Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Biochemical Analyses of the Interactions between Human
Immunodeficiency Virus Type 1 Vpr and p6
Gag
YONCHU JENKINS,1OWEN PORNILLOS,2REBECCA L. RICH,3DAVID G. MYSZKA,3
WESLEY I. SUNDQUIST,2ANDMICHAEL H. MALIM1,4*
Departments of Microbiology1and Medicine,4University of Pennsylvania School of Medicine, Philadelphia,
Pennsylvania 19104-6148, and Department of Biochemistry2and Center for Biomolecular Interaction Analysis,3
University of Utah School of Medicine, Salt Lake City, Utah 84132
Received 13 June 2001/Accepted 2 August 2001
The nonstructural human immunodeficiency virus type 1 Vpr protein is packaged into progeny virions at significant levels (⬃200 copies/virion). Genetic analyses have demonstrated that efficient Vpr packaging is dependent upon a leucine-X-X-leucine-phenylalanine (LXXLF) motif located in the p6Gag domain of the
structural Gag polyprotein. Recombinant proteins spanning full-length Vpr (Vpr1–97) or the amino-terminal 71
amino acids (Vpr1–71) formed specific complexes with recombinant p6 proteins in vitro. Complex formation
required an intact LXXLF motif and exhibited an intrinsic dissociation constant of⬃75M. Gel filtration and cross-linking analyses further revealed that Vpr1–71self-associated in solution. Our experiments demonstrate
that Vpr can bind directly and specifically to p6 and suggest that oligomerization of both Vpr and Gag may serve to increase the avidity and longevity of Vpr-Gag complexes, thereby ensuring efficient Vpr packaging.
Human immunodeficiency virus type 1 (HIV-1) encodes a 97-amino-acid accessory protein called Vpr (an initialism for viral protein R). Although Vpr is dispensable for HIV-1 growth in culture, the protein presumably plays an important role(s) in replication in vivo since simian immunodeficiency virus derived from rhesus macaques lacking the two vpr ho-mologs (vprandvpx) replicates at reduced levels and displays attenuated pathogenicity in infected animals (12, 15). Al-though significant quantities of Vpr are packaged into budding HIV-1 particles (5, 23, 25, 26), the biological significance of this remains to be fully elucidated. For instance, the stimula-tion of postentry viral nucleoprotein complex nuclear import (11, 13), the capacity of virion-associated Vpr to induce target cell apoptosis or G2/M arrest (16, 27), and the Vpr-mediated
recruitment of the DNA repair enzyme uracil DNA glycosylase into virions (24) have each been reported. From a practical standpoint, efficient virion incorporation has enabled research-ers to target heterologous proteins into assembling virions as Vpr fusion proteins (39); this approach has been used to study the early steps of virus infection (4, 8, 36, 37), to deliver inhibitory factors to virions (38), and to tag virions fluores-cently for direct visualization during infection (30, 33).
The structural p55Gagprotein creates the viral particle and is
either directly or indirectly responsible for packaging of all of the essential components of the virion (10). Gag is translated as a multidomain polyprotein that assembles into an immature viral particle and then buds from the cell. Coincident with budding, Gag is proteolytically processed into a series of new, discrete proteins—termed matrix, capsid, nucleocapsid, and p6. These Gag-derived proteins rearrange to form the mature, infectious virus, which is characterized by the presence of a
central conical core structure. In addition to the Gag proteins, HIV-1 cores also contain the essential enzymes reverse tran-scriptase, integrase, and protease and the viral RNA genome, as well as the virus-encoded accessory proteins Vif, Nef, and Vpr (1, 19, 21, 35). Of these latter proteins, Vpr is packaged with substantially greater efficiency, with a recent estimate of the Vpr/Gag ratio being⬃1:7 (25).
The mechanism of Vpr incorporation into virus particles, as well as its retention in uncoated (postentry) cores, is not fully understood. The carboxy-terminal p6Gagregion of p55Gag
ap-pears to play an important role in Vpr packaging because deletions and mutations in p6Gagcan block Vpr incorporation,
and transfer of the p6 coding sequence to a heterologous retroviral Gag protein endows the chimeric virus particles with the capacity to package Vpr (18, 22). Genetic mapping exper-iments have established that a conserved LXXLF motif at positions 41 to 45 in p6Gagis required for efficient Vpr
pack-aging and that an alanine substitution at any of the three conserved hydrophobic residues abolishes Vpr packaging (17). Consistent with these packaging experiments, yeast two-hy-brid analyses have shown that Vpr interacts with p55Gagin a
p6Gag-dependent manner (2, 32). Similarly, in vitro binding
studies have also indicated that Vpr interacts with p55Gag,
although the dependence of this interaction on the p6Gag
re-gion was not demonstrated (2). It is noteworthy that neither of these experimental methods can eliminate the possibility that another molecule, e.g., RNA, might serve as a bridge between Vpr and p6Gag. Moreover, and somewhat unexpectedly, both
two-hybrid and biochemical analyses have indicated that Vpr either interacts weakly or is unable to interact with p6 when the rest of p55Gagis removed (6, 32). Instead, it has been reported
that Vpr can interact with the upstream nucleocapsid or matrix protein (6, 20, 28, 32). Nevertheless, in spite of the lack of an unequivocal demonstration of direct Vpr-p6 binding, it is gen-erally assumed that such interactions do occur in infected cells as a prerequisite to Vpr packaging. In an effort to validate this * Corresponding author. Present address: Department of Infectious
Diseases, Guy’s, King’s and St. Thomas’ Medical School, King’s Col-lege London, 3rd Floor New Guy’s House, GKT Guy’s Hospital, Lon-don SE1 9RT, United Kingdom. Phone: 44 0 20 7955 4472. Fax: 44 0 20 7955 2846. E-mail: [email protected].
