Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Marking and Gene Expression by a Lentivirus Vector in
Transplanted Human and Nonhuman Primate CD34
⫹
Cells
DONG SUNG AN,
1ROBERT P. WERSTO,
2BRIAN A. AGRICOLA,
2MARK E. METZGER,
2STEPHANIE LU,
1RAFAEL G. AMADO,
3IRVIN S. Y. CHEN,
1*
ANDROBERT E. DONAHUE
2Hematology Branch, National Heart, Lung, and Blood Institute, Rockville, Maryland,
2and UCLA AIDS Institute
1and
Department of Microbiology and Immunology and Molecular Genetics and Department of Medicine,
3University of
California, Los Angeles, Los Angeles, California
Received 28 July 1999/Accepted 27 October 1999
Recently, gene delivery vectors based on human immunodeficiency virus (HIV) have been developed as an
alternative mode of gene delivery. These vectors have a number of advantages, particularly in regard to the
ability to infect cells which are not actively dividing. However, the use of vectors based on human
immuno-deficiency virus raises a number of issues, not the least of which is safety; therefore, further characterization
of marking and gene expression in different hematopoietic lineages in primate animal model systems is
desirable. We use two animal model systems for gene therapy to test the efficiency of transduction and marking,
as well as the safety of these vectors. The first utilizes the rhesus animal model for cytokine-mobilized
autologous peripheral blood CD34
ⴙcell transplantation. The second uses the SCID-human (SCID-hu)
thy-mus/liver chimeric graft animal model useful specifically for human T-lymphoid progenitor cell reconstitution.
In the rhesus macaques, detectable levels of vector were observed in granulocytes, lymphocytes, monocytes,
and, in one animal with the highest levels of marking, erythrocytes and platelets. In transplanted SCID-hu
mice, we directly compared marking and gene expression of the lentivirus vector and a murine leukemia
virus-derived vector in thymocytes. Marking was observed at comparable levels, but the lentivirus vector
bearing an internal cytomegalovirus promoter expressed less efficiently than did the murine retroviral vector
expressed from its own long terminal repeats. In assays for infectious HIV type 1 (HIV-1), no
replication-competent HIV-1 was detected in either animal model system. Thus, these results indicate that while lentivirus
vectors have no apparent deleterious effects and may have advantages over murine retroviral vectors, further
study of the requirements for optimal use are warranted.
A number of groups have recently exploited the substantial
available data regarding human immunodeficiency virus type 1
(HIV-1) molecular biology and pathogenesis to develop
vehi-cles for gene delivery based on HIV-1 and other lentiviruses (3,
9, 27, 30, 39, 40, 42, 43, 45, 47, 48, 51, 54). The simplest of these
vectors consists of the minimal
cis
-acting sequences required
for HIV-1 replication. Other HIV-1 genes are expressed in
trans
with a packaging plasmid. The gene to be expressed,
either a therapeutic gene or a reporter gene, is expressed as
part of an internal transcriptional unit inserted between the
long terminal repeats (LTRs) of the vector. Other
modifica-tions of these vectors have resulted in the generation of
tat
-dependent or inducible expression of the gene to be expressed.
A
tat
-dependent vector may be particularly suitable for HIV-1
disease since it will be expressed only in the presence of HIV-1
infection (5, 9). In some cases, HIV-1 vectors are themselves
capable of inhibiting HIV-1 replication (5, 10, 15). Other
ther-apeutic genes have been inserted into HIV-1 vectors, and it is
likely that there will be increasing emphasis on the potential
use of these vectors for treatment of various human conditions
(22, 24, 29, 36, 52). The primary advantage of lentivirus vectors
is that by virtue of nuclear localization signals present in HIV-1
proteins that are part of the preintegration complex (11, 21, 26,
56), these vectors can efficiently infect some nondividing cells
(39, 41, 43, 48, 54), provided they reside or progress through at
least the G1b
state of the cell cycle (32). Other retroviral
vectors based on murine retroviruses require passage of the
cell through mitosis in order to integrate (34, 38, 49). Another
advantage of lentivirus vectors is that they have evolved to
efficiently replicate in human cells. However, the latter factor
also underscores the need to carefully assess the properties of
lentivirus vectors, particularly those derived from HIV-1, prior
to use in humans.
Several animal model systems have been used to evaluate
retroviral vector delivery systems (2, 6, 8, 12, 13, 18, 41, 43). Of
particular relevance to lentivirus vector systems is the ability to
test transduction, reconstitution, gene expression, and marking
and ultimately therapeutic efficacy in model systems for human
and/or nonhuman primates. The rhesus macaque model
sys-tem has been shown to be amenable to transduction of CD34
⫹cells and transplantation and is arguably the closest model
system for human gene therapy (7, 18, 20, 28, 53, 57). In
addition, for some diseases such as AIDS, the rhesus macaque
will also allow for testing of therapeutic efficacy against simian
immunodeficiency virus (17, 23). Use of this transplantation
model, however, is expensive and does not lend itself to the
type of experimental manipulation required to test multiple
variables as do other small-animal model systems, such as the
SCID-NOD (50) and SCID-human (SCID-hu) chimeric mouse
models (37). In the SCID-NOD system, human CD34
⫹cells
are used to reconstitute an irradiated SCID mouse, resulting in
the production of human granulocytes, monocytes, and B cells
(16, 25). In the SCID-hu system, human CD34
⫹cells can be
transplanted following irradiation of a chimeric human
thy-mus/liver (thy/liv) organ previously implanted into the mice to
mimic human thymopoiesis (2, 6, 8). Typically, the SCID-NOD
mouse is used to analyze non-T-lymphoid progenitor CD34
⫹* Corresponding author. Mailing address: 10833 LeConte Ave.,
11-934 Factor Building, Los Angeles, CA 90095-1678. Phone: (310)
825-4793. Fax: (310) 794-7682. E-mail: [email protected].
1286
on November 9, 2019 by guest
http://jvi.asm.org/
cell transplant, whereas the SCID-hu mouse is utilized to
in-vestigate T-lymphoid progenitor CD34
⫹cell transplant.
In this study, we used the rhesus macaque and SCID-hu
system to investigate the properties of transplanted CD34
⫹cells transduced with a lentivirus vector bearing an internal
cytomegalovirus (CMV) promoter. We demonstrate that the
lentivirus vector can result in multilineage hematopoietic cell
marking. However, comparative studies using SCID-hu mice
indicate that the levels of gene expression are substantially
lower than that of a murine retroviral vector using the LTR as
a promoter. We find no evidence for replication competent
HIV-1 in either transplanted rhesus macaques or SCID-hu
mice.
MATERIALS AND METHODS
HIV-1 vector construction and production.An HIV-1-based vector, pHR⬘ -CMV-luciferase (43), was modified to contain the enhanced green fluorescent protein (EGFP) cDNA with expression driven by a CMV promoter (HR⬘CMV EGFP). A packaging plasmid for the HIV-1-based vector, pCMVR8.2DVPR, was derived from pCMVR8.2 (43, 44) by deleting thevprgene from nucleotide positions 5625 to 5731 by oligonucleotide-directed mutagenesis. Numbering of nucleotides starts at the 5⬘end of HIV-1 NL4-3 provirus (1). All vector stocks
were generated by calcium phosphate-mediated transfection of 293T cells (American Type Culture Collection, Manassas, Va.). 293T cells were cultured in Dulbecco’s modified Eagle medium (DMEM) with 10% calf serum, 100 U of penicillin per ml, and 100g of streptomycin per ml. 293T cells (2⫻107) were plated on 175-cm2flasks in 25 ml of the medium and transfected the following day with 5 g of pHCMVG, 12.5g of pCMVR8.2DVPR, and 12.5g of HR⬘CMVEGFP for the HIV-1-based vector. For the murine leukemia virus (MLV)-based vector (SR␣LEGFP with expression of EGFP from the 5⬘LTR), 5 g of pHCMVG (14), 12.5g of pSV⌿⫺env⫺MLV (33), and 12.5g of SR␣LEGFP were used (5). At 8 h posttransfection, the medium was replaced with 35 ml of fresh medium. At 36 and 60 h posttransfection, the medium was harvested, centrifuged at 1,500 rpm for 5 min (Sorvall RT 6000B; Ivan Sorvall, Norwalk, Conn.), and filtered through a 0.45-m-pore-size filter. Further vector concentration was achieved by ultracentrifugation at 50,000⫻gfor 90 min at 4°C. The pellet was resuspended in Iscove’s modified Dulbecco’s medium with 10% fetal calf serum (FCS), 100 U of penicillin per ml, and 100g of strepto-mycin per ml overnight at 4°C. The vectors were concentrated 100-fold and kept in liquid nitrogen until use. Stocks of vectors were titrated by infecting HeLa cells (105) with various amounts of the virus and analyzing for EGFP expression by flow cytometry on day 3 postinfection. The titers of vectors were 108infectious units/ml for the HIV-1 vector and 2⫻107infectious units/ml for the MLV-based vector.
