Copyright © 1967 AmericanSociety forMicrobiology Printed inU.S.A.
Separation
and
Characterization
of
Soluble
Adenovirus Type 9
Components
E. NORRBY, B. NYBERG, P. SKAARET, AmN A. LENGYEL1
DepartmentofVirology,School of Medicine, KarolinskaInstitutet,Stockholm, Sweden
Receivedforpublication2August 1967
Fourdifferentsoluble components ofadenovirus type 9 (Rosen's group II) were
identified. These were a complete hemagglutinin (HA), an incomplete HA,
com-ponentscarrying group-specific comple'ment-fixing (CF) antigen, and components
identified only by their hemagglutination-inhibition (HI) antibody consuming
capacity andantigenactivity inCFtestswithan antiserum againstcomplete HA.
Thecomplete HA sedimented relatively rapidly. It was composed of 12 pentons
(vertexcapsomersplus
projections)
aggregatedintotheformof apentagonaldodec-ahedron. Thelength
of
the projectionswas about 12 to 13 m,. Thus they appearedlonger thanthecorresponding structures oftypes 3 and 11, but shorter than those
oftypes 4 and 5. The rate of sedimentation of completeHA oftype 9 was
inter-mediatetothose
of
thecomplete HA oftypes 3 and 11. TheincompleteHAsedi-mentedtogetherwith componentscarrying group-specificCFantigen,butcould be
separated from those by
anion-exchange
chromatography. Two different antigenswere present in
incomplete
HA. One couldabsorb a group-specifichemagglutina-tion-enhancing antibody, and was sensitive to treatment with trypsin. The other
antigen could absorbthetype-specificHIantibody andwas notdestroyedby trypsin.
Inadditiontothe
incomplete
HA,aseparatepopulationofmoreslowly sedimentingcomponents showed a capacity to absorb HI antibody. These components could
also beidentified inCF tests when an antiserum against completeHAwas applied.
Theincomplete HA,group-specificCFantigen,andslowly sedimentingHIantibody
absorbing componentsaresuggestedtorepresent
isolated
penton, hexon, andfibercomponents,
respectively.
Adenoviruses are
separated
into three sub.groups on the basis of their
hemagglutinating
activity
(14).
Studies on soluble components have beenmainly
concerned with members ofsubgroup III,
e.g., types 2 and 5(8,
13,
17, 20),
and, rrorerecently,
with members ofsubgroup
I, e.g., types 3 and 11
(9-11).
Soluble nonvertexcapsorr
ers, vertex capsomersplus projections,
and isolatedvertex
projections
weredistinguished
in
preparations
of alltheseserotypes. Thesecom-ponents have been named
hexon,
penton, andfiber
antigens
(5),
and thisterminology
will
beused in the
following
presentation. However,
forreasons
given
ina recentreview(Current Topics
ofMicrobiology
andImmunology,
inpress),
the term"antigen"
will bereplaced
by
"component."
The
complete
hemagglutinin
(HA)
ofmembersof
subgroup
Iwasidentifiedas onemorekind ofsoluble component, and was found to be
com-IOn leave from the Institute of
Microbiology,
University MedicalSchool,Budapest, Hungary,on a
World HealthOrganizationresearch
fellowship.
posed of12 pentonsaggregated intheform ofa
pentagonal dodecahedron. No similar structure
has been identified in preparations of members
of subgroup III, with the exception of type 4
(12, 18). Members ofsubgroups I and m also
appear to differ with regardto the
immunologi-cal complexity of their fiber components (11,
17).
Against the background of this diversity in
structural and functional characteristics of
solu-ble components of members of
subgroups
IandIII, it was considered of interest also to study
the
corresponding
components ofa member ofsubgroup II. Adenovirus type 9 was chosen as
an object for this study. Available information
oncharacteristicsofmembers of
subgroup
II ismeager, andmainly derivesfrom studies carried
out by
Wigand's
group(1, 2,
19). They
havedemonstratedthat some of the
hemagglutinating
activity
of "whole" virus material is associatedwith
virions,
but that themajor
part of it is carried by a soluble HA. This solublecompo-1101
on November 11, 2019 by guest
http://jvi.asm.org/
nent is distinct from the soluble group-specific complerrent-fixing (CF) antigen, as
demron-strated by anion-exchange chromatography (4) and columnelectrophoresis experiments (15). In
the following, results are presented ofattempts to identify additional soluble components and to analyze the relationship between different virusproducts with regardtotheir structure and function.
