Design of
Retrovirus Vectors for Transfer and Expression of the
Human ,B-Globin Gene
A. DUSTY
MILLER,'*
M. A.BENDER,"12
EDITH A. S. HARRIS,' MICHAEL KALEKO,' AND RICHARD E. GELINAS'Department of Molecular Medicine, Fred Hutchinson Cancer Research Center, 1124 Columbia Street, Seattle,
Washington
98104,1
andDepartment
of Pathology, University
of Washington,
Seattle,
Washington
981952Received 15April 1988/Accepted 13 July 1988
Regulated expression of the human
0-globin
genehas beendemonstrated in cultured murine erythroleuke-mia cells and in mice after retrovirus-mediatedgenetransfer. However, the low titer of recombinant virusesdescribed to dateresultsinrelatively inefficientgenetransfer, whichlimitstheirusefulness for animal studies and forpotentialgenetherapy in humans for diseasesinvolvingdefective
0-globin
genes.We foundregionsthat interfered with virus production within intron 2 of the j3-globingeneandonboth sides of thegene.Theflanking regions could be removed, but intron 2was required for ,-globin expression. Inclusion of ,I-globin introns necessitatesanantisenseorientation of the genewithintheretrovirus vector. However, wefound noeffectof the antisense Il-globin transcription on virus production. A region downstream of theP-globin
gene thatstimulatesexpression of thegeneintransgenic micewasincludedinthe viruseswithout detrimentaleffectson
virus titer. Virus titersofover106 CFU/mlwereobtained with the final vectordesign, which retained the ability todirectregulated expression of human ,-globin in murineerythroleukemia cells. The vector also allowed transfer andexpression of the human ,I-globingeneinhematopoietic cells (CFU-S cells)inmice.
Anunderstanding of the
0-globin
geneand itscontroland the existence of a variety of human diseases caused bydefects in the
P-globin
gene have led to consideration of disease treatment by transfer of a normal gene into bone marrowof affected individuals. Retrovirusvectorscurrently offer the bestvehicle forgenetransferintomarrowcells, and vectorscarrying theP-globingenehave been usedtotransfer the,-globin geneinto murineerythroleukemia(MEL) cells, human erythroid progenitor cells (BFU-E cells), and bone marrow ofmice. MEL cellsare arrestedatarelatively latestage of erythroid development, but can be induced to differentiate by using a variety of inducers (9, 18). During differentiation, a large increase in mRNA and protein pro-duction from the endogenous globin genes is observed. Following retrovirus-mediated transfer of the human ,B-globingene into MEL cells, mRNAandprotein production from theintroduced human 3-globingene was inducible (4, 7, 13, 15) and intwostudieswasmadeatlevelsapproaching that of the endogenous mouse genes on a per-gene-copy basis (4, 13). Transfer of the
P-globin
gene into human erythroid progenitor cells (BFU-E) resulted in production of RNAfrom the transferred humangene at5% of the levelof theendogenousgene (4). Retrovirusvectors havealso been used totransfer the human P-globin gene into mouse bone marrow. Infusion ofinfectedmarrowintolethally irradiated mice resulted in tissue-specific expression of the gene in erythroid cells, albeit atlevels 100-fold lowerthan those of theendogenous mousegenes(8).One problem with current vectors which severely limits theirusefulness istheirrelatively low titer, from 7 x 103to 5 x
105
CFU/ml(4,7, 8, 13, 15). The low frequency ofgene transfer into bone marrow cells capable of reconstitutingmice(18 DNA-positive animals of 104 tested) (8) isapossible consequence of this problem. Amphotropic retroviral
vec-tors have been used to transfergenes into human
hemato-poietic progenitorcells,including erythroidprogenitors,but
* Corresponding author.
only the highest-titer vectors (106 to 107 CFU/ml) yield useful frequencies of transfer (11, 12). Indeed,wefindvery poorinfection of human BFU-E cells by usingour
P-globin
vector(4). Thus, development ofhigher-titerviruses
carry-ing the ,B-globin gene is requiredto facilitate gene transfer experiments and forpotential application of these techniques togenetherapy in humans.
Inthisstudy, we examined thefactorsleadingtothelow titer of ,-globin viruses and made alterations in the virus which resulted in a substantial increase in titer without
affecting'-globingeneexpression. In addition,we incorpo-rated into the viruses a region downstream of the human
P-globin
genethatisimportant for regulated humanP-globin
expression in transgenic mice (3, 14, 26). The new virusallowed efficient infection of murine hematopoietic cells (CFU-S cells) and human 1-globin expressionwas detected
in resultant hematopoietic colonies in mice.
MATERIALS AND METHODS
Cellculture.CellsweregrowninDulbecco modifiedEagle medium with high glucose (4.5 g/liter) supplemented with
10%calfserum
(*2
cells)or10% fetalbovineserum(all othercell lines). Concentrations of G418 are calculated by using
the weight of dry powder, of which about 50%wasactive. Previously described cell lines include PA317 (19) (ATCC CRL 9078), adenine phosphoribosyltransferase-negative
(APRT-) tetraploid MEL cells selected tobe semiadherent (gift ofP. Mellon, originally obtained from A. Deisseroth), thymidinekinase-negative (TK-) NIH 3T3 (20), and 42 (17). PA317retroviruspackaging cells used herewereeither from an early passage of the cell line or were reselected in
hypoxanthine-aminopterin-thymidine medium as described
previously (5). For induction ofdifferentiation, MEL cells
were seeded at 5 x 104 cells per ml in medium containing
3 mM N,N'-hexamethylene-bisacetamide (HMBA; Sigma ChemicalCo., St. Louis, Mo.)on day1. Onday 4, medium wasremoved from the cellsfollowing low-speed
centrifuga-tion andthe cellswere resuspended in the samevolume of
4337 0022-538X/88/114337-09$02.00/0
Copyright C1988, American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 10, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 10, 2019 by guest
http://jvi.asm.org/
(2163) (2422) (2482) (3287) Pstl Pstl AvrIl Xbal
I
i
l
II
I 11
I
_HP-VIRUSES
F-SX-VIRUS SA-VIRUSES
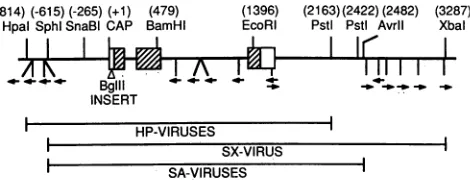
FIG. 1. Human P-globin gene and surrounding sequences.
Se-quencenumbersaregiven relativetothe RNAcapsiteat +1.
13oxes
indicate exons, and hatched regions indicate the protein-coding
region.ABglII site (AGATCT)wasinserted between bases+40and
+41,asindicated,tomarkthegene(4). Landmark restriction sites aregiven, andconsensus poly(A)signals (AATAAA) and orienta-tion of the signalsareindicatedbyarrows.Insertsusedin different vectorsarediagrammed below the--globingene.
