
A
A
sIF--T
AT
T-d12121
-2 122
A
GA
CC
-AC
AC
A
Cs
A
T A
._
_rm
T
T
TAc
Go
TT
-T
T-T
T Te
C
G _b
A'
T
A
T T
C.
C
._o
.... V O -
S
-4ma a.w
00
4S
e
0
.0 40 Ta.
T
e
T0 - - 5 a
i-FIG. 2b
[9iWLT]JA
[G1
I.:
VW
-VOL. 30,1979
t;c
CA
IN la,Ac
c
G
a_A O
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.84.420.69.657.2]TABLE 1. Deletionmutantsin the0.53-0.60region
Decrease in mol. wt. based on: Ammno acids No. of
nucleotides
Additional
Virus beforestart deleted amino acids Total Observed
of deletion DNA sequence small-t
frag-ment
WT - - - 174 -
-883 164 57 3 167 842
-884 111 247 3 114 7,356
890 111 27 1 +53b 165 1,055 1,000
891 91 25 18 109 8,060 6,500
2102 136 15 33" 169 631 500C
2112 86 269(-2) 0 86 10,471
-2121 94 234(-4)d 5 99 9,136' 2,000
2122 94 234(-4)d 5 99 9,136' 2,500
aThe occurrence ofasmall-t-related polypeptide in
dl884-infected cells
isunclear, as discussed in the text.bUnderlined numbers refertoaminoacids read in thesameframeaswild type.
'The decrease in sizewasoriginally reported as 1,500 daltons (7), but this estimate has been reevaluated (J.
Feunteun, unpublished data;seealsoref. 17).
d Numbersin parentheses refer to additional nucleotides present in the mutant DNA (as a result of the
ligationevent?),butnotinthewild-typeDNA.
'Adecreaseof 3,395 daltons is calculated when the small-t mRNA splicing event (which occurs here in the
coding sequence of this mutant) is taken into account (see the text).
A CT.GAG.GTA.TTT.GCT.TCT.TCC.TTA.A AT.CC T.GG T.G T T.G A T.GC A.ATGITAC.T G C.A A A.CAA.T GG.CC T.
38 .
- 2121-2122
GAG.TGT.GCA,AAG.A A A.ATG.T CT.G CT.AAC.T GC.A TA.TGC.TTG.CT G.T GC.TTA.CTG.AGG.ATG.AAG.CAT.
445
84
G AA.AAT.AGA.A AA.TTA.T AC.AGG.AAA.GAT.C C A.CT T.G TG.TG G.G TT.GAT.T GC.TAC.TGC.TTC.GAT.T G C.
508 2102 _
T T T.A GA.AT G.T G G.T T T.G GA.CTT.G AT.CT T.TGT.G AA.GG A.A CC.T TA.CTT.CTG.T G G.T G T.G AC.AT A.A T T.
t,71
GGA.CAA.ACT.ACC.TAC.AGA.GAT.TTA.AAAG.CTC GC TAAAATATAAAATATTTTAAGTGTATAATG
6,34 2121- 22 A AA C
884_
1611TAAACTACTGATTCTAAIIT611161TGTATTTTG:IA6TITCCCA.AC C.TAT.GGA.ACT.GAT.GAA.