Rhesus leukapheresis procedure.Young adult rhesus macaques (Macaca mu-latta) that were serologically negative for simian T-cell lymphotropic virus, sim-ian immunodeficiency virus, simsim-ian AIDS-related type D virus, and herpes B virus were used. Experimental animals were quarantined and housed in
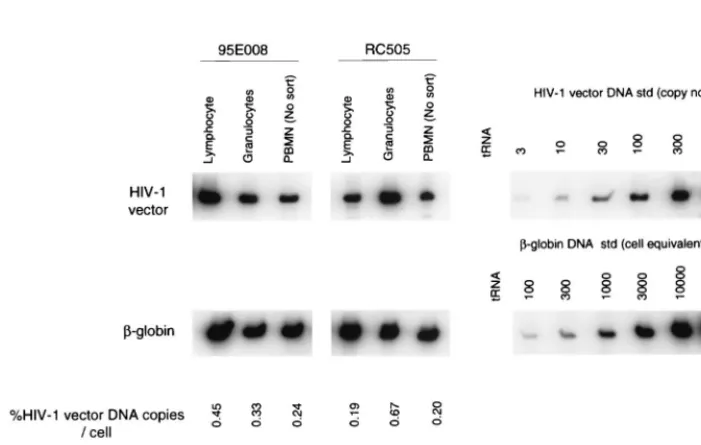
[image:2.612.54.550.85.157.2]accor-FIG. 1. PCR analysis of rhesus macaque cell fractions following transplant. DNA from rhesus macaque granulocytes, lymphocytes, and whole PBMN were analyzed for HIV-1 vector transduction by PCR 10 weeks after reconstitution of rhesus macaque 95E008 and 16 weeks after reconstitution of rhesus macaque RC505. Granulocyte and lymphocyte populations were sorted from PBMN to, respectively, 98 and 99% purity, based on forward and side scatter. HIV-1 vector DNA-specific signal was compared with that of amplified-globin DNA sequences to determine the number of vector copies per cell (%HIV-1 vector DNA copies/cell, calculated as number of HIV-1 vector DNA copies/number of cell equivalents⫻1/10⫻100). For PCR amplification, 10-fold less DNA was used for-globin DNA PCR in order to obtain quantitative-globin DNA signals. Quantitative HIV-1 vector DNA and-globin DNA standards (std) were assayed along with DNA from a nontransduced rhesus (negative control) in parallel. No signals were detected for the negative control (data not shown). Percent EGFP expression was determined by flow cytometric analysis at the same time point as PCR. tRNA served as the negative control for the PCR assay.
TABLE 1. Outcome of G-CSF- and SCF-mobilized peripheral blood transplantation using immunoselected CD34
⫹cells
Animal (cytokine[s] used invitro, frequency of transductiona)
No. of CD34⫹ cells reinfused
(107)
No. of CD34⫹cells/
kg (107)
% EGFP-expressing
cells reinfusedb Day leukocyte count⬎1,000 Day platelet count⬎50,000
95E008 (SCF, BID)
4.0
1.0
3.2
12
Did not fall below 50,000
95E009 (SCF, BID)
1.6
0.4
13.2
15
16
RC504 (SCF
⫹
IL-6, SID)
1.4
0.4
9
11
11
RC505 (SCF
⫹
IL-6, SID)
1.6
0.4
6
11
Did not fall below 50,000
aSID; once a day; BID; twice a day.
bPercentage of CD34⫹cells expressing EGFP on the day of infusion.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.127.478.437.659.2]dance with federal guidelines (44a) and the policies set by the Veterinary Re-search Program of the National Institutes of Health. The protocols were evalu-ated and approved by the Animal Care and Use Committee of the National Heart, Lung, and Blood Institute.
Rhesus macaques received a combination of granulocyte colony-stimulating factor (G-CSF; 10g/kg of body weight/day) and stem cell factor (SCF; 200 g/kg/day) (both kindly provided by Amgen, Inc., Thousand Oaks, Calif.) by subcutaneous injection for 4 days. Mobilized (i.e., cytokine-stimulated) leuka-pheresis cell product (mPB) was collected from the peripheral blood by using a CS3000 Plus blood cell separator (Baxter Healthcare, Fenwal Division, Deer-field, Ill.) with a single, small-volume chamber and other modifications made to the fluid path of the CS3000 Plus blood cell separator (19). This allowed
leuka-pheresis procedures to be performed on rhesus macaques weighing⬍5 kg. Four rhesus macaques were transplanted. All four animals received a total dose of 1,000 R of total-body␥irradiation over 2 consecutive days prior to reinfusion of the transduced cells. Peripheral blood was collected on the designated days and evaluated by PCR analysis or flow cytometry for EGFP DNA or expression, respectively.
Immunoselection of nonhuman primate CD34ⴙcells.mPB CD34⫹cells were isolated according to the manufacturer’s instructions by magnetic selection using a biotinylated CD34⫹ antibody (clone 12.8; CellPro, Inc., Bothell, Wash.), streptavidin MicroBeads, and MACS separation columns (Miltenyi Biotec, Inc., Auburn, Calif.). Purity following the immunoselection procedure was routinely ⬎95% as assessed by flow cytometry using an ELITE flow cytometer (Beckman Coulter Corp., Miami, Fla.) after staining with an allophycocyanin-conjugated anti-CD34 monoclonal antibody (MAb; clone 563; a kind gift from Gustav Gaudernack, Institution of Transplantation Immunology, Rikshospitalet, The National Hospital, Oslo, Norway, with allophycocyanin conjugation performed by Molecular Probes, Inc., Eugene, Oreg.).
Transduction of immunoselected rhesus macaque CD34ⴙcells.
Immunos-elected CD34⫹cells were transduced with the vesicular stomatitis virus G pro-tein (VSV-G)-pseudotyped HIV-1 vector (HR⬘CMVEGFP) either once or twice a day for 48 h for all four animals on RetroNectin (BioWhittaker, Walkersville, Md.)-coated non-tissue culture-treated six-well plates (Becton Dickinson Lab-ware, Franklin Lakes, N.J.) according to the manufacturer’s instructions. Trans-ductions for two animals (95E008 and 95E009) were performed twice a day in SCF alone (100 ng/ml) and 8 protamine sulfate (8g/ml); the other two animals (RC504 and RC505) were transduced with the same HIV-1 vector only once a day at the same multiplicity of infection for the 48-h period in medium supple-mented with SCF at 100 ng/ml, interleukin-6 (IL-6) at 50 ng/ml, and protamine sulfate at 8g/ml. The HIV-1 vector was used at a multiplicity of infection of approximately 5. EGFP expression was measured by flow cytometry using a standard filter setup for fluorescein (525-nm band-pass filter).