MATERIALS AND METHODS
Virus and cell cultures. The prototype strain
("Hicks") of adenovirus type 9 was used. Before
arrival in this laboratory, it had been passed3 times onKBand 10 timesonHeLacells.Materialtobeused in thepresent experiments was preparedinahuman bonemarrowcellline,denoted Masa cells (7),orina
human embryonic lung cell line, Lu 106 cells. The
techniques for handling these cells were previously
described (9, 10).
Medium and cells harvested atanadvanced stage
of virus-induceddegenerationwereconcentrated 10to
20times by forced dialysisagainstpolyethylene glycol
(Carbowax 6000, Union Carbide Chemicals Co.,
New York, N.Y.). Theconcentrate was frozen and
thawedthreetofivetimes,afterwhichcelldebriswas
eliminated. Threeconsecutiveultracentrifugations for
1 hr at 20,000 rev/min (rotor 40, Spinco Division,
BeckmanInstruments Inc., LosAngeles, Calif.) were
thenperformedtoremovevirionsand emptycapsids
from the material. Infectivitytiterswerereducedbya factor of105or morebythis treatment.
Preparation ofimmunesera.Antiserawereprepared
byhyperimmunizationof rabbits withconcentratesof
different prototype adenovirus strains. Amounts of 4 mlof material mixed with Freund'scomplete
adju-vant were given intramuscularly as a primary
immunization. Thiswas followed 4 to 5 weeks later
byanintravenous booster of 1 to2ml. Theanimals
were thenexsanguinated after another week.
Determination of complete and incomplete HA. Serialtwofold dilutions (0.4 ml) of material in
phos-phate-buffered0.067 M,(pH 7.2) physiological saline
(PBS), or the same buffer containing heterotypic
adenovirusantiserum,wereusedfordetermination of
complete and incomplete HA, respectively. The
hemagglutination-enhancement (HE) antibody titer of serawas determined in chessboardtitrations against a reference preparation of incomplete HA prepared
by erythrocyte absorption of"whole" virus material
(10, andseebelow).TwotofourHEserumunitswere
employedper dilution intests for incomplete
adeno-virustype9 HA, andthe testswere incubated for 1
houratroomtemperaturebefore addition of
erythro-cytes. Either fresh rat or human 0 erythrocytes
(0.2 ml)wereaddedintheform ofa0.5%suspension.
Readings were taken by bottom patterns. The final
tubeexhibitingacompleteorclearcut partial
aggluti-nation was considered to contain one unit of
HA (HAU).
Hemagglutination-enhancement and hemagglutina-tion-inhibition antibody consumption (HEC andHIC) tests.Thetheoreticalbackgroundtothesetests,which
were used foridentification ofvertexcapsomers and
projections, respectively, present either inanisolated
orcombined form, wasalready described (11). Both testscomprisedthemixing of serial twofold dilutions
(0.2ml) ofmaterial with 0.1 ml ofadilution of
homo-typic antiserum. After incubation ofthese mixtures
for 1 hr at roomtemperature,0.1 ml ofanindicator
antigen was added. This antigen was representedby
incomplete adenovirus type I I HA (10; J. Gen.
Virol., in press) and "whole" virus materialof type9
in the HEC and HIC tests, respectively. The
homo-typicantiserumwasdilutedtocontaintwo hemagglu-tination-inhibition (HI) or HE units per 0.1 ml,
referringtoeach oneof thesetwoantigens.Thehighest dilutionofmaterial whichcould absorb all HE
(ab-senceofhemagglutination) orHIantibodies (presence
ofhemagglutination) was consideredto containone
HEC or HIC test unit (HECU and HICU),
respec-tively.
Determinationof CF antigen activity.Themodified
drop technique described by Svedmyretal. (16) was
used. Twounitsofserumandtwounitsofcomplement
wereapplied perantigen dilution.Group-specificCF
antigen was determined with antivirion sera against
type 5. Antigens, capable offormingapart of
com-plete HA, were assayed by use of antisera obtained
after immunization withpurifiedpreparations of this
component(seebelow).Noneof theserausedreacted
withnonviral components.
Zonal centrifugation. Thetechniqueforpreparation,
handling, and harvesting of linear 5 to 20%sucrose
gradients was already described (9).