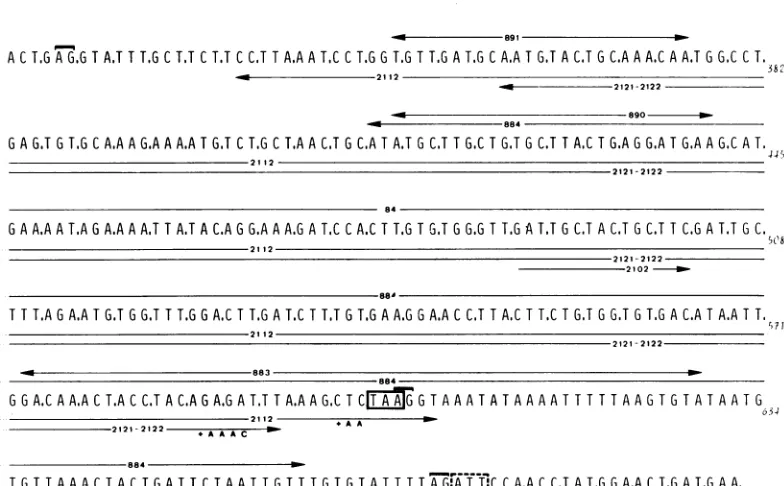
FIG. 3. SV40mutantswith deletions inthe 0.53-0.60region ofthe DNA. The nucleotidesequenceofthe
DNA message strand(samepolarityasearly mRNA)is shown(8).Theextentofthe deletionsofthe800series
mutantsisrepresented by horizontalarrowsabove thesequence, andarrowsbelow the sequence indicate that ofthe 2100seriesmutants.Nucleotide additions observed in the2100seriesmutants(see text)arenoted below
theirrespectivedeletionrepresentation.The boxed TAA tripletisthe termination codonofthe small-t gene.
The codon boxed in broken lines is thetheoreticallyearliestpositionwherelarge-Tcanpickupagain after
thesplice. TheA-G sequences thatarecovered withabracketdenote thesplicingsitesofthelarge-T (E326
andE671) and small-t (E606andE671)genesasreported byReddyetal.(22).
680 VOLCKAERT
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.65.459.63.219.2] [image:7.505.67.459.331.574.2]NUCLEOTIDE SEQUENCE OF SV40 SMALL-t MUTANTS 681
port of polyadenylic acid-containing mRNA
from the nucleustothecytoplasm in vertebrate
cells. If so, the presence ofa small amount of
small-t-related polypeptide in d1884-infected
cells couldoriginatebysomeleakage of the
pre-mRNAthrough the nuclear membraneor,
alter-natively, by the occurrence ofanother splicing
event onthesamemRNA.
Deletion mutants and splicing. All
mu-tantsused in thisstudy produceapproximately normalamountsoflarge-T antigen. Theregion
spliced outof the large-T mRNAhas been
re-portedtoextendfrom E326 toE671 (22).Since
thedeletions in thesemutantsfall withinthese
boundaries, we conclude thatmost of the
nu-cleotide sequence in the intervening region of the large-Tgene isunnecessary for thecorrect splicing of the large-T mRNA. Some of the
deletions go rather close to these crossover
points (e.g., d12112 at 13 nucleotides from the
first crossoverandd1884at 12nucleotides from
thesecond). These resultssuggestthat the
sig-nals forsplicingarerestricted to alimited area
around thecrossoversite.
Thespliceof thesmall-t mRNA extends from
E606 to E 671 (22) and only the mutantsd1883,
d1884, and d12112 eliminate part of thisregion.
These deletions interfere with thesplicingof the
small-t mRNA, and so far no small-t-related
polypeptide (orcorresponding mRNA)hasbeen
detected in the cytoplasm, except for d1884,
wheresmallamountsof therelevantpolypeptide
have been occasionally detected, as discussed
above. This suggests thatsplicingcould play a
role in thetransportofapolyadenylic
acid-con-taining mRNA from the nucleus to the
cyto-plasmorinitsstabilityinthecytoplasm.All the
other mutantsproduce a detectable
small-t-re-lated polypeptide whose size corresponds to
whatonewouldexpect onthebasis of the DNA
deletion. It is of interest that, for the mutant
d12121-2122,thereadinggoesintoanother frame and, as aresult, the normal small-tsplicenow
occursinside thecoding region for the
small-t-relatedmutantpolypeptide. Itfollows from this
that theway anmRNAissplicedisindependent
oftheway itisbeing used for translation. The
fact that about normal amounts of large-T
mRNA arepresent in all these mutants shows
that theshortsplicepresent insmall-t mRNA is
not anobligatoryprecursor for the normallarge
splice in large-TmRNA. An analogous
conclu-sion may also be relevant for the relationship
betweenSV40 19Sand16SlatemRNA's.