Quantitative PCR assay.Each PCR amplification was performed as described elsewhere (58). In brief, to detect HIV-1 vector sequences, one of the oligonu-cleotide primers for each pair used was end labeled with32P, and 25 ng was included in the reaction (usually 5⫻106to 1⫻107cpm). The second oligonu-cleotide primer was not labeled, and 50 ng was incorporated into each reaction. Each reaction mixture contained 0.25 mM each of the four deoxynucleoside triphosphates, 50 mM NaCl, 25 mM Tris-HCl (pH 8.0), 5 mM MgCl2, 100 mg of bovine serum albumin per ml, and 1.25 U ofTaqDNA polymerase (Promega, Madison, Wis.). The reaction mixture was overlaid with 25l of mineral oil and then subjected to 25 cycles of denaturation for 1 min at 94°C and polymerization for 2 min at 65°C. The reaction was performed on a Perkin-Elmer thermocycler. Amplified products resulting from the PCR were analyzed by electrophoresis on 6% nondenaturing polyacrylamide gels and visualized by direct autoradiography of the dried gels. Quantitative analysis of the amplified products was performed with a PhosphorImager (Molecular Dynamics, Sunnyvale, Calif.), and data were analyzed with the ImageQuaNT program (Molecular Dynamics). The nucleotide sequences of the oligonucleotide primers (M667 and AA55) used for HR⬘CMV EGFP DNA detection were derived from the nucleotide sequence of the HIV-1 LTR as previously described (58). A pair of oligonucleotide primers comple-mentary to the first exon of the human-globin gene (58) was used in each reaction mixture in PCR analyses to normalize the total amount of rhesus macaque cellular DNA present. During PCR amplification, labeled -globin-specific oligonucleotides were incorporated into the reaction at 5⫻106to 1⫻ 107cpm.
[image:3.612.88.262.72.489.2]Quantitation of HIV-1 vector DNA during PCR amplifications was performed by analyzing a standard curve of dilution of HR⬘CMVEGFP plasmid DNA digested withHindIII, which does not cleave vector sequences. This DNA was diluted in 0.01 g of rhesus macaque peripheral blood mononuclear cell
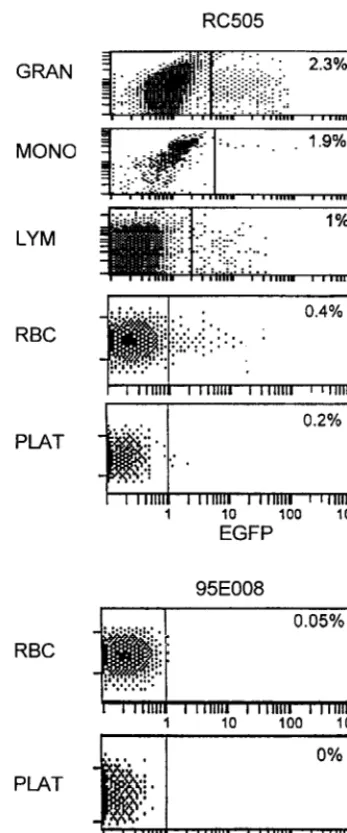
FIG. 2. Flow cytometric analysis of rhesus macaque PBMC at 30 weeks. Circulating leukocytes at 30 weeks were evaluated by flow cytometry in animals transduced with the HIV-1 vector (HR⬘CMVEGFP). Granulocytes (GRAN), monocytes (MONO), lymphocytes (LYM), erythrocytes (RBC), and platelets (PLAT) were gated according to size (forward scatter) and granularity (log 90° scatter) and analyzed for EGFP expression. Results for HIV-1 vector-transduced rhesus macaques RC505 and 95E008 are shown. EGFP expression was detected by flow cytometry in all the subpopulations in RC505. The other three trans-planted animals had EGFP expression in granulocytes, monocytes, and lympho-cyte populations but did not demonstrate any detectable fluorescence in eryth-rocytes and platelets. Data for rhesus macaque 95E008 are shown. Thexaxis is log fluorescent intensity of EGFP; theyaxis represents the gated population based on forward and side scatter (logarithmic fluorescence intensity).
TABLE 2. EGFP expression after activation of PBMN
aAnimal
Stimulated PBMN Unstimulated PBMN
% EGFP [
3H]thymidine
incorporation
(cpm) % EGFP
[3H]thymidine
incorporation (cpm)
95E008
0.03
113,650
0.02
1,964
95E009
0.09
53,135
0.21
291
RC504
0.05
88,973
0.03
362
RC505
0.15
35,913
0.13
170
Mock
0.00
64,415
0.00
325
aRhesus macaque PBMN (106) were obtained from reconstituted rhesus
ma-caques at 23 weeks (98E008 and 98E009) or 18 weeks (RC504 and RC505) postreconstitution. These cells were cultured for 3 days in the presence of human IL-2 and immobilized anti-monkey CD3 MAb. As a control, PBMN were cul-tured in parallel in the absence of growth-stimulating agents. [3H]thymidine
incorporation was measured by pulse-labeling cells for 6 h on day 3.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.311.551.581.679.2](PBMN) DNA per ml. The copy number of HIV-1 vector included in the standard curve ranged from 3 to 1,000. Standard curves for rhesus macaque -globin DNA were obtained by amplification of 0.001 to 0.3g of rhesus macaque cellular DNA (100 to 30,000 cell equivalents) from rhesus macaque PBMN.
Transduction and immunoselection of gene-transduced human fetal liver-derived CD34ⴙcells by flow cytometry.Human fetal liver-derived CD34⫹cells were purified from a fetal liver as previously described (6). Cells were cultured in Iscove’s modified Dulbecco’s medium with 100 ng each of IL-3, IL-6, and SCF (kindly supplied by Amgen) per ml, 20% FCS, penicillin (100 U/ml), and strep-tomycin (100g/ml). Fetal liver-derived CD34⫹cells (2⫻106) were transduced with VSV-G-pseudotyped HR⬘CMVEGFP or SR␣LEGFP vector by incubating
2 ml of a 40-fold dilution of the HR⬘CMVEGFP vector stock or 10-fold dilution of the SR␣LEGFP vector stock in the presence of Polybrene (8g/ml) at 37°C for 2 h on day 1 or 2 after CD34⫹cell purification from fetal liver, respectively. On day 3 after CD34⫹cell purification from fetal liver, vector-transduced CD34⫹cells were stained with a MAb to human CD34 (Becton Dickinson, Mountain View, Calif.) conjugated with phycoerythrin. EGFP and CD34⫹cells were sorted on a FACStarplus(Becton Dickinson). Thy/liv implants of irradiated (300 rads) animals were directly injected with 105EGFP and CD34⫹cells on day 4 after CD34⫹cell purification from fetal liver.
[image:4.612.54.551.78.417.2]Flow cytometric analysis for human thymocytes.Thymocytes were obtained by biopsy from reconstituted thy/liv implants of SCID-hu mice 4 weeks postrecon-stitution. Thymocytes were stained with a MAb to human CD1, CD3, CD4, CD5
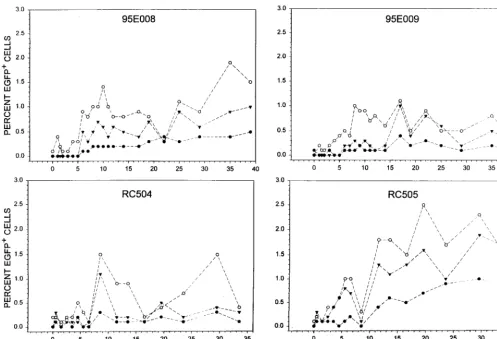
FIG. 3. Kinetics of EGFP expression following transplant. The percentage of EGFP-expressing granulocytes (E), lymphocytes (F), and monocytes () was
determined at various time points as described in the legend to Fig. 2 for all four animals receiving the lentivirus-transduced immunoselected CD34⫹cells. The percentage of EGFP-expressing cells is shown over a 30- to 40-week evaluation period.