Anion-exchange chromatography. Columns
meas-uring 1.5 X 40cm were packed with diethylamino-ethyl (DEAE)-Sephadex A25 (Pharmacia Fine Chemicals, Uppsala, Sweden). Prior to packing, the
gel was allowed to swell in an 0.04 M
tris(hydroxy-methyl)aminomethane (Tris) chloride buffer of pH
8.4 and washed carefully in the same buffer. The
temperature of fractionation was about 17 C. The
flow rate was maintained at approximately 5 to 10 ml cm-2 hr-1. Linear NaCl gradients were used for
elution of material retained by the column. A 4- to 8-mlamount ofmaterial, dialyzedagainstthe
above-mentioned Tris chloride buffer, was fractionated in each experiment. Equal fractions (volume of 3.0 to
3.5 ml) werecollected by use of an automatic drop
counter.
Electron microscopy. The negative-contrast
tech-niquewas used. Onedrop ofa 2%sodium
tungsto-silicate (STS) solution wasmixed on dentalwaxwith
onedropof thematerialtobeexamined.Priortothis,
the latterhad beendialyzed againsta1%ammonium
acetatesolution.Bovine albumin inafinal
concentra-tion of 0.01% was added if needed to improve the
spreadingofmaterialdroppedontothecarbon-coated
grids. Examinations weremade by use ofaJEM-SY
electron microscope at primary magnifications of
40,000to50,000.
Treatment with trypsin. Preparations to be treated
were dialyzed against a solution containing 0.15 M
NaCl,0.05 M Trischloride buffer (pH 7.2) and 0.001 M
CaCl2. Stock solutions oftrypsin (twicecrystallized,
Fluka AG, Buchs SG, Switzerland) and soybean
1102
on November 11, 2019 by guest
http://jvi.asm.org/
SOLUBLE
5 10 15 20
Fraction number
FiG. 1. Separation ofsoluble adenovirus type 9
com-ponents by zonalcentrifugationi in a linear 5 to 20%
sucrose gradient at 25,000 rev/min (SW25, Spinco)
for 5.5 hr. Fractions were collected dropwise through
the bottom of the tube (left). The following activities
were recorded: O, complete HA; *, incomplete HA
determined inthe presence of an antiadenovirus type 15
serum; 0, group-specific CF antigen assayed in tests
includinganantiadenovirus type 5 serum.
trypsin inhibitor (five times crystallized, Nutritional
BiochemicalsCorp., Cleveland, Ohio) were prepared
in the samebuffer.
RESULTS
Zonalcentrifugation ofa mixture of all soluble components. As in previous experiments (9, 10, 12), the zonal centrifugation technique was used
as a means to trace the possible occurrence of a
rapidly
sedimenting complete HA. The results ofone such experiment are presented in Fig. 1.
Under the conditions of centrifugation, the
complete HAwas recovered close to the bottom
of the tube. In the low-density region of the
gradient, group-specific
CF and an incompleteHA were recovered. Tests with rat and human 0 erythrocytes gave a similar distribution of both
complete and incomplete HA. The incomplete
HA was demonstrable in the presence of an
antiserum against type 15, another
mrember
ofRosen's subgroup II, but also in the presence of other heterotypic antisera, e.g., against types 3
(subgroup I) and5 (subgroupIII).
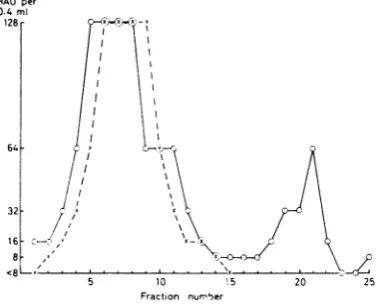
Comparison of the rate of sedimentation of
complete adenovirus types3,9, and11 HA. Since
previous findingshaverevealed thatthecomplete
HA of adenovirus type 11 sediments markedly
morerapidlythan the corresponding component
ofadenovirus type 3 (Norrby, J. Gen. Virol., in
press), it was considered of interest to relate the
sedimentation characteristics of type 9 complete
HA to that of the other two complete HA. It
was found (Fig. 2) that the completetype 9 HA
sedirrented irore slowly than the complete type
11 HA, but still significantly more rapidly than
thecorrespondingtype3 HA.
Ultrastruclural characteristics of complete adenovirus type 9 HA.Complete HA, purified by zonal centrifugation as shown in Fig. 1, was
used for electron
nf
icroscopy. Asmight
havebeen anticipated, this HAwas found tohave, in
principle, the
sanme
composition as the complete HA oftypes 3 (9) and 4(18),
i.e.,
asymmetrical
aggregate of 12
capsorrerlike
structures carryingprojections, each located at the facets ofa
pen-tagonal dodecahedron
(Fig. 3).
