0022-538X/80/01-0340/09$02.00/0
Abelson
Murine Leukemia Virus Mutants with Alterations in
the
Virus-Specific
P120 Molecule
NAOMIROSENBERG`* AND OWENN.WITTE2
Cancer Research Center, Tufts University School of Medicine, Boston, Massachusetts 02111,1 and Cancer ResearchCenter,Massachusetts Instituteof
Technology, Cambridge,
Massachusetts021392Abelson murine leukemia virus (A-MuLV) is a replication-defective virus that
transformsbothfibroblastsand hematopoieticcellsin vitro. The virus encodes a
120,000-molecular-weight protein (P120) that is composed of Moloney murine leukemiavirus-derived gag gene sequences and A-MuLV-specific sequences. This
protein is the only A-MuLV-encoded protein that has been detected, and thus P120 is a candidate for the transforming protein of A-MuLV. We now report isolation and characterization of three new A-MuLV isolates that do not synthe-size P120 but do produce analogous proteins of larger (160,000 molecular weight)
and smaller (100,000 and 90,000 molecular weight) size. All of these A-MuLV isolates transform fibroblasts and lymphoid cells in vitro. Because the different A-MuLV proteins vary in the A-MuLV-specific region of the molecule, these variants may set a maximum limit on the size of the A-MuLV transforming
protein.
Abelson murine leukemia virus (A-MuLV) is areplication-defective, transforming retrovirus that arose in asteroid-treated BALB/c mouse
injected with Moloney murine leukemia virus (M-MuLV) (1). The virus induces a fatal
non-thymiclymphosarcoma3 to 4weeks after injec-tion into neonatal mice and transforms both fibroblasts and hematopoieticcellsin vitro (18,
21). A-MuLV stocks always contain a replica-tion-competent virus (such as M-MuLV) that provides the defectiveA-MuLV withenzymatic functions andstructuralproteins (9, 21).
The defective nature ofA-MuLV was dem-onstrated directly by Scher and Siegler (21)
when they isolated an A-MuLV-transformed nonproducer cell line called ANN-1. A-MuLV rescued from ANN-1 cells with cloned helper virus inducedtypical Abelson lymphosarcomas,
indistinguishable from the disease induced by
the uncloned virus prepared from tumor ex-tracts.ANN-i-derivedvirusthusbecamewidely
used and served as the prototype cloned
A-MuLV strain.
Heteroduplex mappingandSinuclease
map-ping studies using ANN-i-derived A-MuLV
show that the 5.6-kilobase A-MuLV RNA ge-nomehas tworegionsofprecise homologywith M-MuLV:a 730-baseregionatthe 3' end anda
1,320-baseregionatthe 5' end(D.Baltimore,A
Shields, G. Otto,S. Goff, P. Besmer, 0. Witte,
and N. Rosenberg, Cold Spring Harbor Symp. Quant. Biol., in press; A. Shields, G. Otto, S.
Goff, M. Paskind,and D. Baltimore, submitted forpublication). The 3.5-kilobase regioninthe
center ofthe A-MuLV genome is not
homolo-gous to M-MuLV, and hybridization studies
show that this region is not present in other replication-defective murine retroviruses(8;
Bal-timore et al., in press). These A-MuLV se-quences are present innormalmouseDNAand
probably represent a portion of the genome of the BALB/c mouse from which A-MuLV was
originally isolated.Because these sequences
dis-tinguishA-MuLVfromits M-MuLV parent and
other murineretroviruses, theymay be
respon-sible for the unique biological properties of
A-MuLV.
Analysisof theproteins expressed inANN-1
cells and A-MuLV-transformed cellsderived us-ing virus recovered directly or indirectly from ANN-1 cells has identified a single protein of
120,000molecularweight (P120)that isencoded by the 5.6-kilobase genomic RNA of A-MuLV (18, 19, 26). Thisprotein contains some of the determinants of theM-MuLV gag geneproducts
(p15, p12,andapart ofp30),and theremaining
90,000 molecularweightis made up of
A-MuLV-specificsequences(25;Baltimoreetal.,inpress).
These A-MuLV-specific sequences share
sero-logical determinants with a normal mouse
cel-lularproteinof150,000 molecularweight
(NCP-150) that is present inspleen, thymus,and bone marrow(25a).