10537
on November 9, 2019 by guest
http://jvi.asm.org/
model further, we have characterized the direct binding of Vpr to p6 in vitro by using purified recombinant proteins. By adopt-ing this approach, we have been able to define kinetic and thermodynamic details of the interaction.
Studies of HIV-1 Vpr in vitro have frequently been ham-pered by difficulties in obtaining and/or maintaining prepara-tions of soluble proteins. In the course of testing different subfragments of Vpr in a variety ofEscherichia coliexpression systems, we found that the amino-terminal 71 residues of Vpr from the YU-2 isolate (hereafter called Vpr1–71) can be
pro-duced at high levels by using a pET27b-based vector (Nova-gen). This fragment corresponds closely to a protease-resistant fragment of Vpr identified by others (41). Vpr1–71-His6 was
purified under nondenaturing conditions by using nickel che-late affinity chromatography, and the hexahistidine tag was then removed by enterokinase cleavage (this left a nonnative five-amino-acid [aspartic acid4-lysine] extension at the
pro-tein’s carboxy terminus). The resulting Vpr1–71 protein was
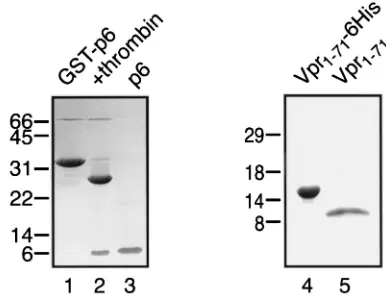
purified to near homogeneity by anion-exchange chromatog-raphy on Q Sepharose (Fig. 1, lane 5). Preparations of Vpr1–71
typically displayed good solubility at concentrations of up to
⬃15 mg/ml in detergent-free buffers of low ionic strength (e.g., 20 mM HEPES-NaOH [pH 7.3]). HIV-1 p6 was expressed as a glutathioneS-transferase (GST) fusion protein and was iso-lated by glutathione affinity chromatography, released from the GST moiety by thrombin cleavage, and purified to near homo-geneity by cation-exchange chromatography on S Sepharose (Fig. 1, lane 3).
In addition to its solubility and purity, Vpr1–71was judged to
be a useful reagent for studying the in vitro binding properties of Vpr based on the following functional criteria. One, the fusion of either Vpr1–71 or full-length Vpr to heterologous
proteins results in their nuclear accumulation (9). Consistent with this demonstration of nuclear import function, cytoplas-mic cytoplas-microinjection of our recombinant Vpr1–71 into HeLa
cells, followed by an indirect immunofluorescence assay,
re-sulted in localization to the nucleus (data not shown). Two, and most pertinent for the experiments discussed here, Vpr1–71,
like wild-type Vpr, can recruit appended proteins into progeny virions (Fig. 2). This was shown by cotransfection of 293T cells with thevpr-deficient HIV-1 provirus vector pYU-2/⌬vprand expression vectors for maltose-binding protein (MBP) (pCMV/MBP) or MBP-Vpr fusions (pCMV/MBP-Vpr and pCMV/MBP-Vpr1–71) (9). Whole-cell lysates and pelleted
viri-ons were then analyzed by Western blotting by using an MBP-specific primary antibody (New England Biolabs). Even though the full-length MBP-Vpr chimera was packaged at a higher level, both Vpr fusions were specifically incorporated into viri-ons whereas MBP alone was not (compare lanes 5 and 6 with lane 4). These results are consistent with previous reports that the C-terminal end of Vpr is not required for packaging but that truncation of Vpr may diminish protein stability (7).