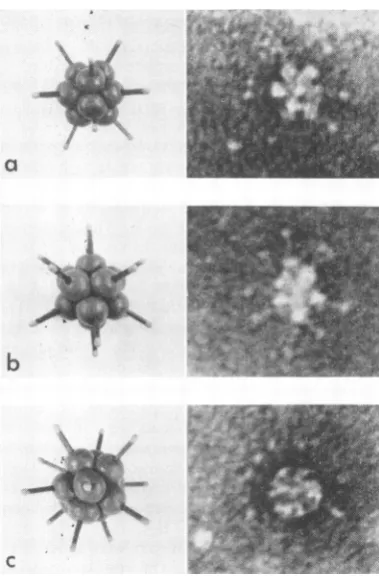
Thecapsorrer-like structures hadan outer diameter of 70 to60
A,and they formeda coreofcompleteHAwith
a diameter
varying
between 240 and 290 Ade-pendingupontheorientation ofthe
particle.
Theprojectionsextendingfrom thecorehada length
of 110 to 140 A, thus
giving
the "star"-shapedHAanoverall diarreter of500to560 A.
Interaction between different soluble
compo-nents and erythrocytes. Preparations of soluble
adenovirus type 9componentswere
subjected
tofourconsecutive
absorptions
withrat or, in mostexperirrents,
human 0 erythrocytes. The lattertype ofcell was
preferred
becausethey
were lessprone to undergo spontaneous
lysis.
Thecon-centrations of
complete HA,
incomplete
HA,and
group-specific
CFantigen
were determinedin the different samples. The results from one
experiment are shown in Table 1. The complete
HA was rapidly eliminated by the treatment,
HAU per
0.4ml 3
[image:3.471.35.223.52.221.2]Fraction number
FIG. 2. Zonalcentrifugationofa mixtureof
adeno-virus type3, 9, and 11 materialsin a linear5 to
20%lo
sucrosegradient (left = bottom oftube) for5.5 hrat
25,000 rev/min (SW 25, Spinco). Symbols: 0, HA
activitydetermined withmonkey erythrocytesat37 C.
After incubation of thetests at4 C,onl.y theleftpeak
of activity remained, revealing that this represents
complete adenovirus type 11 HA; *, HA activity
determinedwith rat erythrocytes.
VOL. 1, 1967
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.471.242.428.403.571.2]a
b
C'.:f ...
FIG. 3. Ultrastructuralappearanceofparticles
pres-ent in apreparation of complete HA purifiedby rate
zonalcentrifugation (see Fig. 1). The threeparticles
have been interpretedasbeing viewedalonga (a)
two-fold, (b) threetwo-fold,and (c) fivefoldaxis ofsymmetry.
X400,000.
whereas the incomplete HA was not so well
absorbed. Thetiter ofincomplete HA, asshown
in Table 1, wasnot significantly reduced.
How-ever, the outcome ofthe different experiments
waspartly dependent upon the titers of the
ma-terialpriortoabsorption. Ifthe ratioof
erythro-cyte concentration to quantities of biological activity washigher than in the experiment
illus-trated in Table 1,acomplete elimination of both
complete and incomplete HA could be obtained. In contradistinction, the titer of group-specific CF antigen always remained unchanged by the absorptions.
It has been reported that receptor-destroying
enzyme (RDE) caneliminate the agglutinability
of human 0 erythrocytes by some members of
Rosen's subgroup II (6). We therefore investi-gated the possibility that treatment with RDE could be used as arreans ofbreaking the
asso-ciation between complete HA and erythrocytes, which normally is irreversible. Packed
erythro-cytes from the first cycle ofabsorption ofvirus
material were washed three times in PBS to remove components which had not attached to the cells. Theerythrocyteswerethenresuspended in a standard reagent cholera filtrate product (N.V. Philips-Roxane, The Netherlands) diluted
1:8inPBSandincubated for 1 hr at room
tem-perature. The red cells were then removed by
low-speed centrifugation. As can be seen from
Table 1, this treatment of the cells with RDE ledto an elution of allcomplete HA. The eluate
did not contain any group-specific CF antigen.
In a similar way, an elution of complete HA
from agglutinated rat erythrocytes could be
ob-tained by treatment with RDE. However, in
connection with elution ofHA, therewas a
con-siderable lysis of these latter cells, because of
theirgeneralfragility.
Zonal centrifugation was used as a means to
characterize the
product
eluted from red cells.Figure 4 illustrates the quantitative distribution
in sucrosegradients of complete andincomplete
HA in unabsorbed material and an eluate from redcells.Whereas theformer materialcontained
both complete and
incomplete
HA, only the [image:4.471.52.242.72.361.2]former HA was demonstrable intheeluate.