The P120 protein is the only A-MuLV-en-codedproteinthatwehavedetected by usinga
varietyofserologicalreagentsincludingan anti-Abelson tumor (anti-AbT) serum. In addition,
this molecule isexpressedin all of the
A-MuLV-340
on November 10, 2019 by guest
http://jvi.asm.org/
transformedcells wehave examined(26;
Rosen-berg and Witte, unpublished data). Thus, P120 isacandidate for the transforming protein of
A-MuLV. However, mutants expressing altered
P120 are needed to directly assess the role of
this molecule transformation.
Wenowreportthe isolationofthreeA-MuLV
strainsvarying in the size of the A-MuLV-spe-cificprotein. Although all three of these variants transform fibroblasts andlymphoidcellsinvitro,
the variants set a maximum size for the
A-MuLV-specific sequences thatmay be respon-sible for cellular transformation. One of the A-MuLV variants expresses alarger,
160,000-mo-lecular-weight (P160) protein. This A-MuLV strainappearstobeanaturallyoccurring variant that ispresentina secondnon-ANN-1-related
A-MuLV viruspool maintainedatthe National Institutes of Health. The P160 molecule has also been observedby R. Risser and co-workers using these virus stocks (D. Grunwald and R. Risser, personal communication). The other two A-MuLV variants produce smaller proteins, 100,000and 90,000 molecular weight (P100, P90), andwereisolated fromAb-NRK cells (8).
Pro-teins madeby these variants are
nonglycosyla-tedphosphoproteins andexpressthe samegag
genedeterminantsasP120. Thenewisolatesare
stable upon cloning and transform both
fibro-blast andlymphoid cells. The lymphoid
trans-formants aresimilar morphologically and with
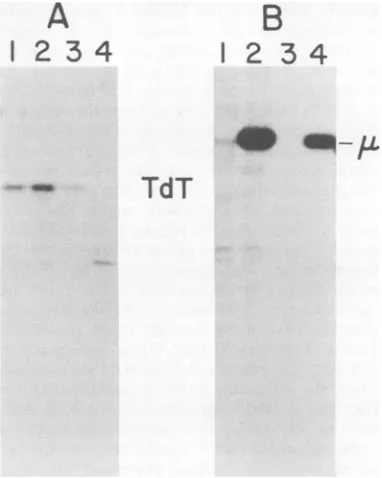
respect to expression of the lymphoid-specific markers immunoglobulin and terminal deoxy-nucleotidyl transferase (TdT).
MATERIALS AND METHODS Cells and viruses. Routine maintenance of cell cultures hasbeen described elsewhere (16, 17). Ab-NRK cells were originallya giftof E. Scolnick,
Na-tional Institutes of Health. ABPC-22 cells (9)were a
gift of M. Potter, National Institutes ofHealth, and
RAW309Crcells(11)were agiftof P.Ralph,Memorial
Sloan-Kettering Institute. Lymphoid cells
trans-formedbythe different A-MuLVisolateswerederived
fromindividual foci ofinfected bonemarrow cells by usingtheagartransformationassay (16)and adapted togrowinliquidcultureaspreviously described (17).
Clones ofA-MuLV-transformed fibroblastswere
de-rivedby platingthe infected cellsinmicrotiterwells
aftera 1.5-hadsorption period. Thetrypsinized cells
were plated at aconcentration of0.3to 0.4cell per well.
All virus stocks were prepared from 24-h culture
fluids and passed through0.45-,um filters beforeuse.
The titers offocus-forming virus in A-MuLV stocks
weredeterminedusingNIH/3T3cells (21).Titers of helpervirusweredeterminedbyassayonS+L- cells
(2)orbythe XCplaqueassay(20).
A-MuLV(P120) (i.e., A-MuLV producinga 120,000-molecular-weightA-MuLV-specific protein)was
pre-pared using ANN-1 cellssuperinfectedwith M-MuLV clone 1 (5). A-MuLV(P160) was prepared using RAW309Cr cells or NIH/3T3 cells transformed by filtered culture fluids from RAW309Cr. The RAW309Cr cells used in theseexperiments release A-MuLVincombination withanNB-tropic, XC plaque-positivehelper virus(datanotshown) that is probably related tothe M-MuLV in theoriginalA-MuLVtumor
extracts.A-MuLV(P100) and A-MuLV(P90)were
de-rived from clones of Ab-NRK cells that had been treated with 10,ug offluorodeoxyuridine per ml for 4 days beforecloning. The A-MuLVwasrescued from thesenonproducerclonesby usingtheamphototropic
virusisolate292 (12) andpassedontoNIH/3T3 cells. Theresultingtransformed cultures ofNIH/3T3cells wereused for routine virusproduction.