TABLE 3. Cocultivation with activated human PBL
aSample ELISAp24
(ng/ml)
MAGI cell assay (no. of blue cells
in a field)
% EGFP expression
Before cocultivation After cocultivation
PBMN from HIV-1 vector-transduced macaque 95E008
⬍0.008
3
0.54
⬍0.001
PBMN from HIV-1 vector-transduced macaque RC505
⬍0.008
2
1.15
⬍0.001
PBMN from mock-transduced macaque 96E025
⬍0.008
2
⬍0.001
⬍0.001
HIV-1 uninfected MAGI cell culture supernatant
⬍0.008
6
NA
NA
HIV-1 (NL4-3)-infected human PBL
1,303
⬎800
NA
NA
aPBMN (107) from lentivirus vector-transduced rhesus macaques were cocultured with PHA-activated human PBL (107) for 4 weeks. Every 7 days, activated human
PBL (107) were added to the culture. At 4 weeks postcocultivation, culture supernatants were harvested, measured for p24, and subjected to MAGI assay as described
in Materials and Methods. Blue cells in a field were counted. Magnification was 100-fold (objective lens [10⫻] and eyepiece lens [10⫻]). Approximately 2,500 MAGI cells could be observed in a field when the cells were confluent. NA, not applicable.
on November 9, 2019 by guest
http://jvi.asm.org/
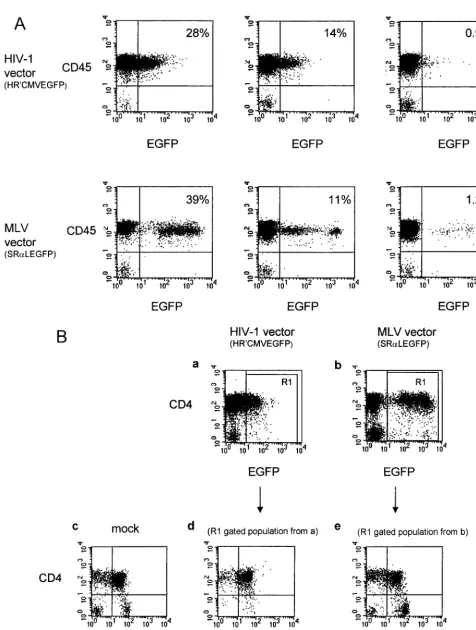
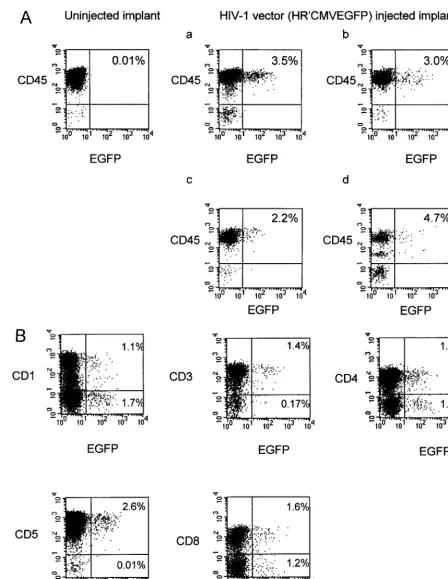
[image:4.612.54.550.612.694.2]FIG. 4. (A) EGFP expression 4 weeks after reconstitution in human CD45⫹thymocytes of reconstituted thy/liv implants in SCID-hu mice. EGFP expression at 28, 14, and 0.9% was detected in human CD45⫹thymocytes in three SCID-hu mice receiving a thy/liv transplant transduced with the HIV-based vector and at 39, 11, and 1.2% in three SCID-hu mice receiving a thy/liv transplant transduced with an MLV-based vector 4 weeks postimplant. Less than 0.2% of EGFP expression was detected in thymocytes from a SCID-hu mouse receiving nontransduced cells as a thy/liv implant (data not shown). Ten thousand events were analyzed. (B) Distribution of
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.62.538.65.695.2]CD8, or CD45 directly conjugated with phycoerythrin or periodinin chlorophyll protein (Becton Dickinson). Samples were run on a FACScan flow cytometer, and data analyzed with the CellQuest program (Becton Dickinson). Ten thou-sand events were acquired for analysis.
Cell culture for in vitro activation studies.Human thymocytes and rhesus macaque PBMN were cultured at a concentration of 106/ml in flat-bottom cul-ture plates. Culcul-ture plates were coated with goat anti-mouse immunoglobulin G (GAM; Tago, Burlingame, Calif.). GAM (10 g/ml) in phosphate-buffered saline (PBS; pH 7.4) was added to wells and incubated for 2 h at 37°C. Plates were then washed three times with PBS. Anti-human CD3 MAb (T3; 4 mg/ml in PBS; Coulter, Hialeah, Fla.) or anti-monkey CD3 MAb (1 mg/ml; BioSource Inter-national, Camarillo, Calif.) was added on to the GAM-coated culture plates and incubated at 37°C for 1 h. The immobilized GAM provides a solid phase for binding of anti-human or monkey CD3 MAb as previously described (35, 55). After washing, thymocytes obtained from thy/liv implants of SCID-hu mice or rhesus macaque PBMN were cultured in the presence of IL-2 (10 U/ml; Amgen), anti-human CD28 MAb, and immobilized anti-human CD3 MAb or anti-monkey CD3 MAb in Iscove modified Dulbecco’s medium–20% FCS supplemented with penicillin (100 U/ml) and streptomycin (100 mg/ml), respectively. For control, cells were cultured in parallel in the absence of IL-2, anti-CD28 MAb, and immobilized anti-CD3 MAb. After 3 days of stimulation in vitro, thymocytes were analyzed by flow cytometry for EGFP expression. [3H]thymidine incorpo-ration was measured by pulse-labeling cells for 6 h on day 2 as previously described (58).
Coculture of rhesus macaque PBMN and activated human PBL.PBMN (107) from lentivirus vector-transduced or mock-transduced rhesus macaques were cocultured with 107 phytohemagglutinin (PHA)-activated human peripheral blood lymphocytes (PBL) in RPMI with 10% FCS and human IL-2 (10 U/ml). As a control, 107PHA-activated human PBL infected with 1 ng of HIV-1 NL4-3 were cocultured with 107PHA-activated human PBL. Every 7 days, 107activated human PBL were added to the culture. At 4 weeks postcocultivation, culture supernatants were harvested, measured for p24 by enzyme-linked immunosor-bent assay (ELISA), and subjected to MAGI cell assay.
MAGI assay.The CD4-positive LTR-galactosidase-expressing HeLa (MAGI) cell indicator line (31) was obtained from the AIDS Research and Reference Program, Division of AIDS, National Institute of Allergy and Infectious Dis-eases, and maintained in DMEM supplemented with 10% calf serum, 100 U of penicillin G sodium per ml, 0.1 mg of streptomycin sulfate per ml, 0.15 mg of G418 sulfate per ml, and 0.1 mg of hygromycin B per ml. Cells were seeded at a density of 8⫻104per well in a 12-well plate 24 h prior to infection. Cell coculture supernatants of rhesus macaque PBMN and activated human PBL (300l) were adsorbed to cells in the presence of Polybrene (10g/ml) for 2 h at 37°C prior to the addition of 1 ml of medium. As controls, culture supernatant (300l of p24 [1,303 ng/ml] of HIV-1 NL4-3-infected human PBL and uninfected culture supernatant of MAGI cells (300l) were used. Following incubation for 2 days at 37°C, the cells were fixed and stained with 5-bromo-4-chloro-3-indolyl-D
-galactopyranoside (X-Gal) as previously described (31).
Direct injection of HIV-1 vector into thy/liv implant of SCID-hu mouse.Prior to vector injection, SCID-hu thy/liv mice were irradiated (200 rads). At 24 h (implants a and b) or 4 days (implants c and d) postirradiation, 100-fold-con-centrated HIV-1 vector was directly injected into irradiated thy/liv implants of SCID-hu mice. Approximately 50l of the vector stock was injected in each irradiated implant. The titer of the HIV-1 vector was 108infectious units/ml.
RESULTS
Transduction and transplantation of immunoselected mPB
CD34
ⴙcells in rhesus macaques.
We used an HIV-1 vector
bearing an internal CMV immediate-early promoter for
ex-pression of the gene encoding EGFP. We evaluated this HIV-1
vector in the rhesus macaque transplantation model.
Nonhu-man primate immunoselected hematopoietic growth factor
mPB CD34
⫹cells were transduced on non-tissue-coated
six-well plates treated with the recombinant fibronectin fragment
CH-296 (RetroNectin) either once or twice a day for 2
con-secutive days following collection. Because lentivirus vectors
can transduce nonmitotic cells, the CD34
⫹cells were
stimu-lated with combinations of cytokines (SCF alone or SCF plus
IL-6) that induced less cell division than combinations typically
used and may therefore induce less lineage-specific
differenti-ation. On average, 7.6% (range, 3.2 to 13.2%) of the CD34
⫹cells expressed EGFP following transduction with the
lentivi-rus vector (Table 1). This transduction efficiency is superior to
that observed for murine retroviral vectors, as most cells
cul-tured under these conditions at various time points are not yet
susceptible to murine retroviral infection (data not shown).