TABLE 1. Effect of four consecutive erythrocyte
absorptions ondifferentbiological
activities ofapreparation of
solubleadenovirus type 9
components
CompleteHA Incom- Group-No.ofabsorptionsa (units/0ml pleteHA specitic
M) (unfits! CFunits/
m) 0.4
ml)b
0.62mlc0 12,800 256
1 128 1,024 256
2 <4 1,024 256
3 <4 1,024 256
4 <4 512 256
Eluate from cells of 12,800 -'d <4
absorptionI
aEachindividualabsorption wasperformed by
addition ofpackederythrocytesto afinal
concen-tration of10%.Afterincubationfor1 hr at room
temperature, thecellswereremovedby low-speed
centrifugation.
bDetermined in the presence ofan
antiadeno-virus type 15serum.
cAssayed in CF tests by use ofan
antiadeno-virustype5serum.
dNot measurable due to the presence ofhigh
concentrations ofcompleteHA.
eWashed and packed erythrocytes were
resus-pendedin standard reagent cholerafiltrate,diluted
1:8in PBS, to theoriginalvolume of the absorbed
material.
1104 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.471.254.451.348.519.2]TABLE 2. Effect of trypsin treatmentofdifferent soluble components ofadenovirus type 9
Treatment Complete HA Incomplete HECU HICU Grou Typeofprepna withb units per HAunits 0p4ler 0IC mer specificSFU
trypsinb 0.4ml per 0.4mlc . m . m per 0.02mId
Complete HA _ 3,200 NTe NT NT <2
+ <8 <8 NT NT NT
Incomplete HAplus group-specific _ <4 256 8 2 128
CFantigen + NT 16 <2 2 128
SlowlysedimentingHIC test <<4 <4 <2 4 <2
positivematerial + NT NT NT 4 NT
aComponentspreparedbyzonal
centrifugation
(Fig.
1 and5)
wereused.bIncubation withtrypsin inafinalconcentration of
0.2%
for 3 hrat37 C inawater bath. Controls-receivedbuffersolutionwithout trypsin.
cAssayedin the presenceofanantiadenovirustype15serum.
dDetermined byuseofanantiadenovirustype 5serum.
eNot tested.
HAUper
0.4ml
128r
5 10 5
Fraction nurnmer
FIG. 4. Distribution of hemagglutinat tested in thepresenceofanantiadenovirustj
ofpreparations ofsolubleadenovirustype9
priortoabsorptionwith (0) andafterelut
erythrocytes (X). Separation of comp
achievedbyzonalcentrifugationundersimile
asinFig.1.
Zonal centrifugation of soluble
excluding complete HA. Materials fr thecomplete HA had been removed cyte absorption (see Table 1) were st
zonal centrifugations at 23,000 rev/m
rotorSW 25) for 45 hr. Under these
of fractionation, incomplete HA a
specificCFantigenwererecoveredas
peak of activities in the high-density
the gradient (Fig. 5). HECtest, andso
HIC test,positivematerial and CFan
onstrated withananti-HA serum alsc
similar distribution. However, a co
fraction of the latter two activities v
by more slowly sedimenting compon
tions 12 to 14, Fig. 5). No clearcut
ltination was seen in the absence of I
antiserum in the same region of the gradient. However, in the presence of such a serum, the
bottom patterns at low dilutions offractions 12
to14werenotidentical withthat of the
erythro-cyte control containing PBS and heterotypic
antiserum. There was someform ofaggregation
of cells, whichwas most clearly revealed by the
relatively more sluggish slipping of sedimented
cells in these
fractions,
ascompared
with othernegative fractions, whenthe rack was held in an
inclinedposition.
Further separation of soluble components by anion-exchange chromatography. Fractionation
on the anion-exchanger DEAE-Sephadex A25
20 25 was tested as an additional
technique
forsepa-rationof soluble type 9components. Linear0.05
ring activity to 0.3M NaCl
gradients
were found togive
aype15serum good resolution of components in the elution
components diagram. As can be seen from Fig. 6, the se-Fion from rat quence of elution was incomplete HA,
group-7onents was specific CF antigen, and complete HA.
Appar-arconditions ently the former two activities were carried by
different solublecomponents.
Effect of trypsin treatment on the activity of
components different soluble components. Preparations of
rom which complete HA,
incomplete
HAmixed withgroup-by
erythro-
specificCFantigen, andslowlysedimentingHICabjected
to test positive material, which had been separatedin
(Spinco
by zonal centrifugations as described in Fig. 1conditions and 5, were used in these
experiments.