Proteinlabeling studies. All radiochemicalswere purchased from NewEnglandNuclearCorp., Boston,
Mass.Techniques forlabeling,immunoprecipitation,
andgelelectrophoresis have been described in detail elsewhere (22, 24, 25).Briefly, rapidly growing cells werelabeled with50,uCiof[3S]methionineper ml in serum-free medium lacking methionine. Cells were labeledincomplete medium with[3H]glucosamineat
100,uCi/ml.Endoglycosidase Hreactionswere as de-scribed (27).Acellextract wasprepared from107 cells by usinglysis buffer (10mMNaPO4, pH 7.5,100mM NaCl, 1% Triton X-100, 0.5% sodium deoxycholate, 0.1%sodiumdodecyl sulfate [SDS])at0°Cand clari-fiedat 100,000xg for2.5 h. Antisera were added to the extract, and the immunoprecipitates were col-lected with Staphylococcus aureus (6) and analyzed onSDS-polyacrylamide gels (7) developed by fluorog-raphy (3).
Antisera used were rabbit anti-M-MuLV reverse transcriptase/p15, rabbit anti-M-MuLV gp7O, goat anti-Rauscher MuLV p12, goat anti-Rauscher plO, goat anti-M-MuLV virion, mouse anti-AbT, rabbit anti-mouse immunoglobulin (a gift of V. Sato), and rabbit anti-calf TdT serum. The characterization of these reagents has been describedelsewhere (22, 24-26; A. Silverstone, L. Sun,0. N. Witte, and D. Balti-more, submitted forpublication). Virus proteins and TdTwereexaminedby electrophoresis of immunopre-cipitatesthrough 10% polyacrylamide gels; immuno-globulin molecules were examined in the presence of 2-mercaptoethanol on 12.5% gels and in the absence of2-mercaptoethanolon 5to 20% gradient gels.
RESULTS
Detectionofa strainof A-MuLV produc-ing a larger form of P120. The A-MuLV-encoded P120 proteinhas been expressed in all of the more than 100 different lymphoid and fibroblast transformed cells we have examined (26; Rosenberg and Witte, unpublished data). These cell lines were all derived in vitro in a similarfashion, either by cloninginfected cells
atlimiting dilution,in the case of thefibroblasts, or by using the agarose transformation assay (16) for lymphoid cells.In the case of lymphoid
cells, allof the celllines expressed asimilarset
on November 10, 2019 by guest
http://jvi.asm.org/
342 ROSENBERG AND WITTE
ofdifferentiation markers and are probably anal-ogous to primitivecells related to B lymphocytes (22, 23). The A-MuLV used in all of these
ex-periments was derived either directly or
indi-rectlyfrom ANN-1 cells.
Toinvestigate the universality of P120
expres-sion, we examined two A-MuLV-transformed
cell linesisolated from tumor tissue by two other laboratories. Two celllinesexpressing different
phenotypic markers were used: the ABPC-22
cell line, a typical immunoglobulin-secreting
plasmacytoma (9), and RAW39OCr, a
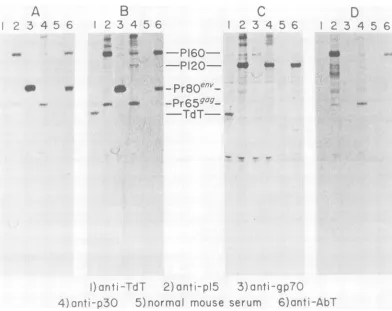
macro-phage line (11). [35S]methionine-labeledcell ex-tractsfromthesecelllines were immunoprecip-itated withantiseraspecific for M-MuLV and
A-MuLVproteinsandanalyzed by electrophoresis
(Fig. 1). The previously described lymphoblas-toid 2M3 cell line (17, 26) produced P120 that reacted with anti-p15, anti-p30, and anti-AbT sera(Fig.1C).However, the ABPC-22 myeloma
B
!
2
3
4
5 6
4:a
cell line (Fig. 1D) and the macrophage cell line
RAW309Cr(Fig.1A)both did notproduce P120, butdidproduce a 160,000-molecular-weight
pro-tein (P160) that was precipitated withanti-p15, anti-p30,and anti-AbT sera. The weak
precipi-tation of P160 with anti-p30 serum was not
observed consistently.Atypical lymphoblastoid
cell line isolated by infecting spleen cellswith virus fromRAW309Cr,the R5cellline(Fig.1B), alsoproduced P160. Thus, P160 expression was notrelated tothe highly differentiated pheno-types expressed by RAW309Cr and ABPC-22
cells.