Presence and expression of vector in rhesus PBMN
sub-populations.
Four animals were transplanted with autologous
CD34
⫹cells transduced with the HIV-1 vector. All animals
had uneventful hematopoietic reconstitution following
total-body
␥
irradiation and autologous transplant, with leukocyte
counts returning to 1,000 cells/
l within 15 days of transplant
and platelet counts returning to greater than 50,000/
l within
11 days of transplant (Table 1). We monitored the presence of
vector in different hematopoietic lineages by
fluorescence-ac-tivated cell sorting followed by PCR analysis as well as by
direct flow cytometric analysis for EGFP in gated cell
popula-tions. As determined by PCR analysis, approximately 0.1 to 1%
of circulating leukocytes contained vector DNA in sorted
lym-phocytic and granulocytic cell fractions 10 weeks (animal
95E008) or 16 weeks (animal RC505) following transplant
(Fig. 1). In one animal, RC505, we observed a higher level of
vector DNA in the granulocyte subpopulation.
EGFP expression was detected in PBMN beginning at 2
weeks after transplant. EGFP expression at 30 weeks
post-transplant in various PBMN subpopulations is shown in Fig. 2.
EGFP is readily detected in granulocyte, monocyte, and
lym-phocyte subpopulations. As determined by the mean
fluores-cence intensity, the expression in lymphocytes was the least
efficient. Further analysis indicated that both B and T
lympho-cytes (CD20 and CD2 positive, respectively) as well as NK cells
(CD16 CD56 double positive) expressed the EGFP gene (data
not shown). The percent of cells expressing EGFP was
moni-tored over a period of 34 to 39 weeks (Fig. 3) in each of four
transplanted rhesus macaques. EGFP expression was observed
in granulocyte, lymphocyte, and monocyte subsets. Although
there was variation in the level of marking over time, we
observed a general increase in marking between 5 to 10 weeks
following transplant. In the animal (RC505) with the highest
levels of marking, the proportions increased over time to
be-tween 1 to 2% of PBMN. To date, 14 months following
trans-plant, the proportion of PBMN expressing EGFP has
re-mained relatively stable, with the highest levels ranging from 1
to 3%.
EGFP expression in erythrocytes and platelets.
Of
particu-lar interest is the observation that low but significant levels of
EGFP could be detected 30 weeks posttransplant in the
eryth-rocytes and platelets of one animal, RC505, with the highest
overall proportion of marked PBMN (Fig. 2). Marking of these
cell types has not previously been observed in rhesus macaques
transplanted with CD34
⫹cells transduced with a murine
ret-rovirus (murine stem cell virus [MSCV]-based) vector (46).
Since the majority of these cells would not be expected to
harbor vector DNA, we presume that we were detecting
per-sisting EGFP protein. We cannot formally exclude the
possi-bility that we were detecting EGFP only in nucleated precursor
cells. However, this appears unlikely since if it were the case,
EGFP-expressing thymocytes 4 weeks after reconstitution of thy/liv implants in SCID-hu mice. At 4 weeks after injection of human EGFP⫹CD34⫹cells, thymocytes from gene-transduced and reconstituted thy/liv implants were obtained by biopsy of SCID-hu mice. Obtained thymocytes were stained for human CD4 and CD8 markers and evaluated for EGFP expression by flow cytometric analysis. As shown in the lower panels, thymocytes found to express EGFP were gated and analyzed for CD4 and CD8 distribution. EGFP expression was detected in both CD4 and CD8 single-positive cells as well as CD4 CD8 double-positive and CD4 CD8 double-negative thymocytes. EGFP-expressing thymocytes showed a CD4 and CD8 profile similar to that of non-gene-transduced reconstituted implant controls (mock). Ten thousand events were analyzed.
on November 9, 2019 by guest
http://jvi.asm.org/
FIG. 5. EGFP expression in human thymocytes 4 weeks after direct injection of HIV-1 vector in thy/liv implants of SCID-hu mice. Prior to vector injection, SCID-hu thy/liv mice were irradiated (200 rads) to induce progenitor cell proliferation and to kill resident thymocytes. At 24 h (implants a and b) or 4 days (implants c and d) postirradiation, 100-fold-concentrated HIV-1 vector was directly injected into irradiated thy/liv implants of SCID-hu mouse. Four irradiated implants (a through d) were each injected with approximately 50l of the vector stock. The titer of HIV-1 vector was 108infectious units/ml. (A) At 4 weeks after direct injection of HIV-1 vector
into thy/liv implants, EGFP expression was analyzed in human CD45⫹thymocyte populations by flow cytometry. (B) EGFP expression was analyzed in human CD1, CD3, CD4, CD5, and CD8 thymocyte subpopulations. Representative data from implant a are shown.
on November 9, 2019 by guest
http://jvi.asm.org/
the majority of circulating erythroid precursor cells would have
to express vector, inconsistent with the frequency of marked
cells in the other hematopoietic lineages.
Activation of rhesus PBMN does not enhance the levels of
vector expression.
The mean fluorescence intensity of EGFP in
lymphocytes was about fivefold weaker than that in
granulo-cytes (Fig. 2). We tested whether activation of PBMN cells with
IL-2 and anti-CD3 MAb could enhance expression. Treatment
of rhesus PBMN with IL-2 and anti-CD3 resulted in an
ap-proximately 100-fold increase in [
3H]thymidine incorporation.
Activation of PBMN did not appear to significantly increase
the proportion of cells expressing EGFP (Table 2) or EGFP
fluorescence intensity (data not shown).
Replication-competent HIV-1 is not evident.
To date, more
than 50 weeks posttransplant, all four animals remain healthy
and have not demonstrated circulating antibody to HIV p24 by
Western blotting, circulating p24 antigen by ELISA, or
evi-dence of circulating HIV-1 virus by quantitative HIV-1
ultra-sensitive reverse transcription-PCR (data not shown). We
further confirmed that there was no evidence of
replication-competent or latent HIV-1 in the cells by coculture of 10
7rhesus macaque PBMN with activated human PBL followed by
passage and repeated coculture for 4 weeks. The supernatant
was then assayed for HIV-1 p24 by ELISA and for
competent HIV-1 by MAGI assay. No evidence for
replication-competent HIV-1 was found (Table 3).
HIV-1 vector-transduced CD34
ⴙcells can reconstitute
irra-diated SCID-hu thy/liv grafts.
We previously used human thy/
liv chimeric grafts transplanted into SCID-hu mice as a model
for progenitor cell gene therapy using murine retroviral vectors
(2, 6). This small-animal model system allows the investigation
of marking of T-cell lineages in the human thymus throughout
thymopoiesis, not easily addressed experimentally in the rhesus
macaque model. SCID-hu thy/liv chimeric grafts transplanted
3 to 4 months previously were irradiated and injected with 5
⫻
10
5human fetal liver CD34
⫹cells transduced with an HIV-1 or
MLV-based vector (SR
␣
LEGFP). Both the HIV-1 and MLV
vector-transduced CD34
⫹cells can reconstitute SCID-hu thy/
liv mice (Fig. 4). At 4 weeks after introduction of CD34
⫹cells
into thy/liv implants, the percentage of EGFP-expressing cells
ranged from 0.9 to 28% for HIV-1 vector-transduced implants
and from 1.2 to 39% for MLV vector-transduced implants (Fig.
4A). We confirmed that the EGFP-expressing cells were
hu-man cells by using anti-CD45, a marker for huhu-man
lympho-cytes. Three-color flow cytometric analysis determined that
when thymocytes were first gated on EGFP-positive cells and
subsequently analyzed for expression of CD4 and CD8, the
distribution of CD4
⫹and CD8
⫹cells for HIV-1 or MLV
vector-transduced cells was similar to that for mock-infected
cells, indicating that progenitor cells common to CD4
⫹and
CD8
⫹cell lineages were likely to have been transduced with
both vectors (Fig. 4B, compare d and e with c).