Several groups have reported that different fragments of Vpr can self-associate (3, 14, 29, 31, 41). To test whether the Vpr1–71protein could also self-associate, we carried out both
gel filtration (Fig. 3) and covalent cross-linking (Fig. 4) anal-yses. Vpr1–71samples ranging in concentration from 8.6 to 500 M were applied to a Superdex 75 column (Pharmacia), and the column was developed in 130 ml of buffer (100 mM NaCl, 20 mM sodium phosphate, pH 7.2). A subset of these elution profiles is shown in Fig. 3A. In each case, Vpr1–71eluted in two
separate peaks, with the first always appearing at the void
[image:2.587.64.257.75.223.2]FIG. 2. Virion packaging of MBP-Vpr fusion proteins. 293T cells (107) were cotransfected with pYU-2/⌬vprand a vector encoding MBP, MBP-Vpr, or MBP-Vpr1–71, as indicated. At 24 h, culture supernatants were clarified by centrifugation and filtered through 0.45-m-pore-size membranes, and virions were pelleted through a 20% sucrose cushion by ultracentrifugation at 100,000⫻gfor 90 min. The transfected cells (lanes 1 to 3) and pelleted virions (lanes 4 to 6) were resuspended in loading buffer and resolved in parallel by SDS-PAGE. Gag and MBP-containing proteins were detected by Western blotting by using a p24Gag-specific monoclonal antibody or an MBP-specific rabbit anti-serum, respectively, and an enhanced chemiluminescence assay. FIG. 1. Purification of recombinant p6 and Vpr1–71. Shown are
SDS-PAGE and Coomassie blue staining of GST-p6 following gluta-thione-Sepharose affinity chromatography (lane 1), GST-p6 after cleavage with thrombin (lane 2), p6 after purification over S Sepharose (lane 3), Vpr1–71-His6following Ni1⫹chelate affinity chromatography (lane 4), and Vpr1–71after cleavage with enterokinase and purification over Q Sepharose (lane 5). In both cases, mass spectrometry confirmed that the final products had the expected molecular masses (data not shown). The values on the left are molecular masses in kilodaltons.
on November 9, 2019 by guest
http://jvi.asm.org/
volume (⬃47 ml). The fraction of Vpr1–71in this peak slowly
increased as the protein was stored in solution for extended periods, and upon collection and rechromatography, this frac-tion eluted exclusively with the void volume (data not shown). Based on their large Stokes radii and invariant behavior, we have assumed that these complexes represent large, inert ag-gregates of Vpr1–71.
In contrast, the retention time of the second peak increased with decreasing protein concentrations (Fig. 3A and B). The smaller size and concentration-dependent mobility of this peak suggested that it was composed of discrete Vpr1–71oligomers
that could rapidly and reversibly reequilibrate. The masses of Vpr1–71complexes in the second peak were estimated by
com-paring the concentration-dependent retention times to a stan-dard curve of retention times of globular proteins whose masses were known (Fig. 3C). These experiments indicated that the maximal and minimal hydrodynamic masses of Vpr1–71
were⬃11.7 kDa (at limiting low concentrations) and ⬃31.3 kDa (at limiting high concentrations). We assume that the smaller species represents monomeric Vpr1–71 (actual mass,
9.051 kDa), and our data are therefore most consistent with the formation of trimeric Vpr1–71 complexes at high protein
concentrations.
A limitation of these experiments is that gel filtration mea-surements reflect the Stokes radius (rather than true mass) of the protein or complex under study. We attempted to circum-vent this problem by using equilibrium sedimentation experi-ments to quantitate Vpr oligomerization. Although these data were again most consistent with the trimerization of Vpr1–71,
we were unable to obtain satisfactory global fits of our data over a large range of protein concentrations, presumably owing to difficulties with fitting of the fraction of Vpr1–71that
aggre-gated irreversibly.
To evaluate the oligomerization of Vpr1–71 by covalent
cross-linking, 50 M Vpr1–71 was incubated with increasing
concentrations of the bifunctional agent ethylene glycol bis-(succinimidylsuccinate) (EGS; Pierce) for 60 min at 37°C. Cross-linked adducts were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and visual-ized following staining with Coomassie blue (Fig. 4). As above, unmodified Vpr1–71migrated as a single species with an
ap-parent relative molecular mass of⬃9 kDa (lane 1). At all of the concentrations of EGS added, two additional bands of Vpr1–71were observed (lanes 2 to 7). Based on their relative
electrophoretic mobililties, these species appear to correspond to dimers and trimers. Although very small quantities of high-FIG. 3. Size exclusion chromatography of Vpr1–71. (A) Elution
[image:3.587.312.530.69.223.2]pro-files of pure Vpr1–71on Superdex 75 at the indicated initial concen-trations. (B) Concentration dependence of the elution volume for the aggregate (䡺) and oligomeric (f) fractions. (C) Standard curve show-ing elution volumes of protein markers (E) and Vpr1–71(F) at the indicated concentrations. The molecular mass (MW) standards are as follows: bovine serum albumin, 68 kDa; ovalbumin, 45 kDa; chymo-trypsinogen A, 25 kDa; cytochromec, 12.5 kDa.