A more complete survey of the serological
determinantspresent on P160 is shown in Fig. 2.
Immunoprecipitation of P160 from the R5 spleen cell line showed that the protein
con-tained determinants of p15, p12, and p30, but notplO. Anti-AbTserum(lane 7) but not normal mouseserum(lane6) alsoprecipitated P160. In
2 3 4 56
-P160-
-P120-..
2
3
4 5 6,*
aw-Pr80e
;K-am
-Pr65-q-"--
TdT--)1jt TdT 2)anti.-.pIl
5)normal mouse serum 6)ant-AbT
FIG. 1. Analysis of virus-specificproteinsinA-MuLV-transformed celllines. Cell linesRAW309(A),R5
(B),2M3(C),and ABPC22(D)werelabeled with[35S]methionine (100 .Ci/107cells,1h,37QC)and extracted
aspreviouslydescribed (24).A volumeof106cells wasimmunoprecipitatedwith 5,ul ofanti-TdT(lane 1),
anti-M-MuLVp15reversetranscriptase (lane 2), anti-M-MuLVgp7O (lane 3), anti-M-MuLVp 30 (lane 4),
normal mouse serum (lane 5), and mouse anti-Abelson MuLV tumor regressor serum (AbT) (lane 6).
Precipitates were collected with S. aureas, denatured, and analyzed on a 10%o SDS-polyacrylamide gel
developed byfluorography (3, 6, 7).
A
- A >4 6i
I"s.x...R
L,F-3)
ant g pon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.70.462.280.591.2]1
2345
6 7
-P160O-
t
~
Pr8Oenv-_
* *
*
*
Pr659"-
-FIG. 2. Further serologicalanalysis ofP160. The R5celllinewas labeled,extracted, immunoprecipi-tated, and analyzedasdescribed inFig. 1. Lane1,
Anti-M-MuLVp15/reverse transcriptase; lane 2, anti-M-MuLVgp70; lane3,anti-M-MuLVp30;lane 4, anti-Rauscher MuLVplO; lane5, anti-Rauscher MuLVp 12;lane6, normalmouse serum;lane7, anti-AbT.
addition to thegaggene-related determinants, the P160molecule alsocontained A-MuLV-spe-cificdeterminants, becauseprecipitation of P160 by anti-AbT was not blocked by excess
M-MuLV virionprotein (datanotshown). No
reac-tivity with anti-gp70 (Fig. 1 and 2) orwith re-verse transcriptase determinants was detected (data not shown). In addition, P160 was
effi-ciently labeled with inorganicphosphate butnot
with[3H]glucosamine, and the size of the mole-culewasnotalteredbytreatmentwith endogly-cosidase H(datanotshown).
A-MuLV strains synthesizing smaller forms of P120. In another series of experi-ments,weattemptedtoisolateP120mutantsby
selecting for flat variants ofthe Ab-NRK cells
(a P120 producer) (8, 26). Ab-NRK cells were
treatedwith10,ug of fluorodeoxyuridineperml
for4days andclonedatlimiting dilution. After 3weeks,twocloneswerechosen forstudy. One of the clones, AbNRK-1, was morphologically
distinct,beingcomposedofelongated,refractile cells that formedan incomplete cell layer, one
cellinthickness. The secondclone,Ab-NRK-3,
wassimilarinmorphologytotheparentAbNRK cell line, being composed ofcells with a more
regular epithelioid morphology that formed denselypackedlayers.
Todetermine whether themorphologyofthe
two clones was controlled bythe A-MuLV ge-nome,theviruswasrescuedfrom themwiththe
amphotropic wild mouse virusisolate 292 (12)
and passed to NIH/3T3 cells. A-MuLV that transformed NIH/3T3 cells efficiently was
re-covereJ
from both clones. The titers of focus-forming virus, measured in the NIH/3T3 celltransformationassay(21),wereabout thesame:
105focus-forming units for virus recovered from Ab-NRK-3 and 2 x 105 focus-formingunits for
virus recovered from Ab-NRK-l. In addition,
the morphology of the NIH/3T3 cells
trans-formedby both virusstockswas
indistinguisha-ble and typical of A-MuLV-transformed
fibro-blasts.