Retrovirus vectors. Retrovirus vectors expressing the neomycin phosphotransferase (neo) drug resistance gene wereusedto transferP-globingenes. The prefix pindicates
the plasmid form of the virus. Retrovirussequencenumbers are as described previously (27). The vector pLNL-XI:C contains theneogeneinserted intoaDNA clone ofMoloney
murine leukemiavirusinplace of Moloney murine leukemia
virus sequences from 1039 to 7673 and contains XhoI, HindIII, ClaI restriction sites between theneogeneand the 3' long terminal repeat (LTR) for insertion of additional genes(5). ThevectorpLNSL7 contains (in thedirection of transcription) the 5' LTR through base 544 from Moloney murine sarcoma virus, bases 569 to 1038 from Moloney murineleukemiavirus,toBglII-to-SaII fragment from
trans-poson Tn5 containing the neogene (2), aPvuII-to-HindIII fragment from simianvirus40 containingthesimianvirus 40
early promoter, unique StuI, AvrII, HindIII, and ClaI sites for inserting cDNAs, and bases 7764 through the 3' LTR from Moloney murine leukemia virus. In addition, thegag
startcodon in pLNSL7 waschanged from ATGtoTAG to prevent possible gag protein translation (5). Both
LNL-XHC and LNSL7 were produced at equal titer by using retrovirus packaging cell lines.
3-Globingenes modifiedby insertion ofaBglII site as a
transcriptional marker (Fig. 1) (4)wereinserted into pLNL-XHC between the neo gene and the 3' LTR (pLNB*HP, pLNB*HP
MG',
and pLNB*HP MG-) and into pLNSL7 between theneogeneandthe 3' LTR inplaceof the simian virus 40 promoter (all other ,-globin vectors). Retrovirus vector names indicate the order ofgenes in the virus and important features of the vector. For example, LNB*HP indicates a retrovirus vector consisting ofa viral LTR (L) driving neo (N) followed by a transcriptionally marked ,-globin gene (B*) contained in an HpaI-to-PstI (HP) ge-nomic fragment. The vectors LNB*MG+, LNB*MG-, and LNB*WT-describedinapreviouspublication (4)have been renamed LNB*HPMG',
LNB*HP MG-, and LNB*HP, respectively, to allow consistent nomenclature use in thisreport. To make LNB*SA RP (reversed promoter), the SnaBI (-265)-to-BgIII (+43) fragment containing the ,3-globin promoter(Fig. 1) in LNB*SAwas excised and
rein-serted inreverseorientationby usingaBgIII linkertoadapt the blunt-endSnaBI sitestotheBglIIsites. TheBglIIsite is notpresentinthenormal 3-globingenebut ispresentinthe marked gene (Fig. 1). To make LNB*SA P- (promoter minus), the same fragment was removed from LNB*SA entirely, again using a BglII linker to join the SnaBI and BglII sites.
Generationof virus from retrovirus vector constructs. Virus
was generated from plasmid constructs as previously de-scribed(22). Briefly, plasmids containingthe viral constructs
were transfected into p2 ecotropic retrovirus packaging cells,and after 2daysviruswasharvested and used to infect PA317 amphotropic retrovirus packaging cells. The cells
werethen seeded into selectivemedium,andclonal celllines containing single integrated proviruses were isolated. The structures ofintegrated P-globin viruses were analyzed by Southern analysis (16) with a ,-globin minigene probe. By
usingrestriction enzymes that cleave only in each LTR of the virus(KpnI)andotherenzymesthat cutoncewithin and at sites presentat both ends of the 3-globininsert (BamHI andHindlll),weconfirmed that thefragmentsizesproduced by the integrated provirus matched those of the original plasmid construct.
Virus assay. Virus was harvested from virus-producing cells by incubating confluent dishes of the cells with fresh
medium for 16 h and thenremoving the medium and sub-jectingit tocentrifugationat3,000 x gfor 5minto remove
cells and debris. Forassayof virusescarryingtheneo gene, recipientcellswereseeded at5x 105per60-mm dishonday
1.Onday 2,themediumwaschangedto mediumcontaining 4 ,ug ofPolybrene perml andvarious amountsoftest virus
wereadded. Onday 3, the cellsweresplit1:10 into medium containing1mgofG418 (about50%active)perml. Colonies
were stained and counted on day 9. Amphotropic helper virus was measured by the S+L- assay as previously
described (20). The presence of helper virus in mice was
monitored by using the XC assay (23) as previously de-scribed(20), exceptthatrecipient NIH3T3 TK- cellswere
cocultivated with 10
[lI
of fresh blood instead of virus-containing medium for 16 h and blood cells were washed fromtheplate beforethe cells were trypsinizedfor the XCassay.
Infection of murine bone marrow cells and CFU-S assay. 5-Fluorouracil (150 mg/kg; LyphoM'ed Inc.) was adminis-tered intravenously to 6- to 8-week-old female C57BL/6J mice(Jackson Laboratory, BarHarbor, Maine). Micewere
sacrificed48'hlaterbycervicaldislocation,andmarrowwas
flushed from femurs and tibias by using cocultivation
me-dium (Iscoves medium with 10% heat-inactivated fetal
bo-vine serum [Hyclone Laboratories, Logan, Utah], 10% WEHI-3cell conditionedmedium,4,ugofPolybreneperml, penicillin, and streptomycin). Marro'w wascocultivated on
subconfluent monolayers of irradiated (2,100 rads) virus producercells for24 h.Hematopoieticcellswerewashed off
themonolayerand cultured for 36 to 48h in the presence of
1 mgofG418 per ml. Marrow was washed with and
resus-pended in Hanks buffered saline solution withoutcalcium, magnesium,orphenolred andinjectedintofemaleWBB6F1/ J-W/Wv mice (Jackson Laboratory) which had received
either no irradiation or400 rads. At 12 days postinjection, individual CFU-S colonies were dissected out and
dissoci-ated and total nucleic acid was isolated(4).
RESULTS
LNB*HP viruses: j8-globinviruses containing the HpaI-to-PstIgenomic fragment. Figure1 depictsthe human ,-globin gene and surrounding sequences. Before insertion into
vi-ruses, the ,-globin gene was modified by insertion of a
6-base-pair BglIIsite into the nontranslatedregionofexon1 (Fig. 1). This wasdone to allow measurement of transcrip-tion from the gene in human cells already expressing the endogenous ,-globin gene. When we began these
experi-(-814)(-615) (-265) (+1) (479)
HpalSphI SnaBi CAP BamHI
A
K_
m~IIER
r.
Bglll - .