FIG. 4. Cross-linking of Vpr1–71. Vpr1–71at 50M was incubated with the indicated molar ratios of EGS for 60 min at 37°C and visu-alized by SDS-PAGE and Coomassie staining. Cross-linking interme-diates representing monomer, dimer, and trimer forms of Vpr1–71are indicated. The values on the left are molecular masses in kilodaltons.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.587.48.284.71.554.2]er-order oligomers were sometimes observed at high cross-linker concentrations (⬃5 mM), these species never formed efficiently. Overall, these data again support the conclusion that Vpr1–71can trimerize in solution.
Having defined some of the properties of the Vpr1–71,
re-agent, we next turned our attention to the analysis of Vpr-p6 interactions. As an initial approach to the examination of di-rect binding, standard pull-down experiments were conducted by using purified Vpr1–71and GST-p6; despite testing a variety
of assay conditions, we consistently failed to detect an inter-action between the two proteins (data not shown). Although the absence of observable binding might have reflected a lack of complex formation, we also considered the alternative pos-sibility that the p6-Vpr1–71complex dissociates rapidly and is
therefore undetectable under the nonequilibrium binding con-ditions required in this experimental configuration (i.e., one that includes extensive washing steps).
The Vpr-p6 interaction was therefore examined by using optical biosensor experiments that measure binding in real time and have the potential to detect transient complexes. All surface plasmon resonance measurements were performed with a BIACORE 2000 or 3000 instrument (Biacore AB, Upp-sala, Sweden) equipped with research grade CM5 sensor chips. Initially, the binding of pure, soluble, monomeric p6 to hexa-histidine-tagged Vpr proteins was examined. Vpr-His6proteins
were captured directly from crude bacterial expression lysates by using an immobilized anti-His4monoclonal antibody
(Qia-gen). Importantly, this strategy circumvented the aforemen-tioned difficulties associated with purifying aggregation-prone Vpr proteins and made it possible to study binding to both Vpr1–71 and full-length Vpr (Vpr1–97). Approximately 1,000
response units (RU) of Vpr1–97-His6was captured in flow cell
2, and approximately 600 RU of Vpr1–71-His6was captured in
flow cell 3. Flow cells 1 and 4 served as reference surfaces. Replicates of the analyte, p6, in 20 mM Tris–100 mM NaCl– 0.005% polyoxyethylene 20-sorbitan monolaurate (P20) (pH 8.0) were injected over the four flow cells at concentrations of 7.5, 15.5, 31.2, 62.5, 125, and 250 M at a flow rate of 50
l/min. The analyses were performed at 20°C, and no regen-eration of the flow cell surfaces was required; data were col-lected at a rate of 2 Hz.
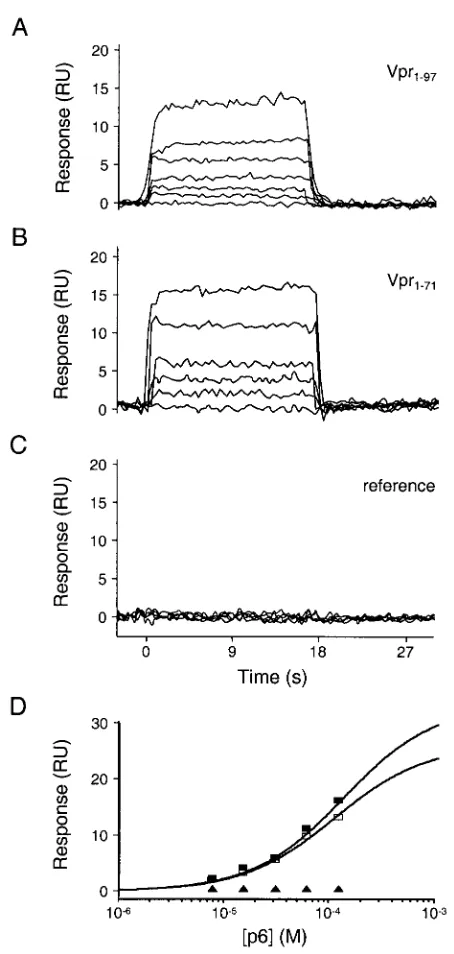
As illustrated in Fig. 5A to C, p6 bound each Vpr protein in a concentration- and Vpr-dependent fashion. Both Vpr-p6 complexes dissociated (and associated) very rapidly, with the half-lives of both complexes being less than 1 s. These rapid rates of dissociation are consistent with our inability to detect Vpr-p6 binding by using pull-down assays. Dissociation con-stants were estimated by fitting the equilibrium phases of the p6 binding isotherms by using simple 1:1 binding models (40), and both complexes bound with dissociation constants of⬃75
M (Table 1). To test the specificity of the interaction, we also examined the binding of p6 to a surface derivatized with Vpr1– 71-His6harboring a leucine-to-alanine substitution at residue
30 (A30L). This mutation, which inhibits Vpr packaging in vivo (7), prevented detectable p6 binding (Table 1) and thereby confirmed the validity of studying Vpr-p6 interactions by sur-face plasmon resonance.