The MLV vector resulted in gene expression higher than
that of the HIV-1 based vector.
In comparing the levels of
EGFP expression for the MLV and HIV-1 vectors in
trans-duced thymocytes, we noted that the HIV-1 vector expressed
at levels of fluorescence intensity approximately 10-fold-lower
than that of the MLV vector. This difference was consistent
among reconstituted implants (Fig. 4). Although we did not
directly compare MLV and HIV-1 vector expression in rhesus
macaques, the fluorescence intensity of the HIV-1 vector in
SCID-hu thymocytes is similar to the fluorescence intensity in
PBL in the rhesus macaques when analyzed with similar flow
cytometric settings (data not shown).
Activation of thymocytes does not induce increased level of
EGFP expression.
In transplanted rhesus macaques, we found
that EGFP expression could not be further induced following
activation of PBMN (see above). We similarly tested whether
we could induce EGFP expression following activation of the
thymocytes. Thymocytes from transplanted SCID-hu mice
were stimulated in vitro with IL-2 and with anti-CD3 and
anti-CD28 MAbs for 3 days. The rate of thymidine
incorpora-tion of the stimulated cells increased approximately 100-fold
over that of nonstimulated cells. The level of EGFP
expres-sion, however, was only slightly induced following activation
with anti-CD3 and IL-2 (Table 4). These results are consistent
with our previously published studies with an MLV vector (6).
The HIV-1 vector is expressed throughout thymopoiesis.
We
previously demonstrated that a murine retroviral vector used
to transduce human CD34
⫹cells and transplanted to SCID-hu
mice was expressed in various thymocyte subpopulations
throughout thymopoiesis (6). We assessed whether gene
ex-pression directed by the HIV-1 vector was similarly expressed.
For these studies, we modified the transduction and transplant
protocol by using a new and more rapid approach to assess the
HIV-1 vector. The vector was directly injected into irradiated
thy/liv implants of SCID-hu mice that had been irradiated to
kill resident thymocytes and induce progenitor cell function. At
4 weeks after transplant of the transduced CD34
⫹cells into
thy/liv implants, EGFP expression was detected in CD45
⫹thy-mocyte populations (Fig. 5A). Similar percentages of
EGFP-expressing cells were detected in all thymocyte subpopulations
tested (CD1, CD3, CD4, CD5, and CD8), indicating vector
expression throughout thymopoiesis (Fig. 5B). These results
are consistent with that previously observed for the MLV
vec-tor (6).
Reconstituted SCID-hu implants did not show any
deleteri-ous effects.
Human thymocytes of the SCID-hu thy/liv mouse
are highly susceptible to death when challenged with infectious
replication-competent HIV-1. No toxic effects were observed
in those implants receiving the HIV-1 or MLV
vector-trans-duced CD34
⫹cells. The percentages of CD4
⫹, CD8
⫹, and
CD4
⫹CD8
⫹cells expressing EGFP remained similar to that
[image:8.612.312.552.83.198.2]of both mock-transduced and MLV vector-transduced animals
(Fig. 4B). Thus, no HIV-1-associated pathogenicity was
ob-served in the transduced animals, indicating that no
replica-tion-competent HIV-1 was present in the viral stocks or
gen-erated after transduction, consistent with the results observed
in transplanted rhesus macaques.
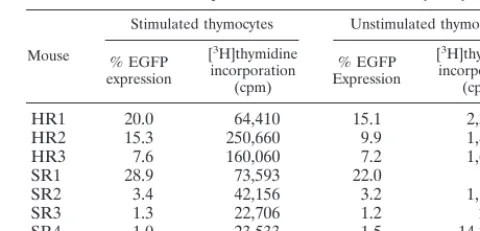
TABLE 4. EGFP expression after activation of thymocytes
aMouse
Stimulated thymocytes Unstimulated thymocytes
% EGFP expression
[3H]thymidine incorporation
(cpm)
% EGFP Expression
[3H]thymidine incorporation
(cpm)
HR1
20.0
64,410
15.1
2,302
HR2
15.3
250,660
9.9
1,884
HR3
7.6
160,060
7.2
1,635
SR1
28.9
73,593
22.0
192
SR2
3.4
42,156
3.2
1,195
SR3
1.3
22,706
1.2
241
SR4
1.0
23,533
1.5
14,226
aHuman thymocytes (106) obtained from reconstituted thy/liv implants with HIV-1 vector (HR) or MLV vector (SR) at 4 weeks postreconstitution were cultured for 3 days in the presence of IL-2, anti-human CD28 MAb, and immo-bilized anti-human CD3 MAb. As a control, thymocytes were cultured in parallel in the absence of growth-stimulating agents. After 3 days of stimulation in vitro, cells were analyzed by flow cytometry for EGFP expression. [3H]thymidine in-corporation was measured by pulse-labeling cells for 6 h on day 3.
on November 9, 2019 by guest
http://jvi.asm.org/
DISCUSSION
Our results demonstrate that successful transplantation and
marking are obtained in rhesus macaque and SCID-hu mice
that have been transplanted with nonhuman and human
CD34
⫹cells, respectively, transduced with an HIV-1-based
lentivirus vector. Infusion of myeloablated rhesus macaques
resulted in reconstitution and marking of lymphoid, myeloid,
and granulocyte lineages in all animals. In the case of one
animal with the greatest overall levels of marking, EGFP
ex-pression was observed in erythrocytes and platelets, not
previ-ously observed with MLV vectors (46). Parallel experiments
conducted with human CD34
⫹cells in SCID-hu mice
demon-strated reconstitution of thymopoiesis and evidence of vector
throughout various stages of T-cell differentiation. Other
in-vestigators have demonstrated multilineage marking in a
SCID-NOD model system (41); however, that model system
did not examine T-lineage gene transfer and marking, critical
for evaluation of efficacy and potential deleterious effects of a
vector derived from HIV-1.
Gene expression in different cell types is dependent on the
relative strength of promoters used. The HIV-1 vector uses an
internal CMV promoter, whereas the murine retroviral vector
uses the vector LTR. HIV-1 vector expression of EGFP as
monitored by the mean fluorescence intensity in lymphocyte
subpopulations was relatively weak. This level of expression
was at least 10-fold lower than that for the MLV-based vector
in SCID-hu thymocytes. Although rhesus macaque
lympho-cytes were marked, only a low fluorescence intensity of
expres-sion from the HIV-1 vector was detected following transplant,
about fivefold lower than expression in rhesus macaque
gran-ulocytes, consistent with the lower level of expression observed
in the SCID-hu thy/liv thymocytes. For both human and rhesus
macaque lymphoid populations, no increase in expression was
observed following ex vivo T-cell activation.
We cannot say with certainty whether we have successfully
transduced in rhesus macaques a pluripotent hematopoietic
stem cell, but several lines of evidence suggest that the HIV-1
vector may have transduced an early progenitor cell prior to
commitment to the myeloid or lymphoid pathway. First, the
relative degrees of marking for both lymphoid and granulocyte
compartments are similar for each rhesus macaque as
deter-mined by PCR. Second, these relative proportions have either
remained stable or even increased in certain instances 30
weeks following reconstitution (data not shown). Third, in the
animal with highest overall levels of marking, EGFP
expres-sion could be observed in multiple hematopoietic lineages,
including erythrocytes and platelets. Additional studies,
how-ever, will be required to improve transduction efficiency and
the level of EGFP expression in desired subpopulations of
hematopoietic cells.
Concerns have been raised regarding the use of vectors
derived from HIV-1 in humans (4). Since these vectors are
defective for HIV-1 envelope and generated by cotransfection
with a VSV-G-expressing envelope vector, it is highly unlikely
that a replication-competent virus could be formed from
re-combination between members of two distinct families of
vi-ruses. Nevertheless, we formally showed in both transplanted
rhesus macaques and SCID-hu mice that
replication-compe-tent HIV-1 did not result. Therefore, we believe that lentivirus
vectors in principle are suitable for use in humans; however,
further refinements in vector design including enhanced
ex-pression will be necessary before lentivirus vectors can be used
effectively for gene delivery.