Focusisolates of cells transformed withvirus
derived from thetwoAb-NRKcell cloneswere
examined forexpressionof the
A-MuLV-specific
protein. All of the clones derived from virus rescued from AbNRK-1 synthesized, instead of P120, a smaller, 100,000-molecular-weight
(P100) protein. The clones derived from virus
rescued from AbNRK-3 fell intotwogroups,one
synthesizing P120 and another synthesizing a
90,000-molecular(P90)protein.Serological
anal-ysis of the P100 andP90molecules is shown in Fig. 3. Extracts from a nonproducer cell line
synthesizingP90wereprecipitated with the
M-MuLVgag gene reagentsanti-p15, anti-p12,and
anti-p30sera (Fig.3A, lanes 2, 3, and 4), butnot
byanti-plOserum(Fig.3A, lane5).P90wasalso
precipitated by anti-AbT serum (Fig. 3A, lane
6). A similar pattern of precipitation was
ob-tained whenextracts from a producer cell line synthesizingP100 wereexamined(Fig. 3B).This
molecule reacted with anti-p15, anti-p30, and
anti-p12sera(Fig.3B, lanes 2, 3, and 4) and
anti-AbT (Fig. 3B, lane 6), but not with anti-plO
serum (Fig. 3B, lane 5). Bothproteins reacted with anti-M-MuLV virion serum (lane 1), and neither reacted with anti-gp7Oserum (lane 7).
The presence of A-MuLV-specific
determi-nants in P90 was demonstrated in a blocking experiment (Fig. 4)whichshowed thatP90was
precipitated with anti-AbT (Fig. 4B, lane 1)
alone or withanti-AbTin thepresence of60 or 180,ug of M-MuLV virion proteins (lanes 2and
3). P90precipitation via gag gene determinants with anti-M-MuLV serum (Fig.4A, lane 1)was
blocked by the presence of60and 180
jig
ofon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.59.231.72.359.2]344 ROSENBERG AND WITTE
MuLV virion protein (Fig. 4A, lanes 2 and 3). P100 molecules also had A-MuLV-specific
de-terminants that couldbedemonstratedin
block-ingexperimentswithanti-AbTserum (data not
shown).
Retrospective examinationofAbNRK-1 cells
and the culture ofNIH/3T3 cellstransformed
by mass infection with AbNRK-1-derived
A-MuLVshowedthat all ofthesecells synthesized
P100. The NIH/3T3 culture transformed by mass infection with Ab-NRK-3-derived A-MuLVsynthesizedboth P120 and P90, but only P120 could be detected in Ab-NRK-3 cells.
Thus, althoughastable changeintheA-MuLV genome in the fluorodeoxyuridine-treated cells
doesnotappear relatedtothealtered
morphol-ogyofsomeof thesecells,thisscreening proce-duredid resultin therecoveryofaltered
P120-like proteins.
Stability of the A-MuLV variants. Virus
stocks fromA-MuLV-transformedcells
synthe-sizingthe various sizes ofproteinswereusedto
transform aseries ofnew clones to determine
whether the size of theA-MuLV-specificprotein
was a stable,heritable trait. Clones of both fi-broblasts and bone marrow cells derivedusing
A-MuLV(P160) and A-MuLV(P100)
synthe-sized theappropriateproteins (Table 1). In ad-dition,anewseries of ANN-1-derived A-MuLV clones were also examined, and all of these clonessynthesized P120 (Table 1).Mostof the clonesderivedusingthe mixed P90 and P120
A-MuLVstock
expressed
onlyP90. Thehighpro-A
A
!~
;_..J
-w
_ < 5 5
..i
40
[image:5.504.318.410.70.251.2]P100 P90 PrB0 Pr65 <
FIG. 3. Analysis ofA-MuLVP90 and P100
pro-teins. Bone marrow cells transformed with
A-MuLV(P90) (A) andA-MuLV(PIOO) (B) were pre-paredandanalyzedasdescribedin Fig. 1.Lane1,
Anti-M-MuLVvirions; lane 2,anti-pl5/reverse
tran-scriptase; lane3, anti-p12; lane 4, anti-p30; lane5, anti-plO;lane6, anti-Abt; lane 7,anti-gp70.
A
3
*
t-
-P90
FIG. 4. P90contains serologicaldeterminants re-active with anti-AbT serum.