Biosensor analysis was also used to quantitate the Vpr-p6 binding interaction in the opposite orientation. In this format, GST fusion proteins were captured by using an immobilized
GST-specific antibody as follows: flow cell 1, 1,200 RU of GST-p6 (wild type); flow cell 2, 1,200 RU of GST-p6R42A,S43A
(contains alanine substitutions at the second and third posi-tions of the LXXLF motif); flow cell 3, 700 RU of recombinant GST (reference sample); flow cell 4, 1,000 RU of GST-p6L44A
(contains an inactivating mutation in the LXXLF motif) (17). To collect equilibrium binding data, triplicate injections of Vpr1–71were allowed to flow over the four flow cells at
con-FIG. 5. Biosensor analysis of p6 binding to Vpr1–97and Vpr1–71. Representative sensorgrams of p6 injected at concentrations of 0, 7.5, 15.5, 31.2, 62.5, and 125M over Vpr1–97, (A) Vpr1–71(B), and a reference surface (C). (D) Responses at equilibrium (t⫽15 s) were plotted versus the injected p6 concentrations. Responses were fitted to simple 1:1 binding models to determine dissociation constants (Kds) of 71⫾8 and 84⫾6M for Vpr1–97and Vpr1–71, respectively. Symbols: f, Vpr1–97;䡺, Vpr1–71;Œ, reference surface.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.587.307.534.70.547.2]centrations of 0.7, 1.4, 2.9, 5.7, 11.5, 22.9, and 45.8M and a flow rate of 50l/min. Analogous binding studies were per-formed for two additional packaging-defective mutant forms of p6, L41A (⬃1,200 RU) and F45A (⬃1,300 RU) (17).
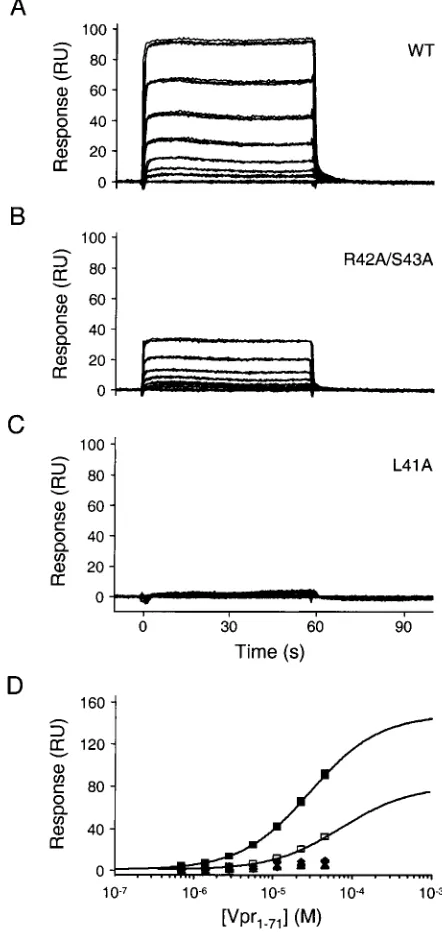
Consistent with our previous results, Vpr1–71bound to p6 in
a dose-dependent fashion, with complex dissociation again be-ing very rapid (Fig. 6A). Within a sbe-ingle experiment, the bind-ing measurements were highly reproducible (compare the overlaid triplicates in Fig. 6A), although absolute binding lev-els did vary somewhat between independent repetitions. The apparent dissociation constant derived from four such repeti-tions was 18⫾ 11M. Both the variability and apparently tighter Vpr1–71-p6 complexation observed in this orientation
likely reflect avidity effects arising from Vpr oligomerization and aggregation; as a result, the data do not provide a true measure of the intrinsic equilibrium dissociation constant of the complex.
Specificity of binding was evaluated by using GST-p6 fusions that carried alanine substitutions in the LXXLF motif. As shown in Fig. 6B and C (and Table 1), each of the mutations known to block the packaging of Vpr into virions failed to bind Vpr1–71at detectable levels. In contrast, noninterfering
substi-tutions of residues 42 and 43 (17) reduced the apparent bind-ing affinity only modestly (Kd⫽71M). Finally, we also tested the binding of Vpr1–71 to a seven-residue minimal peptide
spanning p6 residues 39 to 45 (immobilized as a GST fusion). Consistent with the ability of this motif to mediate efficient Vpr packaging when appended to a heterologous Gag protein (17), the apparent binding affinity was reduced only⬃1.7-fold rela-tive to that of full-length p6 (Table 1). In sum, our binding data demonstrate that pure recombinant p6 and Vpr proteins form a direct, specific complex in vitro. Moreover, the sequence requirements for binding correlate very well with the proven requirements for Vpr packaging in vivo. The most parsimoni-ous interpretation of these experiments is that the virion in-corporation of Vpr involves a direct interaction with the LXXLF motif of p6Gag.