INSERT
(1396) EcoRI
7
on November 10, 2019 by guest
http://jvi.asm.org/
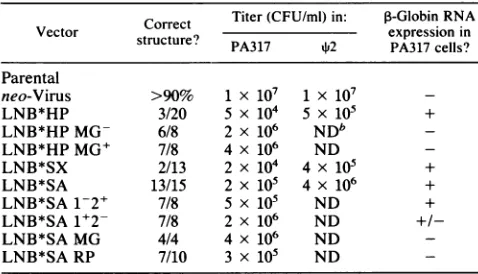
[image:2.612.70.305.70.160.2]TABLE 1. Characteristicsof PA317 cells containing ,-globinvectorsa
Titer(CFU/ml)in: P-GlobinRNA
Vector Correct expression in
structure? PA317
*2
PA317 cells?Parental
neo-Virus >90% 1 x 107 1 X 107
LNB*HP 3/20 5 X 104 5 X 105 +
LNB*HPMG- 6/8 2 x 106 NDbb
LNB*HPMG+ 7/8 4 x 106 ND
-LNB*SX 2/13 2 x 104 4 x 105 +
LNB*SA 13/15 2 x 105 4 x 106 +
LNB*SA 1-2+ 7/8 5 x 105 ND +
LNB*SA 1+2- 7/8 2 x 106 ND
+/-LNB*SA MG 4/4 4 x 106 ND
-LNB*SA RP 7/10 3 x 105 ND
-a Clon-alPA317 celllinescontaining the indicated retrovirusvectors were
generatedasdescribed in Materials and Methods. The number of cloneswith
the correctproviral struicture is indicated as the ratio of clones with the correct structure tothe totalanalyzed. The highest titer obtained fromPA317 cellscontaininganunrearrangedvirus is indicated for eachvector.Synthesis ofP-globin RNA in PA317 cells containing each vector is indicated, as
determined byNorthern analysis withaP-globinprobe. Virus from PA317
cells was used toinfect s2cells, infected cloneswereisolated, andthe best
titer obtained from%P2cellscontaininganunrearrangedprovirus is indicated.
Atthetime ofassay,the
q$2
cellswerehelper virus free (<10PFU/ml), asmeasured by theXCassay (20,23).After continuedpassage,helper viruswas
detected,ashaspreviously been reported fortheparental neo-virus (22).
bND,Notdetermined.
ments, the region from HpaI (base -814) to PstI (base
+2163) appeared to be sufficient for developmentally regu-lated tissue-specific expression of the gene in transgenic mice, andweinserted this fragment intoa selectable retro-virus in reverse orientationwith respect toviral transcrip-tion. We also inserted the same fragment containing an
intronless
3-globin
gene, or minigene, into the selectable retrovirus in both orientations. Virus generated by usingPA317 retroviruspackagingcells(19) was madeathightiter from vectors carrying the 3-globin minigene in either the
forward (LNB*HP
MG')
or reverse(LNB*HP
MG-) orien-tation, while the vectorcontaining the ,B-globin gene in the reverse orientation (LNB*HP) yielded virus with a much lower titer (Table 1) (4). Conversely, transcription ofP-globin was only detected in fibroblasts, MEL cells, or
BFU-E colonies infected with the vector
carrying
theP-globin gene with introns and not in cellsinfected with either
vectorcarrying a ,-globin minigene (4). Thus, transfer ofa transcriptionally active,B-globingeneappearedtobe
incom-patible with producing high-titervirus.
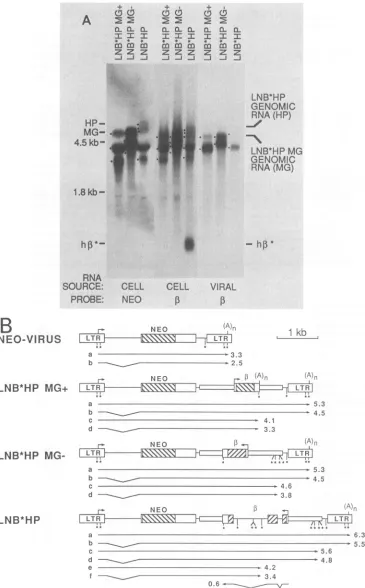
To understand how thedifferent,-globin inserts affected theproduction of virus,weexamined RNAfromPA317cells containing the different vectors and fromvirions produced by these cells (Fig. 2A).
PA317
cells infected with theparental neo-virus contained two predominant viral RNAs
(depicted in Fig. 2B), a full-length viral RNA which is
packaged into virions and a spliced RNA which is not
packagedbecause it lacks the signal requiredforpackaging
(1, 5). In contrast, PA317 cells infected with the ,B-globin viruses contained multiple viral RNAs, and virions produced
by the cells contained atleast two RNAs, of which only a minorportionwasthefull-length viral RNA(Fig. 2A). The
effectsontiter thatweobserved(Table 1) correlated with the
proportion of full-length viralRNAobserved in cells and in virions, which was reduced for the minigene viruses and was
barelydetectablefor the LNB*HP virus(Fig. 2A). Thus, the
low titer ofthese
P-globin
viruses was apparently due to thelow abundance of
full-length
viral RNA invirus-producing
cells and in virions
produced by
the cells.To understand the reasons for the low abundance of full-lengthviralRNA,we
analyzed
thetranscription
patterns of the vectors, and we propose thetranscription
patternsshownin
Fig.
2B toexplain
the RNA data inFig.
2A. Thetranscripts
found in PA317 cellsharboring
the LNB*HPMG'
virus can be correlated with knowntranscriptional
signals
in the virus. There are twopoly(A)
signals
in thisvirus, onein the 3' LTRandone at the endofthe
3-globin
minigene
wherepolyadenylation
of3-globin transcripts
nor-mally occurs[signals
labeled(A)n,
Fig. 2B].
Inaddition,
splicing
can occurbetween the 5' LTR andneosequencesin theparental
neo-virus(1,
5).
Thus,
we expect four viraltranscripts
in cellscontaining
LNB*HPMG'
(Fig. 2B).
Indeed,
threebandsweredetected in PA317 cellscontaining
the LNB*HP
MG'
virus(indicated
by
dots inFig. 2A)
corresponding
inorderofdecreasing
sizetoexpected
RNAs a,bplus
c, and d(Fig.
2B),
assuming
that theabundant28S rRNA(4.5
kilobases[kb])
caused bands b and c tocomi-grate.
Analysis
ofcellularRNAwithneoor,-globin probes
revealed a similarpattern
of bands. The sizes of the two RNA speciesinvirions wereconsistent withexpected
pack-aging
ofunspliced
RNAsaandc(Fig. 2B).
Thus,
viral RNAspecies
found in cellscontaining
LNB*HPMG'
and in virionsproduced
by
the cells match thepredicted
RNAspecies
depicted
inFig.
2B. Inaddition,
since RNAcorre-sponding
to thefull-length
viraltranscript
was observed incells and in
virions,
this indicates thattranscription
canoccuracrossthe
P-globin
poly(A)
signal
presentin the virus. In the LNB*HP MG-virus,
the,B-globin minigene
is inserted inreverseorientationwith respecttoviraltranscrip-tion.Noknown
poly(A)
orsplicing signals
arepresentin the insertedP-globin
sequences, but there is one consensuspoly(A)
signal (AATAAA)
in the3-globin
geneandfoursuchsignals
clustered at the 3' end ofthe insert(asterisks,
Fig.