All SV40 mRNA's are spliced, either before,
beyond,orwithin thecodingregion(8,22). We
know of noexample of avertebrate, cellular or
viral, polyadenylic acid-containing mRNA that
reaches the cytoplasm without undergoing at
leastonesplicingevent (onepossibleexception
is tumor virus 35S RNA, which apparently
reaches thecytoplasm partly intactafter
injec-tion intonuclei; 27). Nevertheless, the link
be-tweensplicing andtransport from the nucleus to
the cytoplasm could be more complex and/or
indirect, since Paucha and Smith (20) found that
inseveral mutants (d1885, d1890) the amount of
cytoplasmic mRNA coding for the
small-t-re-lated polypeptidewasmarkedlyreduced relative
towildtype.
ACKNOWLEDGMENTS
We thank A. Van de Voorde for helpful discussions, Jose
Vander Heyden for expert technical assistance, and T. Shenk for communicating hisresults to us prior to publication.
This research wassupported by grants from the Kanker-fonds of the AlgemeneSpaar- en Lijfrentekas (ASLK) and the Geconcerteerde Akties of the Belgian Ministry of Science.
LITERATURE CITED
1. Berk, A. J., and P. A.Sharp.1977.Spliced earlymRNAs of simian virus40.Proc.Natl. Acad. Sci. U. S. A. 76: 1274-1278.
2. Bouck, N., N. Beales, T.Shenk,P.Berg, andG. Di Mayorca. 1978. New region of the simian virus 40 genomerequiredforefficient viral transformation. Proc. Natl. Acad. Sci. U. S. A.75:2473-2477.
3. Cole, C. N.,T.Landers,S. P.Goff,S.
Manteuil-Bru-tlag,and P.Berg.1977.Physicaland genetic charac-terization of deletionmutantsof simian virus40
con-structed in vitro.J. Virol. 24:277-294.
4. Crawford,L.V., C.N.Cole,A. E.Smith,E.Paucha,
P.Tegtmeyer,K.Rundell, and P. Berg. 1978. Or-ganization and expression ofearlygenes of simian virus 40.Proc. Natl. Acad. Sci. U. S. A.75:117-121.
5. Crawford,L. V.,and P. Z. O'Farrell. 1979.Effect of
alkylationonthephysical propertiesof simian virus 40 T-antigenspecies.J. Virol.29:587-596.
6. Feunteun, J., M. Kress, M.Gardes,and R. Monier. 1978.Viable deletion mutants in theSV40 early region. Proc.Natl. Acad. Sci. U. S. A. 75:4455-4459.
7. Feunteun, J.,L. Sompayrac,M.Fluck, and T.
Ben-jamin.1976.Localizationof genefunctions inpolyoma
virusDNA. Proc. Natl. Acad. Sci. U. S. A. 73:4169-4173.
8. Fiers, W., R.Contreras, G. Haegeman, R. Rogiers, A. Van deVoorde,J. VanHerreweghe,H. Van Heu-verswyn, G. Volckaert, and M. Ysebaert. 1978. Complete nucleotide sequence of SV40 DNA. Nature (London) 273:113-120.
9. Fluck, M. M., R. J. Staneloni, and T. L. Benjamin. 1977.Hr-t and ts-A: twoearly gene function of polyoma virus.Virology77:610-624.
10.Friedmann, T., R. F. Doolittle, and G. Walter. 1978. Aminoacid sequence homology between polyoma and SV40 tumour antigens deduced from nucleotide se-quences. Nature(London) 274:291-293.
11.Friedmann, T.,P.LaPorte, and A. Esty. 1978. Nucleo-tidesequence studies ofpolyomaDNA.J. Biol. Chem. 253:6561-6567.
12 .Glynn,I.M., and J. B.Chappell. 1964. A simple method
for thepreparation of 'P-labeled adenosine triphos-phate of highspecific activity. Biochem. J. 90:147-149.
13. Khoury,G., M. A. Martin, T. N. Lee, K. J. Danna, and D.Nathans.1973. Amap of simian virus 40 transcrip-tion sites expressed in productively infected cells. J. Mol. Biol.78:377-389.