It is noteworthy that both HIV-1 Gag and Vpr oligomerize and form higher-order structures in solution (3, 10, 14, 29, 31, 41). It is most likely that such interactions will significantly increase the avidities and decrease the dissociation rates of Vpr-Gag complexes that form during virion assembly. Thus,
although we cannot rule out the possibilities that Vpr might contact additional domains within p55Gagor that p6Gagmight
[image:5.587.310.531.80.546.2]adopt a different structure in the context of the Gag precursor, we believe that efficient Vpr packaging need not require any additional sites of contact beyond the LXXLF motif. Interest-ingly, analysis of virus cores purified from mature HIV-1 viri-ons has revealed that Vpr remains associated with the core but TABLE 1. Estimated dissociation constants for HIV-1
Vpr-p6 complexes
Protein-protein interaction Kd(M) Vpr1–97-His6–p6 ...71 (8),a66 (6) Vpr1–71-His6–p6 ...84 (4), 77 (3) Vpr1–71A30L-His6–p6 ...—b
GST-p6–Vpr1–71...18⫾11c GST-p6L41A–Vpr
1–71...— GST-p6R42A,S43A–Vpr
1–71...71 (3) GST-p6L44A–Vpr
1–71...— GST-p6F45A–Vpr
1–71...—
aThe values in parentheses represent estimated errors of 1 standard deviation
in the final digit and were derived from the fits of a single binding experiment. Data from multiple independent binding experiments are listed sequentially.
b—, binding was too weak to quantify (⬎200M).
cThis value is the average of four independent measurements made with
different protein preparations. The error is the standard deviation in the re-peated experiments.
FIG. 6. Biosensor analysis of Vpr1–71binding to p6. Representative sensorgrams of Vpr1–71 injected in triplicate at concentrations of 0, 0.72, 1.4, 2.9, 5.7, 11.5, 22.9, and 45.8M over wild-type (WT; A), R42A/S43A (B), and L41A (C) GST-p6. (D) Responses at equilibrium (t ⫽50 s) were plotted versus the injected Vpr1–71concentrations. Responses were fitted to simple 1:1 binding models for the wild-type and R42A/S43A p6 proteins. Binding of the other mutant p6 proteins was too weak to quantify. Symbols:f, wild-type GST-p6;䡺, R42A/ S43A;}, F45A;Œ, L44A;F, L41A.
on November 9, 2019 by guest
http://jvi.asm.org/
that the p6 protein is lost (1, 35). We suggest that this obser-vation is also in accordance with our avidity model; specifically, because the fully processed p6 protein is a monomer (34), its removal from the rest of Gag would lower the affinity of the Vpr-p6 complex and facilitate the rapid dissociation of p6 away from the core. Indeed, we speculate that the use of avidity to amplify weak but specific binding interactions may provide a general mechanism for packaging of viral components. By sub-sequently exploiting proteolysis to separate packaging signals from sites of oligomerization, it may then be possible to free virion components and enable them to execute important func-tions in newly infected cells.
Y.J. and O.P. contributed equally to this work. We thank Laurie Zimmerman for secretarial support.
This work was supported by NIH research grants AI46942 (M.H.M.), AI09996 (Y.J.), and AI45405 (W.I.S.).
REFERENCES
1.Accola, M. A., A. O¨ hagen, and H. G. Go¨ttlinger.2000. Isolation of human immunodeficiency virus type 1 cores: retention of Vpr in the absence of p6(gag). J. Virol.74:6198–6202.
2.Bachand, F., X. J. Yao, M. Hrimech, N. Rougeau, and E. A. Cohen.1999. Incorporation of Vpr into human immunodeficiency virus type 1 requires a direct interaction with the p6 domain of the p55 gag precursor. J. Biol. Chem.274:9083–9091.
3.Bogerd, H. P., R. A. Fridell, W. S. Blair, and B. R. Cullen.1993. Genetic evidence that the Tat proteins of human immunodeficiency virus types 1 and 2 can multimerize in the eukaryotic cell nucleus. J. Virol.67:5030–5034. 4.Bouyac-Bertoia, M., J. D. Dvorin, R. A. M. Fouchier, Y. Jenkins, B. E. Meyer,
L. I. Wu, M. Emerman, and M. H. Malim.2001. HIV-1 infection requires a functional integrase NLS. Mol. Cell7:1025–1035.
5.Cohen, E. A., E. F. Terwilliger, Y. Jalinoos, J. Proulx, J. G. Sodroski, and W. A. Haseltine.1990. Identification of HIV-1vprproduct and function. J. Acquir. Immune Defic. Syndr.3:11–18.
6.de Rocquigny, H., P. Petitjean, V. Tanchou, D. Decimo, L. Drouot, T. Delau-nay, J. L. Darlix, and B. P. Roques.1997. The zinc fingers of HIV nucleo-capsid protein NCp7 direct interactions with the viral regulatory protein Vpr. J. Biol. Chem.272:30753–30759.