2B). 1-Globin
andneoprobes
revealedtranscripts
of similarsize in LNB*HP MG--infected cells
(indicated
by
dots inFig. 2A)
whichcorrespond
in order ofdecreasing
size toproposed
transcripts
a, c,b,
and d(Fig. 2B), although
as noted above theseparation
ofbands b and ciscomplicated
by
theirproximity
tothe 4.5-kb rRNA band. Twotranscripts
weredetected invirions harvested fromPA317 cellscontain-ing
LNB*HP MG-(Fig.
2A),
and their sizes matchedproposed
transcripts
aand c(Fig. 2B). Thus,
RNAspecies
found in PA317 cells containing LNB*HP MG- and in
virions
produced by
the cells can be accounted forby
splicing
known to occur in theparental
neo-virus
and the presence of an additional RNA terminationsignal
in theP-globin insert,
which coincides with several consensuspoly(A)
signals.
Note thatanalysis
of RNA in LNB*HPMG'
and LNB*HP MG- virions revealsequivalent-size
bands which
correspond
to thepredicted full-length
viraltranscript,
asexpected
since the virusesare the samesize,
anddifferent-sizesmaller bandscorresponding
toearly
RNAterminationat different
positions
in thetwoviruses.Finally,
onemight
expectthat the patternof RNAspecies
observed fortheLNB*HP virus would be similartothatof
LNB*HP
MG-,
but shifted up 1 kb in size to reflect the additionof the3-globin
introns. The observed pattern wasmore
complex
(Fig. 2A),
but consistent with premature RNA termination within intron 2 of the3-globin
gene(Fig. 2B).
Proposed
RNAs e and f(Fig. 2B)
predominated
in PA317 cellscontaining
the virus(lower
twodots,
Fig. 2A),
and RNAspecies
e wasthemajor
RNAinvirionsproduced
from these cells(lower dot, Fig. 2A). Thus,
it appears thaton November 10, 2019 by guest
http://jvi.asm.org/
(I
A
22
0 0-0m
*
ror
*7 7j
z
1.8kb
-
hJ*-RNA SOURCE:
PROBE:
CELL
NEO
+ +
(50O (50(
(
25
2 2CL 0-Q n- a- L1
co comm coI
Z Z Z Z Z Z
OoQJ Qn DJ
.
9
CELL VIRAL
p
B
NEO-VIRUS
a
b
rN Eo(A)n
TRf
I1ci
I~~~~~~I
1 kb
2.5
N A) (A)1
=~~~~~~~~~~~~~~~~~~
LNB*HP MG+
a
-b
c 4.1
3.3
5.3
4.5
~~~(A'1
IIT
LNB*HP
MG-a
b
c 4.6
3.8 d
5.3 4.5
LNB*HP
a
b
c
d
e
5.6 4.8 4.2
3.4
6.3 5.5
[image:4.612.126.491.59.647.2]0.66
FIG. 2. Analysis of RNA from,-globin vectorscontaining the Hpal-to-PstI ,-globin fragment. (A) RNAsfromPA317cells containing
P-globinvectorsandfrom virionsproduced by the cellsweresubjectedtoelectrophoresis inagarosegelscontaining formaldehyde and probed
by usinganHpaI-to-PstIp-globinminigene probeor aneoprobe bystandard methods (16). The positions of the human,-globinmRNA(hP*), thefull-length RNAs from the minigene viruses (MG), and the full-lengthRNAfrom theintron-containing,B-globinvirus (HP)areindicated.
Positionsofthe28S (4.5 kb) and 18S (1.8 kb)rRNAsareindicated,andspecific bands discussed in thetextareindicated by dotstotheleft
of eachlane.(B) Proposed RNA speciestranscribed from retrovirusvectors.Approximate sizes of theRNAsaregiven in kilobases assuming
a200-base-pair poly(A) tail.Asterisks indicateconsensuspoly(A) signals (AATAAA)inthetranscriptional orientation of the viralpromoter,
(A)nindicatesknownpoly(A) sites in the orientation of the viralpromoter,andarrowsindicatepromoters.
LNB*HP GENOMIC RNA
(HP)
LNB*HP MG GENOMIC RNA (MG)
-
hp*
d
(A)
r-ol NEO 11 ri
LTR,.
d.No\"NINIIINI. ri AFm-a
71*1 *
H p
-
MG-4.5
kb
on November 10, 2019 by guest
http://jvi.asm.org/
x
I
mm
z z
I
r-t---4.5kb
-
1.8kb
-PROBE
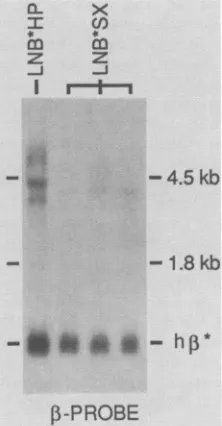
FIG. 3. Comparison of RNA in PA317 cellscontainingLNB*HP
orLNB*SX.Northern (RNA)analysis wasperformedasdescribed
in the legend to Fig. 2A. One PA317 clonal cell line containing LNB*HP and three independent clones containing LNB*SX were
analyzed. Thesizesof the ribosomal markers and thepositionof the
human 3-globin(h,*)RNAareindicated.
sequencesin intron2in thereverseorientation
P-globin
geneact toreduce the amountof full-length viral RNAin PA317 cellscontaining theLNB*HP virus and in virions produced by these cells. RNAwith the size expectedfrom the
P-globin
gene was detected in cells containing this virus. This
P-globin transcript was not detected in virions produced by these cells, even though it is transcribed in an antisense
orientation with respect tothe viralgenome and thus could hybridize to viral RNA. In summary, the low titer of the
P-globin
viruses appears to be due to early termination of viral transcription, which reduces the amount offull-length viral RNAin virions.LNB*SX: a
0-globin
virus containing the SphI-to-XbaIgenomic fragment. Inanattempt toimprove the titer ofvirus containingthe,-globingene, weconstructedavirus thatdid
not contain the HpaI (-814)-to-SphI (-615) region of the
gene which contained the four consensus poly(A) signals
(Fig. 1), in which early termination of RNA transcription appeared to occur. In addition, evidence had accumulated
suggesting involvement of sequences downstream of the
gene in ,B-globin gene regulation (3, 14, 26). Thus, we
included these sequences in the LNB*SX virus, which
contains the region from SphI to XbaI inserted in reverse
orientation intoaneo-virus. PA317celllines containingthis
vector were generated and screened as for the LNB*HP
virus. Unfortunately,weobservedratesofrearrangementof theLNB*SX virus inPA317cellsthatwere ashigh asthose
seen with LNB*HP. In both cases, the rearranged viruses
were smallerthan theparental virus andhad suffered
dele-tions of 3-globinsequences.ThebesttiterfromaPA317cell
clonecontaininganunrearrangedLNB*SX proviruswas2 x
104, evenlowerthatobtained with LNB*HP(Table 1).