14.Lai,C.-J.,and D.Nathans. 1975. A map of
temperature-VOL. 30,1979
on November 10, 2019 by guest
http://jvi.asm.org/
sensitive mutants of simian virus 40.Virology66:70-81. 15.Laskey, R. A., and A. D. Mills. 1977. Enhanced auto-radiographic detection of'Pand In usingintensifying
screensandhypersensitized film. FEBS Lett. 82:314-316.
16.Maxam,A. M., andW.Gilbert.1977. Anewmethod for
sequencingDNA. Proc.Natl. Acad. Sci. U. S. A. 74: 560-564.
17. May,E., M.Kress, and P.May.1978.Characterization of twoSV40earlymRNAs andevidence foranuclear
"prespliced"RNAspecies.Nucleic AcidsRes. 5:3083-3099.
18. Paucha,E., R.Harvey,R.Smith,and A.Smith 1977. Thecell-free synthesisofSV40Tantigen,p. 189-198. InP.May,R.Monier,and R. Weil(ed.), Earlyproteins ofoncogenicDNAviruses. INSERMColloquium,vol. 69.INSERM,Paris.
19. Paucha,E.,A.Mellor,R.Harvey,A. E.Smith,R. M.
Hewick, and M.D.Waterfield.1978.Large and small
tumor antigens fromsimian virus 40 have identical amino termini mappingat 0.65mapunits. Proc. Natl. Acad.Sci. U. S.A. 75:2165-2169.
20. Paucha, E., and A. E. Smith. 1978. The sequences
between0.59 and0.54map units onsimianvirus40
DNAcode fortheuniqueregionofsmall-tantigen. Cell 15:1011-1020.
21. Prives, C., E. Gilboa,M. Revel, and E.Winocour.
1977.Cell-freetranslationof simian virus 40early mes-senger RNA coding for viral T-antigen. Proc. Natl. Acad.Sci. U. S. A. 74:457461.
22. Reddy,V.B.,B.Thimmappaya,R.Dhar,K. N. Sub-ramanian,B. S.Zain,J.Pan,P. K.Ghosh,M.L. Celma, and S. M.Weissman. 1978.Thegenome of
simian virus 40. Science 200:494-502.
23. Sambrook, J., B. Sugden, W. Keller, and P. A. Sharp. 1973.Transcription of simian virus 40. III. Mapping of "early" and "late" species of RNA. Proc. Natl. Acad. Sci. U. S. A. 70:3711-3715.
24. Shenk, T. E., J. Carbon, and P. Berg. 1976. Construc-tion and analysis of viable deleConstruc-tion mutants of simian virus 40. J. Virol. 18:664-671.
25. Sleigh, M. J., W. C. Topp, R.Hanich, and J. F. Sam-brook. 1978. Mutants of SV40 with an alteredsmall-t protein arereduced in theirabilitytotransform cells. Cell 14:7948.
26. Soeda, E., and B. E. Griffin. 1978.Sequence from the genome of anon-transformingmutant ofpolyomavirus. Nature (London) 276:294-298.
27. Stacey, D. W., and H. Hanafusa. 1978.Nuclear
conver-sion ofmicroinjectedavianleukosis virionRNA into an envelope-glycoprotein messenger. Nature (London) 273:779-782.
28. Staneloni, R. J., M. M.Fluck,and T.L Benjamin.
1977.Host-rangeselectionoftransformation-defective hr-t mutants ofpolyomavirus.Virology77:598-609. 29. Thimmappaya, B., and T. Shenk. 1979. Nucleotide
sequence analysisof viable deletion mutantslacking
segments of thesimian virus 40 genomecoding for small tantigen. J. Virol. 30:668-673.
30. Volckaert, G., A.Van deVoorde, and W.Fiers.1978.
Nucleotidesequence of thesimianvirus 40small-t gene. Proc. Natl. Acad. Sci. U. S. A.75:2160-2164. 31. Yang, R., A. Van de Voorde, and W. Fiers. 1976.
Cleavagemap of the simian virus 40 genome by the
restrictionendonuclease III of Haemophilus aegyptius.
Eur. J. Biochem. 61:101-117.
on November 10, 2019 by guest
http://jvi.asm.org/