Thend group- preparations weretreated with trypsin in afinal
acommon concentration of
0.2%
for 3 hr in a water bathregion
of
(37 C). The activity ofthe enzyme was stopped omeof the by addition ofequal amounts of soybeantryp-sthgen
dem-
sin inhibitor. The results of oneexperiment
are) showed a illustrated in Table 2.
)nsiderable This treatment destroyed all complete HA. It
vas carried also eliminated
all
HEC testpositive
activity,
ents
(frac-
but did notreduce the HICtestpositive activity
herragglu- detectable in mixtures of incomplete HA and
heterotypic group-specific CFantigen. Similarly, there was a
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.471.38.227.256.408.2]NORRBY ET AL.
CFU 0.02r
64
32
16 8 4
per ml
I-0- 0 -*-
\
0_-3 6 9 2
Fractiori
numbe-FIG. 5. Distribuition of biological activities of a
preparation ofsoluble adenovirus type 9 components,
from which the complete HA had been removed by
erythrocyte absorptions, after zonal centrifugation in a
linear 5 to 20% sucrose gradient at 23,000 rev/min
(SW25, Spinco) for 45 hr. Fractions were collected
through the bottom of the tube (left). The following
activities were recorded: (a) incomplete HA
deter-mined in the presence ofan antiadenovirus type 15
serum; (AL)HECand(A) HIC test positive material;
(0)
group-specific and (a) HA-specific CFantigensdetermined by titrations with antisera against
adeno-virus type 5 and complete HAof type 9, respectively.
HAUper CFU -r
0.4n albuminpeak C02.
2048r 0 1 p<5 132
I
I
8I
q 'o-o-o
35 45 55 65
Fraction number
FIG. 6. Recovery ofcomplete HA (0), incomplete HA (U), andgroup-specificCFantigendetermined with antiadenovirus type5serum (0) after fractionationon
the anion exchanger DEAE-Sephadex A25. Different components were eluted by introduction ofa linear
0.05 to 0.3 M NaCI gradient in 0.04 M Tris chloride
(pH8.4).
marked reduction of incomplete HA
activity
ofthe latter. However, the titer of
group-specific
CF antigen rerrained unchanged by the
treat-mrent. Slowly sedimenting HIC test positive
mraterial, finally, also was unaffected by
incuba-tion withtrypsin.
DISCUSSION
The complete HA of adenovirus type 9 was
found toappear in the form of aggregates of 12
capsomer-like structures each carrying a
projec-tion. Structures with
principally
the samesym-metrical organization were already identified in
preparations of purified comrplete HA of
adeno-virus type 3 (9), 4 (18), 11 (Norrby, J. Gen.
Virol., in press) 13, 15, and 19 (Gelderblom et
al.,J.Gen.Virol.,inpress). Ithas beensuggested
that a completeHA of this type is formfed by 12
pentons, i.e., vertex capsomers plus projections,
aggregated aroundan internalcomponent
(9).
Different complete HAexhibit some variation
in structural details. As an example, it can be
mentioned that the
complete
HA of adenovirustype 11 sedimented rruch faster than the
corre-sponding component of type 3. This was
inter-pretedasbeing dueto adifference in mass of the
postulated internal component of thesetwoHA
(Norrby, J. Gen. Virol., inpress). In the present
study,thesedimentationrateof thecompletetype
9 HAwasfoundtobeintermediate betweenthose
ofthe types 3and 11 completeHA.The
explana-tion for this behaviorintermsof structural
differ-ences mustremain
partly
amatterofspeculation.The relatively longer
projection
of type 9 ascompared withtypes 3 and 11
might
bea factorof
imrportance.
However, an increase in relativelength of
projections
can influence thesedimen-tationrateintwodifferent ways. Itcancause an
increase in mass, but at the same time it can
increase the frictional ratio of comnponents. It
can be mentioned that the complete adenovirus
type 4HAsedimented with about thesame rate
astype 3 HA,
although
theirprojections,
which arecarried bycoresof similardimensions,
differin length, about 17 and 10 mg, respectively (9,
18). Thus, it appears unlikely that the variation betweensedimentation ratesof the complete HA of types 3 and 9 could be explained bya differ-encein this respect only. The dimensions of the
complete HA of type 9, excluding projections,
are similar to, or possibly slightly larger than,
those ofthe complete HA of type 3. The srrall
difference, iffurther confirmed, could be caused
by structural dissimilarities in either the capso-merlike orthesuspectedinternal component.