Analysis of RNA in PA317 cells containing either the LNB*SXorLNB*HP virus(Fig.3)providedanexplanation
for the low titer of theLNB*SXvirus. Transcriptsfromthe
viral LTR through the ,B-globin gene were reduced in
LNB*SX-infected cells in comparison with those in
LNB*HP-infected cells. In contrast, the amount of RNA
transcribed from the
,-globin
gene was similar in cells infected with either virus. Since theprobe
used in theseexperimentswas a
3-globin
minigene
probe
(positions
-814 to +2163, Fig. 1), it islikely
that an additional RNA termination signal in the PstI-to-XbaIfragment
present inLNB*SX, possiblythe
poly(A)
signal
atposition
+2637(Fig.
1), leadsto terminationbefore the
region
recognized
by
the probe. Apparently, the titer ofthe LNB*SX virus wasnot improvedcomparedwith that ofLNB*HPbecause,
whilewe may have removed one terminationsignal,
we introducedanother.
LNB*SA: a
0-globin
viruscontaining
theSphI-to-AvrII
genomic fragment.
Assuming
that thepoly(A)
signal
atposition +2637
(Fig. 1)
wasresponsible
for the low titer of the LNB*SXvirus,
we deleted thissignal
in our next,B-globin
virus,
LNB*SA,
which contains theSphI
(-615)-to-AvrII(+2482)
region
containing
theP-globin
gene(Fig.
1)
in reverse orientation. This virus still contains the
region
downstream ofthe3-globin
gene involved inregulation
of the gene, whichhasbeenlocalizedbetweenbases+2163 and +2422(3). Incontrasttoresults obtainedwithLNB*HPandLNB*SX, most of the PA317 cell lines infected with
LNB*SA virus contained
unrearranged
proviruses,
and thehighest titer observed was 10-fold
higher
than that of LNB*SX and4-foldhigher
thanthat of LNB*HP(Table
1).
Therefore, we conclude thatsequences inthe
regions
flank-ingthe
P-globin
genefrompositions
-814to -615 andfrompositions +2482 to +3287 inhibit virus
production
when inserted into aretrovirusinreverseorientation. RNAanal-ysissuggeststhatthis resultisduetoprematuretermination
of RNA
transcription,
which reduces the amount offull-length viral RNA and thus also reduces the titer of virus produced.
Role of
I-globin
introns in LNB*SA virus. The titerob-tained from theLNB*SAvirus
(2
x105
CFU/ml)
wasbetter than that of other virusescontaining
aP-globin
gene with introns but was still far from the titer obtained with theminigeneviruses(2 x
106
to4 x 106CFU/ml).
Todetermine which intron wasresponsible
for the decreasedtiter,
we constructed three new viruses with structures identical to thatof LNB*SA exceptthat LNB*SA 1-2+ lacked intron1,
LNB*SA
1+2-
lacked intron2,
and LNB*SA MG lackedboth
P-globin
introns. Most of the PA317 linesgenerated
fromthesevirusescontainedunrearranged
proviruses,
as we found for theparental
LNB*SA virus(Table 1).
The mini-gene virus LNB*SA MG(which
does not containP-globin
introns)hadthehighest
titer,
20-foldhigher
thanLNB*SA,
a result similartothat obtainedpreviously
with the LNB*HPviruses (Table 1). Virus titersfrom the LNB*SA 1-2+ and
LNB*SA 1+2- viruses were intermediate to those of LNB*SA and LNB*SA
MG,
butclearly
the presence ofintron 2 best correlated with reducedvirus titer
(Table
1).
Wenext
analyzed
the RNA in the PA317 cellscontaining
LNB*SAanditsderivatives(Fig.
4).
Atranscript
of the sizeexpected
from the0-globin
gene was present inLNB*SA-andLNB*SA 1-2+-infected cells butnot inLNB*SA 1+2--or LNB*SA MG-infected PA317 cells.
Quantitation
ofP-globin
transcripts
by
an RNAprotection
assay revealed similar amounts in LNB*SA- and LNB*SA1-2+-infected
cells, 7% of this level in LNB*SA 1+2--infectedcells,
andonly 1%in LNB*SA MG-infected cells.
Thus,
transcription
of the3-globin
genestrongly
depended
on the presence ofP-globin
intron 2.Expected
full-length
proviral
transcripts
(indicatedbydots inFig. 4)
wereabundant in LNB*SA MG-and LNB*SA 1+2--infectedcells,
weregreatly
reduced inon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.128.239.74.287.2]1-2- 1+2- 1-2+ 1+2+
-4.5kb
-1.8kb
-hp
A
-PROBE
FIG. 4. Northern analysis of RNA in PA317 cells containing LNB*SAanditsderivatives lacking one or more introns. RNA from independentPA317 clonal cell linescontainingLNB*SAMG (1-2-), LNB*SA 1+2- (1+2-), LNB*SA 1-2+ (1-2+),andLNB*SA(1+2+) wasanalyzed as described in the legend to Fig. 2A. The sizes of rRNAs and the position of the human
P-globin
mRNA(hp*)
are shown, andthefull-length viral RNA species for each cell line is indicated byadottothe left ofeachlane.LNB*SA1-2+-infected cells, and were barely detectable in
LNB*SA-infected cells. As anticipated, the amount of
full-length viral transcript present (Fig. 4) correlated with the titer ofthe virus(Table 1). Finally, the amount of full-length
viral transcript, and thus the titer of virus, was inversely
related to the amount of ,-globin message present in the
virus-infected cells.
Effect of
,3-globin
gene transcription on virus production. Theresults presented above lead to theunfortunateconclu-sion thatthe titerof virusescontaining the
3-globin
gene in reverse orientation is inversely related to the ability ofthe virus to make a3-globin
mRNA. We considered threepossible explanations for this effect. (i) The introns
them-selves, in particular intron 2, contain signals that inhibit
production of thefull-length viral RNA; (ii) the presence of anactivepromoterinterfered withviraltranscription; or (iii) thepresence of the,B-globin mRNA, which has an antisense
orientation with respect to viral mRNA, caused rapid
deg-radation oftheviral RNA or in some other manner affected
accumulationof viral RNA. To test the latter two
possibili-ties,wealtered LNB*SAtopreventproductionof antisense RNA. In LNB*SARPthe ,-globin promoter was reversed, and in LNB*SA P- the promoter was removed entirely.