ACKNOWLEDGMENTS
We thank Barrington Thompson, Earl West, the staff of Bionetics,
Inc., the Laboratory of Small Animal Surgery and Medicine, and the
staff of Jerry Zack’s laboratory for assistance in caring for the animals,
and we thank Frances Ngok for technical assistance. We also thank Liz
Duarte for assistance in preparing the manuscript.
This work was supported in part by the UCLA CFAR and NIH
grants AI39975 and AI36555.
REFERENCES
1.Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R. Willey, A. Rabson, and M. A. Martin.1986. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J. Virol.59:284–291.
2.Akkina, R. K., J. D. Rosenblatt, A. G. Campbell, I. S. Chen, and J. A. Zack. 1994. Modeling human lymphoid precursor cell gene therapy in the SCID-hu mouse. Blood84:1393–1398.
3.Akkina, R. K., R. M. Walton, M. L. Chen, Q. X. Li, V. Planelles, and I. S. Chen.1996. High-efficiency gene transfer into CD34⫹cells with a human immunodeficiency virus type 1-based retroviral vector pseudotyped with vesicular stomatitis virus envelope glycoprotein G. J. Virol.70:2581–2585. 4.Amado, R. G., and I. S. Y. Chen.1999. The promise of gene therapy within
reach? Science285:674–676.
5.An, D. S., K. Morizono, Q. Li, H. Ma, S. Lu, and I. S. Y. Chen.1999. An inducible human immunodeficiency virus type 1 (HIV-1) vector which effec-tively suppresses HIV-1 replication. J. Virol.73:7671–7677.
6.An, D. S., Y. Koyanagi, J. Q. Zhao, R. Akkina, G. Bristol, N. Yamamoto, J. A. Zack, and I. S. Chen.1997. High-efficiency transduction of human lymphoid progenitor cells and expression in differentiated T cells. J. Virol.71:1397– 1404.
7.Bodine, D. M., T. Moritz, R. E. Donahue, B. D. Luskey, S. W. Kessler, D. I. Martin, S. H. Orkin, A. W. Nienhuis, and D. A. Williams.1993. Long-term in vivo expression of a murine adenosine deaminase gene in rhesus monkey hematopoietic cells of multiple lineages after retroviral mediated gene trans-fer into CD34⫹bone marrow cells. Blood82:1975–1980.
8.Bonyhadi, M. L., K. Moss, A. Voytovich, J. Auten, C. Kalfoglou, I. Plavec, S. Forestell, L. Su, E. Bohnlein, and H. Kaneshima.1997. RevM10-expressing T cells derived in vivo from transduced human hematopoietic stem-progen-itor cells inhibit human immunodeficiency virus replication. J. Virol.71: 4707–4716.
9.Buchschacher, G. L. J., and A. T. Panganiban.1992. Human immunodefi-ciency virus vectors for inducible expression of foreign genes. J. Virol.66: 2731–2739.
10. Bukovsky, A. A., J. P. Song, and L. Naldini.1999. Interaction of human immunodeficiency virus-derived vectors with wild-type virus in transduced cells. J. Virol.73:7087–7092.
11. Bukrinsky, M. I., S. Haggerty, M. P. Dempsey, N. Sharova, A. Adzhubel, L. Spitz, P. Lewis, D. Goldfarb, M. Emerman, and M. Stevenson.1993. A nuclear localization signal within HIV-1 matrix protein that governs infec-tion of non-dividing cells. Nature365:666–669.
12. Bunnell, B. A., M. Metzger, E. Byrne, R. A. Morgan, and R. E. Donahue. 1997. Efficient in vivo marking of primary CD4⫹T lymphocytes in nonhu-man primates using a gibbon ape leukemia virus-derived retroviral vector. Blood89:1987–1995.
13. Bunting, K. D., M. Y. Sangster, J. N. Ihle, and B. P. Sorrentino.1998. Restoration of lymphocyte function in Janus kinase 3-deficient mice by retroviral-mediated gene transfer. Nat. Med.4:58–64.
14. Burns, J. C., T. Friedmann, W. Driever, M. Burrascano, and J. K. Yee.1993. Vesicular stomatitis virus G glycoprotein pseudotyped retroviral vectors: concentration to very high titer and efficient gene transfer into mammalian and non-mammalian cells. Proc. Natl. Acad. Sci. USA90:8033–8037. 15. Corebeau, P., and F. Wong-Staal.1998. Anti HIV effects of HIV vectors.
Virology243:268–274.
16. Dick, J. E., M. Bhatia, O. Gan, U. Kapp, and J. C. Wang.1997. Assay of human stem cells by repopulation of NOD/SCID mice. Stem Cells15(Suppl. 1):199–203.
17. Donahue, R. E., B. A. Bunnell, M. C. Zink, M. E. Metzger, R. P. Westro, M. R. Kirby, T. Unangst, J. E. Clements, and R. A. Morgan.1998. Reduction in SIV replication in rhesus macaques infused with autologous lymphocytes engineered with antiviral genes. Nat. Med.4:181–186.
18. Donahue, R. E., S. W. Kessler, D. Bodine, K. McDonagh, C. Dunbar, S. Goodman, B. Agricola, E. Byrne, M. Raffeld, and R. Moen.1992. Helper virus induced T cell lymphoma in nonhuman primates after retroviral me-diated gene transfer. J. Exp. Med.176:1125–1135.
19. Donahue, R. E., M. R. Kirby, M. E. Metzger, B. A. Agricola, S. E. Sellers, and H. M. Cullis.1996. Peripheral blood CD34⫹cells differ from bone marrow CD34⫹cells in Thy- 1 expression and cell cycle status in nonhuman primates mobilized or not mobilized with granulocyte colony-stimulating factor and/or stem cell factor. Blood87:1644–1653.
20. Dunbar, C. E., M. Cottler-Fox, J. A. O’Shaughnessy, S. Doren, C. Carter, R.
on November 9, 2019 by guest
http://jvi.asm.org/
Berenson, S. Brown, R. C. Moen, J. Greenblatt, and F. M. Stewart.1995. Retrovirally marked CD34-enriched peripheral blood and bone marrow cells contribute to long-term engraftment after autologous transplantation. Blood 85:3048–3057.
21.Gallay, P., T. Hope, D. Chin, and D. Trono.1997. HIV-1 infection of nondividing cells through the recognition of integrase by the importin/karyo-pherin pathway. Proc. Natl. Acad. Sci. USA94:9825–9830.
22. Gallichan, W. S., T. Kafri, T. Krahl, I. M. Verma, and N. Sarvetnick.1998. Lentivirus-mediated transduction of islet grafts with interleukin 4 results in sustained gene expression and protection from insulitis. Hum. Gene Ther. 9:2717–2726.
23. Geretti, A. M.1999. Simian immunodeficiency virus as a model of human HIV disease. Rev. Med. Virol.9:57–67.
24. Goldman, M. J., P. S. Lee, J. S. Yang, and J. M. Wilson.1997. Lentiviral vectors for gene therapy of cystic fibrosis. Hum. Gene Ther.8:2261–2268. 25. Greiner, D. L., R. A. Hesselton, and L. D. Shultz.1998. SCID mouse models
of human stem cell engraftment. Stem Cells16:166–177.
26. Heinzinger, N. K., M. I. Bukinsky, S. A. Haggerty, A. M. Ragland, V. Kewal-ramani, M. A. Lee, H. E. Gendelman, L. Ratner, M. Stevenson, and M. Emerman.1994. The Vpr protein of human immunodeficiency virus type 1 influences nuclear localization of viral nucleic acids in nondividing host cells. Proc. Natl. Acad. Sci. USA91:7311–7315.
27. Kafri, T., U. Blomer, D. A. Peterson, F. H. Gage, and I. M. Verma.1997. Sustained expression of genes delivered directly into liver and muscle by lentiviral vectors. Nat. Genet.17:314–317.
28. Kaptein, L. C., V. W. Van Beusechem, I. Riviere, R. C. Mulligan, and D. Valerio.1997. Long-term in vivo expression of the MFG-ADA retroviral vector in rhesus monkeys transplanted with transduced bone marrow cells. Hum. Gene Ther.8:1605–1610.