7.Di Marzio, P., S. Choe, M. Ebright, R. Knoblauch, and N. R. Landau.1995. Mutational analysis of cell cycle arrest, nuclear localization, and virion pack-aging of human immunodeficiency virus type 1 Vpr. J. Virol.69:7909–7916. 8.Fletcher, T. M., III, M. A. Soares, S. McPhearson, H. Hui, M. Wiskerchen, M. A. Muesing, G. M. Shaw, A. D. Leavitt, J. D. Boeke, and B. H. Hahn. 1997. Complementation of integrase function in HIV-1 virions. EMBO J. 16:5123–5138.
9.Fouchier, R. A. M., B. E. Meyer, J. H. M. Simon, U. Fischer, A. V. Albright, F. Gonza´lez-Scarano, and M. H. Malim.1998. Interaction of the human immunodeficiency virus type 1 Vpr protein with the nuclear pore complex. J. Virol.72:6004–6013.
10.Freed, E. O.1998. HIV-1 gag proteins: diverse functions in the virus life cycle. Virology251:1–15.
11.Gallay, P., V. Stitt, C. Mundy, M. Oettinger, and D. Trono.1996. Role of the karyopherin pathway in human immunodeficiency virus type 1 nuclear im-port. J. Virol.70:1027–1032.
12.Gibbs, J. S., A. A. Lackner, S. M. Lang, M. A. Simon, P. K. Sehgal, M. D. Daniel, and R. C. Desrosiers.1995. Progression to AIDS in the absence of a gene forvprorvpx. J. Virol.69:2378–2383.
13.Heinzinger, N. K., M. I. Bukrinsky, S. A. Haggerty, A. M. Ragland, V. Kewalramani, M.-A. Lee, H. E. Gendelman, L. Ratner, M. Stevenson, and M. Emerman.1994. The Vpr protein of human immunodeficiency virus type 1 influences nuclear localization of viral nucleic acids in nondividing host cells. Proc. Natl. Acad. Sci. USA91:7311–7315.
14.Henklein, P., K. Bruns, M. P. Sherman, U. Tessmer, K. Licha, J. Kopp, C. M. de Noronha, W. C. Greene, V. Wray, and U. Schubert.2000. Func-tional and structural characterization of synthetic HIV-1 Vpr that transduces cells, localizes to the nucleus, and induces G2 cell cycle arrest. J. Biol. Chem. 275:32016–32026.
15.Hirsch, V. M., M. E. Sharkey, C. R. Brown, B. Brichacek, S. Goldstein, J. Wakefield, R. Byrum, W. R. Elkins, B. H. Hahn, J. D. Lifson, and M. Stevenson.1998. Vpx is required for dissemination and pathogenesis of SIVSM PBj: evidence of macrophage-dependent viral amplification. Nat.
Med.4:1401–1408.
16.Hrimech, M., X. J. Yao, F. Bachand, N. Rougeau, and E. A. Cohen.1999. Human immunodeficiency virus type 1 (HIV-1) Vpr functions as an imme-diate-early protein during HIV-1 infection. J. Virol.73:4101–4109.
17.Kondo, E., and H. G. Go¨ttlinger.1996. A conserved LXXLF sequence is the major determinant in p6gagrequired for the incorporation of human
immu-nodeficiency virus type 1 Vpr. J. Virol.70:159–164.
18.Kondo, E., F. Mammano, E. A. Cohen, and H. G. Go¨ttlinger.1995. The p6gag
domain of human immunodeficiency virus type 1 is sufficient for the incor-poration of Vpr into heterologous viral particles. J. Virol.69:2759–2764. 19.Kotov, A., J. Zhou, P. Flicker, and C. Aiken.1999. Association of Nef with
the human immunodeficiency virus type 1 core. J. Virol.73:8824–8830. 20.Li, M. S., G. Garcia-Asua, U. Bhattacharyya, P. Mascagni, B. M. Austen,
and M. M. Roberts.1996. The Vpr protein of human immunodeficiency virus type 1 binds to nucleocapsid protein p7 in vitro. Biochem. Biophys. Res. Commun.218:352–355.
21.Liu, H., X. Wu, M. Newman, G. M. Shaw, B. H. Hahn, and J. C. Kappes. 1995. The Vif protein of human and simian immunodeficiency viruses is packaged into virions and associates with viral core structures. J. Virol. 69:7630–7638.
22.Lu, Y.-L., R. P. Bennett, J. W. Wills, R. Gorelick, and L. Ratner.1995. A leucine triplet repeat sequence (LXX)4in p6gagis important for Vpr
incor-poration into human immunodeficiency virus type 1 particles. J. Virol.69: 6873–6879.
23.Lu, Y.-L., P. Spearman, and L. Ratner.1993. Human immunodeficiency virus type 1 viral protein R localization in infected cells and virions. J. Virol. 67:6542–6550.