PA317 celllines containing LNB*SA RP were made and screened asdescribed above. Both the percentage of clones
containing an unrearranged virus and the titer of virus produced from the best clone were similar to results ob-tained with the parental LNB*SA virus (Table 1). As
ex-pected, RNA ofthe size expected from the ,B-globin gene was not detected in PA317 cells containing LNB*SA RP by using a ,-globin probe (data not shown). Viral transcripts detected were similar in sizes and abundance to those of LNB*SA (data not shown). Thus, the low titer of the LNB*SA virus was not due to reverse orientation
transcrip-tion(withrespect to viraltranscription) of the
3-globin
gene orto the presence of ,(-globin RNA in the virus-producing cellswhich has an antisense orientation with respect to viraltranscripts.
Next, we tested the ability of pLNB*SA P- to make high-titer virus. For this assay, we used a transient virus production assay in which retrovirus-packaging cells are transfected with the virus constructs, virus is harvested 2 days later, and the titer of the virus is measured. This assay is much faster than the assayused for the virusesdescribed
sofar, and we found agoodcorrelation between the results of the two assays with ,B-globin viruses. The results show that pLNB*SA, pLNB*SA RP, and pLNB*SA P- all pro-duce virusat atiter that is atleast 20-fold lower than that of pLNB*SA MG or the parental neo-virus (Table 2). That the titer of LNB*SA P- was not improved indicates that the presence of an active ,-globin promoter in LNB*SA is not responsible for the low titer of this virus. Therefore, we conclude that the inverse correlation between ,-globin
expression and virus titer observed for the LNB*SA, LNB*SA
1+2-,
LNB*SA 1-2+, and LNB*SA MG viruses does not reflect inhibition of virus production as a result ofP-globin
gene transcription. Instead, we conclude that ,-globin intron 2 contains signals that inhibit transcription of the full-length viral RNA, which results in reduced virus titer.I-Globin
virusproduction
from*2
retrovirus-packaging
cells. We tested the ability of another retrovirus-packaging cellline,qi2
(17), to produce several of the vectors. Interest-ingly, the titers of ,B-globin viruses LNB*HP, LNB*SX, and LNB*SA were 10- to20-foldhigher from 4i2 cells than from PA317 cells, even though the highest titer of the parental neo-virus was similar in both cell lines(Table 1). The titer of LNB*SA was 10-fold higher than that of LNB*HP orLNB*SX, consistent with results found with PA317 cells. Helper virus was not detected in any of these clonal lines at thetimeofassay and thuscannotexplainthehigh titerof the
,B-globinvectors.
Expression of j8-globin gene in infected MEL cells. We
infected MELcells with severalof the ,-globin virusesand measured expression of the transferred
P-globin
gene inuninducedandinducedMEL cellsby usingan RNA protec-tion assay(Table3). We found that all viruses containing the
P-globin
gene with both introns as well asthe viruscontain-ing only intron 2 (LNB*SA 1-2+) directed similar amounts of correctly initiated ,-globin RNA synthesis in induced cells. In contrast, ,-globin RNA was reduced more than 10-fold in MEL cellsinfectedwith theviruscontainingintron 1but not intron 2(LNB*SA 1+2-) and was not detected in MEL cells infected with the minigene virus. These results
parallelthose obtained with fibroblasts(Fig. 4)and showthat
intron 2 is required for efficient
P-globin
expression. Inaddition, ,-globin RNA was induced to similar extents in MEL cells containing all the viruses except for LNB*SA
MG,inwhich
P-globin
RNAwas not detected. Thus,either [image:6.612.76.272.72.252.2],-globinintroncanbe deletedwithout
affecting
theability
ofTABLE 2. Effect ofP-globinpromoter alterations on virusproduction'
Vector Transient titer
(CFU/ml) pLNB*SA... 1 x104 pLNB*SARP... 6 x 103 pLNB*SAP ... 1 x 104
pLNB*SAMG... 2 x105
neo-Virus... 3 x 105
"Eachofthe indicatedvectors(10,ug)wastransfected intoP2cellsplated the day before at 5 x 105 per 6-cm dish by using calcium phosphate coprecipitationasdescribedpreviously(21).The cellswerefed 1dayafter
transfection,and after 2daysviruswasharvested andquantitated.
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 3. P-Globinexpressionin MEL cells' 1-GlobinRNA Avginduced
Vector [% of totalpoly(A)+RNA] RNA level Avg fold
comparedwith induction
Uninduced Induced LNB*SA(%)
LNB*HP 0.0073 0.066 110 9
LNB*SX 0.0031 0.039 70 13
LNB*SA 0.0044 0.058 100 13
LNB*SA1-2+ 0.0017 0.027 50 15
LNB*SA1+2- 0.0003 0.0023 4 8
LNB*SA MG <0.0002 <0.0002 <0.3
p-GlobinRNAproductionwasmeasured ininduced and uninducedMEL
cellsby anRNAprotection assay aspreviously described(4). Results are
averages of twoexperiments for each vector. Cells analyzedarepools of
G418-resistant MEL cells resulting from infection with virus fromPA317cells
containing the indicatedvectors.
the gene torespond toinduction. Finally, these results show that the presence of theadditionalregion downstream of the
P-globin
gene implicated in control of gene expression(which is present in LNB*SX and LNB*SA but not in
LNB*HP)had nosignificant effect on
P-globin
expressionin MEL cells.The presence of human ,-globin polypeptide in infected
MEL cells was determined byimmunofluorescence with an
antibody for human
f-globin
which does not recognizemouse
P-globin
(24). MELcells infected with the LNB*HPandLNB*SAvirusesdisplayedsimilarfluorescence,with 20 to 40%of the cells in these nonclonal populations showing bright fluorescence (data not shown). Thus, cells infected
with the high-titer LNB*SA virus make human 3-globin polypeptide in amounts similar to those demonstrated
pre-viously fortheLNB*HPvirus (4).
Expression of human I-globin in infected mouse hemato-poietic cells. We used the high-titer virus from
jp2
cellsproducing the LNB*SA virus to infect bone marrow from
mice. Infectedmarrow wasinfused intoW/WVanemic mice, and after 12days spleen colonieswereisolated and analyzed for the presence of vector DNA and for human
P-globin
RNA expression. Twelve-day CFU-S colonies arise from
primitive hematopoietic cells which give rise to colonies
containing various hematopoietic lineages, including
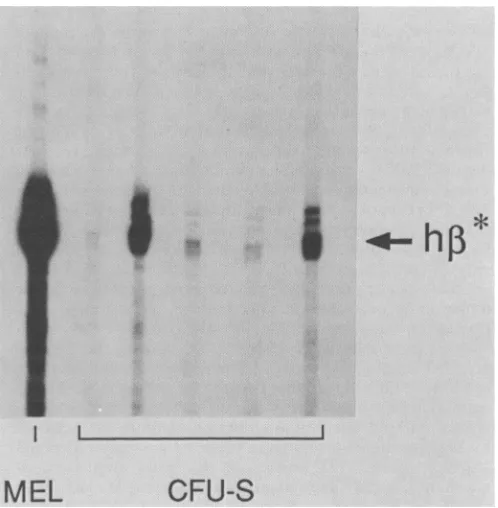
ery-throid cells. In separate experiments, 6 of9 and 10 of10 CFU-Scolonies containedaprovirus of theexpected struc-ture (data not shown). Every infected CFU-S colony
con-tained correctly initiated human ,-globin mRNA, although
the level ofexpression varied over100-fold(Fig. 5).CFU-S colonies containavariable mixtureoferythroidand nonery-throid cells, which may account for part of thevariation in
expression. However, we have no evidence that human
3-globin
expression is indeed erythroid specific in theseexperiments. No helper virus was detected in similarly
treated miceanalyzed after1 month, showing that infection was notthe resultof virus spread. Thus, LNB*SA was able to infect primitive hematopoietic cells and the transduced
globingene wasexpressed in theresulting colonies.