The complete HA of type 9 was effectively elirr inated byerythrocyte absorptions.The
inter-1106
HECUor HICU per
0.4ml
8
4 2
1 <1
J. VIROL.
32 16
8 <8
512
256 1OF
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.471.51.242.55.326.2] [image:6.471.51.242.457.578.2]action between HA and cells appeared to be
irreversible, but an elution of HA could be
ob-tained by treatment with RDE. Treatmrent with
K104 was less effective in bringing about an
elution of complete HA (Norrby and Lengyel, unpublished data). The relationship between
adenovirus type 9 and myxovirus receptors on
red cells is a subject to be studied further. The
effect of treatment of cells with Formalin
indi-cateda possible difference in nature ofreceptors
fortheseviruses(3).
Onthe conditionthat theratios ofthevarious
biological activities were favorable and proper
quantities of erythrocytes were used in
absorp-tion experiments, a product containing
incom-plete HA, but no complete HA, could be
ob-tained. With relatively
higher
concentrations oferythrocytes, both types of components were
eliminated. This behavior is similar to that of
type 11 components (Norrby, J. Gen. Virol., in
press), but different from that of type 3
compo-nents. T'he incomplete type 3 HA could not,
under the different conditons tested, be
elim-inated by absorption with red cells alone (10).
The incomplete type 9 HA sedimented together
with group-specific CF antigenin sucrose
gradi-ents, but could be
separated
from the lattercomponent
by
erythrocyteabsorptions,
as wasdiscussed
above,
anion-exchangechromatogra-phy and also gel filtration on Sephadex G200
(Norrby, unpublished data). It should be
men-tioned that the sequence of elution of various
adenovirus type 9 components from anion
ex-changers is different from that of the correspond-ing components of other serotypes studied
(8,
10). Therelative
positions
forgroup-specific
CFantigen and
complete
HA described abovecor-respondtothose
previously
describedby
Gelder-blometal.
(4)
for different members of Rosen'ssubgroupII.
The
incomplete
HA appears to becomposed
of two different parts. One part was
group-specific,
could absorb HEantibody,
and wastrypsin-sensitive.
It should be noted that theimmunological
specificity
of thisgroup-specific
antigen
is different from that ofgroup-specific
CFantigen,
determinedby
an adenovirus type5 antivirion serum
(see below).
The other partof the
incomplete
HA wastype-specific,
could absorb HIantibody
and was not sensitive totreatment with
trypsin.
Acomparison
withexperiences
gained
from studies of type 3(11)
indicates that the
incomplete
type 9 HArepre-sents pentons.
Accordingly,
thetwo parts of this component mostlikely
represent vertex capso-mers andprojections,
respectively.
Therelation-ship between the occurrence of
incomplete
HA andtoxinactivity
of virus materialcouldnot beincludedin the present study since no activity of
the latterkind was demonstrable even in highly
concentrated virus preparations of type 9. In further analogy with results obtained in studies of type 3 (11), it appears likely that the slowly sedirrenting HIC test positive type 9 component, also identified in CF tests with an
anti-HA serum, represents fiber components.
The slight aggregation of erythrocytes incubated
with the presumed isolated fiber components in the presence of heterotypic antiserum is a
phe-nonrenon whichrequires further analysis.
Fiber components, when forming a part of the complete HA, exhibited a length of about 12 to 13 m,u. It is ofconsiderable interest to note that
this value suggests the occurrence of one more
dirrensional class of fiber components in
addi-tion tothose hitherto identified. Previous studies
have demonstrated projections with a length of
about 10 m,u in adenovirus types 3 (9) and 11
(Norrby, J. Gen. Virol., in press), about 17 m,u
intype 4 (18), and about 25
m,i
in types 2 (Pet-tersson et al., personal communication) and 5 (17, 21). Theseclasses with different fiber lengthsconform with adenovirus subgroups proposed by
Rosen (14), the only exception being the anoma-lous memberofsubgroup III, type 4.
The group-specific CF antigen of type 9 was
carried by soluble components different from
thosediscussedabove, asinferred fromresults of
fractionation by anion-exchange
chromatography
or gel filtration on Sephadex G200 (Norrby,
unpublished data). It appears likely that this
group-specific CF antigen, like the
correspond-ing antigen oftypes 5 (17) and 3 (9), is carried
bynonvertex capsorrers, i.e., hexoncomponents.