24.Mansky, L. M., S. Preveral, L. Selig, R. Benarous, and S. Benichou.2000. The interaction of Vpr with uracil DNA glycosylase modulates the human immunodeficiency virus type 1 in vivo mutation rate. J. Virol.74:7039–7047. 25.Mu¨ller, B., U. Tessmer, U. Schubert, and H.-G. Kra¨usslich.2000. Human immunodeficiency virus type 1 Vpr protein is incorporated into the virion in significantly smaller amounts than Gag and is phosphorylated in infected cells. J. Virol.74:9727–9731.
26.Paxton, W., R. I. Connor, and N. R. Landau.1993. Incorporation of Vpr into human immunodeficiency virus type 1 virions: requirement for the p6 region ofgagand mutational analysis. J. Virol.67:7229–7237.
27.Poon, B., K. Grovit-Ferbas, S. A. Stewart, and I. S. Y. Chen.1998. Cell cycle arrest by Vpr in HIV-1 virions and insensitivity to antiretroviral agents. Science281:266–269.
28.Sato, A., J. Yoshimoto, Y. Isaka, S. Miki, A. Suyama, A. Adachi, M. Hayami, T. Fujiwara, and O. Yoshie.1996. Evidence for direct association of Vpr and matrix protein p17 within the HIV-1 virion. Virology220:208–212. 29.Sawaya, B. E., K. Khalili, J. Gordon, A. Srinivasan, M. Richardson, J.
Rappaport, and S. Amini.2000. Transdominant activity of human immuno-deficiency virus type 1 Vpr with a mutation at residue R73. J. Virol.74:4877– 4881.
30.Schaeffer, E., R. Geleziunas, and W. C. Greene.2001. Human immunodefi-ciency virus type 1 Nef functions at the level of virus entry by enhancing cytoplasmic delivery of virions. J. Virol.75:2993–3000.
31.Schuler, W., K. Wecker, H. de Rocquigny, Y. Baudat, J. Sire, and B. P. Roques.1999. NMR structure of the (52–96) C-terminal domain of the HIV-1 regulatory protein Vpr: molecular insights into its biological func-tions. J. Mol. Biol.285:2105–2117.
32.Selig, L., J. C. Pages, V. Tanchou, S. Pre´ve´ral, C. Berlioz-Torrent, L. X. Liu, L. Erdtmann, J. L. Darlix, R. Benarous, and S. Benichou.1999. Interaction with the p6 domain of the Gag precursor mediates incorporation into virions of Vpr and Vpx proteins from primate lentiviruses. J. Virol.73:592–600. 33.Stauber, R. H., S. Rulong, G. Palm, and N. I. Tarasova.1999. Direct
visu-alization of HIV-1 entry: mechanisms and role of cell surface receptors. Biochem. Biophys. Res. Commun.258:695–702.
34.Stys, D., I. Blaha, and P. Strop.1993. Structural and functional studies in vitro on the p6 protein from the HIV-1 gag open reading frame. Biochim. Biophys. Acta1182:157–161.
35.Welker, R., H. Hohenberg, U. Tessmer, C. Huckhagel, and H.-G. Kra¨usslich. 2000. Biochemical and structural analysis of isolated mature cores of human immunodeficiency virus type 1. J. Virol.74:1168–1177.
36.Wu, X., H. Liu, H. Xiao, J. A. Conway, E. Hehl, G. V. Kalpana, V. Prasad, and J. C. Kappes.1999. Human immunodeficiency virus type 1 integrase protein promotes reverse transcription through specific interactions with the nucleoprotein reverse transcription complex. J. Virol.73:2126–2135. 37.Wu, X., H. Liu, H. Xiao, J. A. Conway, E. Hunter, and J. C. Kappes.1997.
Functional RT and IN incorporated into HIV-1 particles independently of the Gag/Pol precursor protein. EMBO J.16:5113–5122.
38.Wu, X., H. Liu, H. Xiao, J. A. Conway, and J. C. Kappes.1996. Inhibition of human and simian immunodeficiency virus protease function by targeting Vpx-protease-mutant fusion protein into viral particles. J. Virol.70:3378–3384. 39.Wu, X., H. Liu, H. Xiao, J. Kim, P. Seshaiah, G. Natsoulis, J. D. Boeke, B. H. Hahn, and J. C. Kappes.1995. Targeting foreign proteins to human immu-nodeficiency virus particles via fusion with Vpr and Vpx. J. Virol.69:3389–3398. 40.Yoo, S., D. G. Myszka, C. Yeh, M. McMurray, C. P. Hill, and W. I. Sundquist.1997. Molecular recognition in the HIV-1 capsid/cyclophilin A complex. J. Mol. Biol.269:780–795.
41.Zhao, L. J., L. Wang, S. Mukherjee, and O. Narayan.1994. Biochemical mechanism of HIV-1 Vpr function. Oligomerization mediated by the N-terminal domain. J. Biol. Chem.269:32131–32137.