DISCUSSION
We made a
variety
ofalterations to increase the titer of viruses containing the human ,B-globin gene. Several signals in both intron 2 and regions flanking the3-globin
gene interfered with the retrovirus life cycle. We could eliminate theinhibitory flanking signals but not the signal(s) in intron 2, because the presence of this intron was required for:~~~
rt,I
ME
5.-hf3
CFU-S
FIG. 5. HumanP-globinRNA(h1*)synthesis in infectedCFU-S colonies. Bone marrow was cocultivated with
qj2
cells producing LNB*SA, selectedin G418, and infused into anemic W/WV mice. Twelve-day CFU-S colonies were isolated, and total nucleic acid fromprovirus-containing CFU-S colonieswasanalyzedbyanRNA protection assay (4). RNAfrom aMEL cell line infected with a ,B-globin gene-containingviruswasusedas amarker.efficient
P-globin
expression. For the virus containing a forward orientation,B-globin minigene (LNB*HPMG'),
theinterfering signalwas apparently the normal poly(A) signal
present at the end of the
P-globin
gene, which led to decreased full-length virus transcription. Less than full-lengthtranscriptionin other 3-globinviruses alsocorrelated withthe presence of consensuspoly(A) signals (AATAAA)within thevirus,butcouldbeduetoothersignals for3' RNA
processing orfor termination oftranscription. In addition,
notall consensus poly(A) signals are functional and notall functional poly(A) signals share the AATAAA consensus sequence (6); thus, the elimination of AATAAA signals mightnotbesufficienttoincreasefull-length viral
transcrip-tion.
Weexploredthe effecton
P-globin
expression ofaregiondownstream of the,-globingenethat has beenimplicatedin
3-globin
regulation, but found no effect on expression in MELcellsorin fibroblasts. Inaddition,limitedexperimentssuggestthat thisregionhasnoeffecton,-globin expression
inanimals reconstituted withinfectedmarrow(8). However,
inclusion of this region in the LNB*SA virus does not
appreciably affect virus titer; therefore, we retained the sequence. We hope that regions flanking the globin locus which are responsible for position-independent high-level
expression of
3-globin
in mice (10)can eventuallybeincor-poratedinto these vectors. At present, however, the local-ization of these elements is not precise enough to allow insertion of the region without exceeding retrovirus size limits.
Problems that have been encountered whenusing
retrovi-ral vectors to transfer genes can be grouped into three
categories: inappropriatesplicing, early termination of viral
L
Ion November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.318.567.71.325.2] [image:7.612.66.300.84.184.2]RNA, and competition of multiplepromoterscarried by the viruses (forareview, see reference 25). Since the
P-globin
intronswerefound tobe necessaryfor
P-globin
expression, we were limitedto insertion of thegene in reverseorienta-tion toavoid removal oftheintrons bysplicing during virus replication. Less than full-length viral transcription was encountered with reverse orientation inserts, which was reduced but not eliminated inredesigned viruses. We have direct evidence that possible competition between viral and ,-globinpromotersorpossibleeffectsofantisenseRNA did not affect virus titer, since reversal or removal of the ,-globin promoter had no affect on virus titer. The diffi-culties encountered with retrovirus-mediated transfer and expression of 3-globin contrast with the ease with which another transcriptionally regulated gene, a rat growth hor-mone minigene, was transferred and expressed after inser-tion into a retrovirus vector(21). Despite these difficulties, systematic modification of viruses containing the
P-globin
geneledtoa 3-globinviruswhichcanbemadeatarelatively high titer. Still higher titers might be achieved if the ele-ment(s)inintron 2 whichinterferes with transcriptioncanbe altered without affecting ,B-globin expression, for example, by site-specific mutagenesis of the presumed poly(A)signal. In generating PA317cell linescontaining 3-globinviruses,we found many lines containing rearranged proviruses
(Table 1), especially with LNB*HP and LNB*SX. These viruses were in general shorter than the parental virus and had suffered various deletions (some complete) of the
P-globin insert. Similar deletions were found in several inde-pendent clones, suggesting thateitheralternativesplicingor recombination in regions ofhomology was responsible. The rearranged viruses werealways ableto replicate toahigher titer than the original virus. For a given vector there was a correlation betweenthetransient titer of virus obtained from *J2 cells (e.g., Table 2), the highest titer that could be obtainedfrom PA317 cellsinfected withthevector(Table 1), and the proportion of PA317 clones analyzed that contained unrearranged proviruses (Table 1). We interpret thesere-sultstomean thatrecombinant viruses aregenerated during
transfection for all viruses, but only if the transfected virus replicatesverypoorlyarethe rearranged virusesdetectedat highfrequency in the infected PA317 cells. Once packaging cell lines containing single unrearrangedviruseswere
gener-ated, little rearrangement was detected when virus from these cells wasused to infect other cells.
The LNB*SA virus developed here canbe obtained from
4j2
cellsat4 x10'
CFU/ml, and thistiter allowed transfer ofthe
P-globin
gene into murine CFU-S cells. Twelve-day CFU-S cells share several characteristics with moreprimi-tive hematopoietic stem cells. They are present in bone
marrowatlowabundance, they cycle slowly, and they give
risetocellsofmultiplelineages. Weshowed thatupto100% of 12-day CFU-S cells can be infected and that every
infected CFU-S cell expresses correctly initiated human
,*-globin
mRNA. The high titer and ability ofthis virus toexpressthe 3-globingeneinvivo will assistfurther studies of
the long-term expression of the introduced P-globingene in
transplanted mice and in human hematopoietic cells.
ACKNOWLEDGMENTS
WethankCarol Buttimore fortechnical assistance, Thalia
Papa-yannopoulou for help with the immunofluorescence studies, and
Ronald Reeder forsuggestions concerning the manuscript.
M.A.B.wassupportedbytheMedicalScientist Training Program
ofthe National Institutes of Health. This study was supported by
Public Health Service grants CA41455, HL36444, HL37073, and AG00057 from the NationalInstitutesofHealth.