ACKNOWLEDGMENTS
Theexcellent assistance of UlfSchonningin
connec-tion with electron microscopy studies and of Ulla
Carlsson and Birgitta Lindstrom in performance of
biologicaltestisgratefully acknowledged.
Thisinvestigationwassupportedbygrantsfrom the
SwedishMedicalResearchCouncil (project noK
67-16x-548-03 andno. B67-16x-74443C).
LITERATURE CITED
1. BAUER,H., ANDR. WIGAND. 1963.
Eigenschaften
der Adenovirus-Hamagglutinine. Z. Hyg.
Infektionskrankh. 149:96-113.
2. BAUER, H., R. WIGAND, AND W. ADAM. 1964.
Losliche und virusgebundene Adenovirus
Hamagglutinine. Z. Naturforsch. 19b:587-593.
3. BUcKLAND, F. F. 1959. Impairment of viral
haemagglutination of red cells aftertreatment
withformalin. Nature183:1276.
4. GELDERBLOM, H., R. WIGAND, AND H. BAUER.
1965. DEAE-chromatography of adenoviruses
of Rosen's group II.Nature205:625-626.
on November 11, 2019 by guest
http://jvi.asm.org/
5. GINSBERG,H. S., H. G. PEREIRA, R. C. VALENTINE,
AND W. C. WILcox. 1966.A proposed
termin-ology for the adenovirus antigens and virion morphological subunits.Virology 28:782-783.
6. KAsEL,J.A.,W. P.RowE,ANDJ. L.NEMES. 1960.
Modification of erythrocyte receptors by a
factorinadenovirus suspensions. Virology 10: 388-391.
7. KJmLLN, L. 1961. A study of adenovirus-host
cell system by the plaquetechnique. Virology
14:234-239.
8. KLEMPERER, H. G., Am H. G. PEREIRA. 1959. Studies of adenovirus antigens fractionated by chromatography on DEAE-cellulose. Virology 9:536-545.
9. NORRBY, E. 1966. The relationship between the
soluble antigens and thevirion ofadenovirus
type 3. I. Morphological characteristics,
Virology28:236-248.
10. NORRBY, E. 1966. Therelationship between the
soluble antigens and the virion ofadenovirus
type 3. II. Identification and characterization
of anincompleteHA.Virology 30:608-617.
11. NORRBY, E.,ANDP.SKAARET. 1967. The relation-ship between the soluble antigens and the
virion ofadenovirustype 3. III.Immunological
identification of fiber antigen and isolated
vertexcapsomerantigen. Virology32:489-502.
12. NORRBY, E., AND G. WADELL. 1967. Soluble
components of adenovirus type 4.Virology31:
592-600.
13. PEREIRA,H.G., A.C.ALLISON,ANDC.FARTHING.
1959. Studyofadenovirusantigens by
immuno-electrophoresis. Nature 183:895-896.
14. ROsEN, L. 1960. A hemagglutination
inhibition-technique for typing adenoviruses. Am. J.
Hyg.71:120-128.
15. SCHMIDT,W.A. K. 1967. Dieelectrophoretische
Trennbarkeit von Adenovirus Komponenten
im Dichtegradienten. Arch. Ges. Virusforsch. 20:11-19.
16. SVEDMYR, A.,J. F. ENDERS, AND A. HOLLOWAY.
1952.Complementfixation with Brunhildeand:
Lansing poliomyelitis viruses propagated in tissueculture. Proc. Soc. Exptl. Biol. Med. 79: 296-300.
17. VALENTINE, R. C., AND H. G. PEREIRA. 1965.
Antigens and structure of the adenovirus. J.
Mol. Biol. 13:13-20.
18. WADELL, G., E. NORRBY, AND U. SCHONNING.
1967. Ultrastructure of soluble antigens and
the virion of adenovirus type 4. Arch. Ges.
Virusforsch. 21:234-242.
19. WIGAND,R., AND M.STOHR. 1965.Thenature of
monkeycell haemagglutininsofRosen's
group-II ofadenoviruses. Z. Naturforsch. 20b:560-563.
20. WILCOX, W. C., AND H. S. GINSBERG. 1961.
Purification and immunological characteriza-tion oftypes 4and 5 adenovirus-soluble
anti-gens. Proc. Natl.Acad. Sci. U.S.47:512-526.
21. WILCox, W. C., H. S. GINSBERG, AND T. F.
ANDERSON. 1963. Structure of type 5
adeno-virus. II. Fine structure of virus subunits.
Morphological relationship of structural
sub-units to virus-specific soluble antigens from; infected cells. J. Exptl. Med. 118:307-314.
1108 J.