29. Kay, M. A.1998. Hepatic gene therapy for haemophilia B. Haemophilia 4:389–392.
30. Kim, V. N., K. Mitrophanous, S. M. Kingsman, and A. J. Kingsman.1998. Minimal requirement for a lentivirus vector based on human immunodefi-ciency virus type 1. J. Virol.72:811–816.
31. Kimpton, J., and M. Emerman.1992. Detection of replication-competent and pseudotyped human immunodeficiency virus with a sensitive cell line on the basis of activation of an integrated-galactosidase gene. J. Virol.66: 2232–2239.
32. Korin, Y. D., and J. A. Zack.1998. Progression to the G1bphase of the cell cycle is required for completion of human immunodeficiency virus type 1 reverse transcription in T cells. J. Virol.72:3161–3168.
33. Landau, N. R., and D. R. Littman.1992. Packaging system for rapid pro-duction of murine leukemia virus vectors with variable tropism. J. Virol. 66:5110–5113.
34. Lewis, P. F., and M. Emerman.1994. Passage through mitosis is required for oncoretroviruses but not for the human immunodeficiency virus. J. Virol. 68:510–516.
35. Lorre, K., A. Kasran, F. Van Vaeck, M. de Boer, and J. L. Ceuppens.1994. Interleukin-1 and B7/CD28 interaction regulate interleukin-6 production by human T cells. Clin. Immunol. Immunopathol.70:81–90.
36. Marcello, A., and I. Giaretta.1998. Inducible expression of herpes simplex virus thymidine kinase from a bicistronic HIV1 vector. Res. Virol.149:419– 431.
37. McCune, J. M., R. Namikawa, H. Kaneshima, L. D. Shultz, M. Lieberman, and I. L. Weissman.1988. The SCID-hu mouse: murine model for the analysis of human hematolymphoid differentiation and function. Science 241:1632–1639.
38. Miller, D. G., M. A. Adam, and A. D. Miller.1990. Gene transfer by retro-virus vectors occurs only in cells that are actively replicating at the time of infection. Mol. Cell. Biol.10:4239–4242. (Erratum,12:433, 1992.) 39. Miyoshi, H., M. Takahashi, F. H. Gage, and I. M. Verma.1997. Stable and
efficient gene transfer into the retina using an HIV-based lentiviral vector. Proc. Natl. Acad. Sci. USA94:10319–10323.
40. Miyoshi, H., U. Blomer, M. Takahashi, F. H. Gage, and I. M. Verma.1998. Development of a self-inactivating lentivirus vector. J. Virol.72:8150–8157. 41. Miyoshi, H., K. A. Smith, D. E. Mosier, I. M. Verma, and B. E. Torbett.1999. Transduction of human CD34⫹cells that mediate long-term engraftment of
NOD/SCID mice by HIV vectors. Science283:682–686.
42. Mochizuki, H., J. P. Schwartz, K. Tanaka, R. O. Brady, and J. Reiser.1998. High-titer human immunodeficiency virus type 1-based vector systems for gene delivery into nondividing cells. J. Virol.72:8873–8883.
43. Naldini, L., U. Blomer, P. Gallay, D. Ory, R. Mulligan, F. H. Gage, I. M. Verma, and D. Trono.1996. In vivo gene delivery and stable transduction of nondividing cells by a lentiviral vector. Science272:263–267.
44. Naldini, L., U. Blomer, F. H. Gage, D. Trono, and I. M. Verma.1996. Efficient transfer, integration, and sustained long-term expression of the transgene in adult rat brains injected with a lentiviral vector. Proc. Natl. Acad. Sci. USA93:11382–11383.
44a.National Institutes of Health.1985. Guide for the care and use of laboratory animals, rev. ed. Department of Health and Human Services publication no. (NIH) 85-23. National Institutes of Health, Bethesda, Md.
45. Parolin, C., T. Dorfman, G. Palu, H. Gottlinger, and J. Sodroski.1994. Analysis in human immunodeficiency virus type 1 vectors ofcis-acting se-quences that affect gene transfer into human lymphocytes. J. Virol.68:3888– 3895.
46. Persons, D. A., J. A. Allay, J. M. Riberdy, R. P. Wersto, R. E. Donahue, B. P. Sorrentino, and A. W. Nienhuis.1998. Use of the green fluorescent protein as a marker to identify and track genetically modified hematopoietic cells. Nat. Med.4:1201–1205.
47. Poznansky, M., A. Lever, L. Bergeron, W. Haseltine, and J. Sodroski.1991. Gene transfer into human lymphocytes by a defective human immunodefi-ciency virus type 1 vector. J. Virol.65:532–536.
48. Reiser, J., G. Harmison, S. Kluepfel-Stahl, R. O. Brady, S. Karlsson, and M. Schubert.1996. Transduction of nondividing cells using pseudotyped defec-tive high-titer HIV type 1 particles. Proc. Natl. Acad. Sci. USA93:15266– 15271.
49. Roe, T., T. C. Reynolds, G. Yu, and P. O. Brown.1993. Integration of murine leukemia virus DNA depends on mitosis. EMBO J.12:2099–2108. 50. Shultz, L. D., P. A. Schweitzer, S. W. Christianson, B. Gott, I. B. Schweitzer,
B. Tennent, S. McKenna, L. Mobraaten, T. V. Rajan, and D. L. Greiner. 1995. Multiple defects in innate and adaptive immunologic function in NOD/ LtSz-scid mice. J. Immunol.154:180–191.
51. Sutton, R. E., H. T. Wu, R. Rigg, E. Bohnlein, and P. O. Brown.1998. Human immunodeficiency virus type 1 vectors efficiently transduce human hemato-poietic stem cells. J. Virol.72:5781–5788.
52. Takahashi, M., H. Miyoshi, I. M. Verma, and F. H. Gage.1999. Rescue of photoreceptor degeneration in therdmouse by human immunodeficiency virus vector-mediated gene transfer. J. Virol.73:7812–7816.
53. Tisdale, J. F., Y. Hanazono, S. E. Sellers, B. A. Agricola, M. E. Metzger, R. E. Donahue, and C. E. Dunbar.1998. Ex vivo expansion of genetically marked rhesus peripheral blood progenitor cells results in diminished long-term repopulating ability. Blood92:1131–1141.
54. Uchida, N., R. E. Sutton, A. M. Friera, D. He, M. J. Reitsma, W. C. Chang, G. Veres, R. Scollay, and I. L. Weissman.1998. HIV, but not murine leu-kemia virus, vectors mediate high efficiency gene transfer into freshly iso-lated G0/G1 human hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 95:11939–11944.
55. Verwilghen, J., M. L. Baroja, F. Van Vaeck, J. Van Damme, and J. L. Ceuppens.1991. Differences in the stimulating capacity of immobilized anti-CD3 monoclonal antibodies: variable dependence on interleukin-1 as a helper signal for T-cell activation. Immunology72:269–276.
56. von Schwedler, U., R. S. Kornbluth, and D. Trono.1994. The nuclear local-ization signal of the matrix protein of human immunodeficiency virus type 1 allows the establishment of infection in macrophages and quiescent T lym-phocytes. Proc. Natl. Acad. Sci. USA91:6992–6996.
57. Xu, L. C., S. Karlsson, E. R. Byrne, S. Kluepfel-Stahl, S. W. Kessler, B. A. Agricola, S. Sellers, M. Kirby, C. E. Dunbar, and R. O. Brady.1995. Long-term in vivo expression of the human glucocerebrosidase gene in nonhuman primates after CD34⫹hematopoietic cell transduction with cell-free retro-viral vector preparations. Proc. Natl. Acad. Sci. USA92:4372–4376. 58. Zack, J. A., S. J. Arrigo, S. R. Weitsman, A. S. Go, A. Haislip, and I. S. Chen.
1990. HIV-1 entry into quiescent primary lymphocytes: molecular analysis reveals a labile, latent viral structure. Cell61:213–222.
on November 9, 2019 by guest
http://jvi.asm.org/