LITERATURE CITED
1. Armentano, D., S.-F. Yu, P. W. Kantoff, T. von Ruden, W. F. Anderson, and E. Gilboa.1987.Effect ofinternal viralsequences onthe utility ofretroviral vectors. J. Virol. 61:1647-1650. 2. Beck, E., G. Ludwig, E. A.Auerswald, B.Reiss, and H. Schaller.
1982. Nucleotide sequenceandexactlocalizationof the neomy-cin phosphotransferase gene from transposon Tn5. Gene 19: 327-336.
3. Behringer, R. R., R. E. Hammer, R. L. Brinster, R. D.Palmiter, and T. M. Townes. 1987. Two 3' sequences direct erythroid-specific expression of human,B-globingenesintransgenic mice. Proc. Natl. Acad. Sci. USA84:7056-7060.
4. Bender, M. A.,A. D. Miller, and R. E. Gelinas. 1988. Expres-sion of the human ,-globin gene after retroviral transfer into murine erythroleukemia cells and human BFU-E cells. Mol. Cell. Biol. 8:1725-1735.
5. Bender, M. A., T. D. Palmer, R. E. Gelinas, and A. D. Miller. 1987. Evidence that the packaging signal of Moloney murine leukemia virusextends into the gag region. J. Virol. 61:1639-1646.
6. Birnstiel, M. L., M. Busslinger, and K. Strub. 1985. Transcrip-tion termination and 3' processing: the endis in site! Cell 41: 349-359.
7. Cone, R. D., A. W. Benarous, D. Baorto, and R. C. Mulligan. 1987. Regulated expression ofacomplete human,B-globingene encodedby atransmissible retrovirusvector.Mol. Cell.Biol.7: 887-897.
8. Dzierzak, E. A., T.Papayannopoulou,and R.C.Mulligan. 1988. Lineage-specific expression of ahuman,-globingenein murine bone marrow transplant recipients reconstituted with retrovi-rus-transduced stem cells. Nature(London) 331:35-41. 9. Friend, C., W. Scher, J. G. Holland, and T. Sato. 1971.
Hemoglobin synthesis inmurineinducedleukemic cellsinvitro: stimulation of erythroid differentiation by dimethylsulfoxide. Proc. Natl. Acad. Sci. USA 68:378-382.
10. Grosveld, F., G. B. van Assendelft, D. R. Greaves, and G. Kollias. 1987.Position-independent, high levelexpression of the human 13-globin geneintransgenic mice. Cell51:975-985. 11. Hock, R. A., and A. D. Miller. 1986. Retrovirus-mediated
transfer and expression of drug resistance genes in human haematopoieticprogenitor cells. Nature (London)320:275-277. 12. Hogge, D. E., and R. K. Humphries. 1987. Gene transfer to
primary normal and malignant human hemopoietic progenitors using recombinant retroviruses. Blood69:611-617.
13. Karlsson, S., T. Papayannopoulou, S. G. Schweiger,G. Stama-toyannopoulos, and A. W. Nienhuis. 1987. Retroviral-mediated transfer ofgenomic globin genes leads toregulatedproduction of RNA andprotein. Proc. Nati. Acad. Sci. USA84:2411-2415. 14. Kollias, G., J. Hurst, E. deBoer, and F. Grosveld. 1987. The human ,-globin gene contains a downstream developmental specificenhancer. Nucleic Acids Res. 15:5739-5747.
15. Lerner, N., S. Brigham, S. Goff, and A. Bank. 1987. Human ,B-globinexpression aftergenetransferusingretroviralvectors.
DNA 6:573-582.
16. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning, alaboratory manual. Cold SpringHarborLaboratory, Cold Spring Harbor, N.Y.
17. Mann, R., R. C. Mulligan, and D. Baltimore. 1983.Construction of a retroviruspackaging mutant anditsuse toproduce helper-free defective retrovirus. Cell33:153-159.
18. Marks, P. A., and R. A. Rifkind. 1978. Erythroleukemic differ-entiation. Annu. Rev. Biochem.47:419-448.
19. Miller, A. D., and C. Buttimore. 1986. Redesign ofretrovirus packaging cell lines to avoid recombination leading to helper
virus production. Mol. Cell. Biol. 6:2895-2902.
20. Miller, A. D., M.-F. Law, and I.M. Verma. 1985. Generationof helper-freeamphotropic retrovirusesthattransducea dominant-acting, methotrexate-resistant dihydrofolate reductase gene. Mol. Cell. Biol. 5:431-437.
21. Miller, A. D., E. S. Ong, M. G. Rosenfeld, I. M. Verma, and
on November 10, 2019 by guest
http://jvi.asm.org/
R. M. Evans.1984.Infectiousandselectableretrovirus
contain-ing aninducibleratgrowthhormone minigene. Science
225:993-998.
22. Miller,A. D., D. R.Trauber, and C. Buttimore. 1986. Factors involved in the production of helper virus-free retrovirus
vec-tors. Somatic Cell Mol. Genet. 12:175-183.
23. Rowe, W. P., W. E. Pugh, and J. W. Hartley. 1970.Plaqueassay techniques for murine leukemia viruses. Virology 42:1136-1139.
24. Stamatoyannopoulos, G., M. Farquhar, D. Lindsley, M. Brice,
T.Papayannopoulou, and P. Nute.1983. Monoclonalantibodies specific for globin chains. Blood61:530-539.
25. Temin, H. M. 1986. Retrovirus vectors for gene transfer:
efficient integration intoand expression ofexogenous DNAin vertebrate cellgenomes, p. 149-187. In R. Kucherlapati (ed.), Gene transfer. PlenumPublishing Corp.,NewYork.
26. Trudell, M., and F.Costantini. 1987. A3' enhancercontributes to the stage-specific expression ofthe human
P-globin
gene.Genes Dev. 1:954-961.
27. Van Beveren, C., J. Coffin, and S. Hughes. 1985. Nucleotide
sequences complemented with functional and structural
analy-sis,p.766-782. In R.Weiss,N.Teich,H.Varinus,andJ.Coffin (ed.),RNAtumorviruses,vol. 2. ColdSpringHarbor Labora-tory,Cold Spring Harbor, N.Y.
on November 10, 2019 by guest
http://jvi.asm.org/
ERRATUM
Design of Retrovirus Vectors for Transfer and Expression of the Human
f3-Globin Gene
A. DUSTY MILLER, M. A. BENDER, EDITH A. S. HARRIS, MICHAEL KALEKO AND RICHARD E.GELINAS DepartmentofMolecularMedicine, Fred Hutchinson Cancer Research Center, 1124 Columbia Street, Seattle, Washington 98104, and
DepartmentofPathology, University of Washington, Seattle, Washington 98195
Volume 62, no. 11, p. 4337: The lastline of the right-hand column should read ". . . in the same volume of fresh medium plus 3 mM HMBA. Cells were harvested on day 6."