A-MuLV(P90)-trans-formedbonemarrowcell line234-9waslabeled with
[35S]methionineandprepared for
immunoprecipita-tionanalysisasdescribed in Fig.1. (A) Extractfrom 106cellsimmunoprecipitatedwith
5Mul
ofapolyvalent goatanti-M-MuLVvirion antiserum in the absence (lane 1)orpresenceof50pg(lane 2)or 180pg(lane 3)ofunlabeledM-MuLVproteins. (B) Equalsamples
immunoprecipitatedwith5ulofmouseanti-AbT
se-rum asdescribed above.Precipitates were analyzed
asdescribed in Fig.1.
portion of the P90 producers may reflect the
ratio of A-MuLV(P90) and A-MuLV(P120) in
the uncloned, transformedNIH/3T3 cellsused
toprepare thevirusstock. Theamountsof P90
andP120inthiscelllinedetectedby metabolic
labeling indicate that P90 is the
predominant
species (datanotshown).
The P160 molecule is the largest
A-MuLV-specific protein we have found; this A-MuLV
strainmaybe the
parental
A-MuLVisolate.Todetermine whetherP120was achanceisolatein
the ANN-1nonproduceror
represented
asecondmajorvariant ofA-MuLV,we
analyzed
the sizeofthe A-MuLV-specific
protein
in a series ofclones derived froma tumorextract similarto
thoseused in theoriginalScherandSiegler (21)
experimentsthatledtotheisolation ofANN-1.
TheA-MuLVinthistumorextract (a
gift
ofR.Siegler) descendedfrom
original
materialgiven
toDr. SieglerbyH. T.Abelson and L. S.
Rab-steinandwasmaintainedby animalpassagein Boston for several years prior to the in vitro
studies. Seven clones of
A-MuLV-transformed
fibroblasts and
lymphoid
cellswereisolatedus-ing this tumorstock, andall seven clones
syn-thesized P120 (Table 1). Therefore, the P120
strain ofA-MuLVwasthemajor componentof the Boston lineage of
A-MuLV,
at leastattheon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.71.262.415.584.2]TABLE 1. Stabilityofthe A-MuLV-specificprotein
uponcloninga
Protein ex- Proteinexpressed in clon-Source of pressedby ally derived transformants A-MuLV parentcell
line P90 P100 P120 P160 ANN-1 P120 0/50 0/50 50/50 0/50 RAW309Cr P160 0/21 0/21 0/21 21/21 Ab-NRK-1 P100 0/12 12/12 0/12 0/12 Ab-NRK-3 P90,P120 12/16 0/16 4/16 0/16 Tumor extract 0/7 0/7 7/7 0/7
aFiltered A-MuLVstockswereusedtoinfect both bone marrowcells andNIH/3T3 fibroblasts.Afteradsorption,the infectedcellswereclonedby usinglimitingdilution forNIH/
3T3cellsand the agarose transformation assay(16)for bone
marrowcells. Transformed cloneswereisolated,and the size of theA-MuLV-specificproteinwasdeterminedbySDS-gel electrophoresis of [nSlmethionine-labeled immunoprecipi-tates.
timein passage atwhich this extractwas
pre-pared.
Lymphoid-specificmarkersexpressed by
A-MuLV-transformedclones. Previous work
has established the presence of
lymphoid-spe-cific gene markers in A-MuLV-transformed lymphoid cells (4, 9,11,22,23). Direct biochem-ical and immunological assays have demon-strated the presence of the markers TdT and intracellularimmunoglobulin inmany cell lines (22, 23). Thefrequencyofexpression of immu-noglobulin and TdTwasmonitored in A-MuLV-transformed bone marrow clonesto determine whether thetransformed cells derived using the different A-MuLV strainswere similar. Clones
derived using A-MuLV(P90) and A-MuLV-(P160)wereanalyzedalong witha newseriesof
A-MuLV(P120)clones. Bonemarrowcellsfrom
C57BL/6J, C57L/J,andBALB/cAn micewere
used for the transformation studies. The
pres-ence of immunoglobulin and TdT was
deter-mined by SDS-polyacrylamide gel analysis of
[3S]methionine-labeled immunoprecipitates (Fig. 5). Consistent with observationsdescribed
elsewhere, the anti-calf TdT serum used here precipitated a 60,000-molecular-weight protein
from murine cell lines synthesizing TdT
(Silver-stone etal.,submitted forpublication). The anti-mouseimmunoglobulinserumusedhere
precip-itated immunoglobulin M (IgM) and IgG from
cell linessynthesizing these proteins(22).
Screening of a large number of lymphoid
clones showed that most of them synthesized TdT(Table 2).About60% oftheclones synthe-sized immunoglobulin in the form of ,u heavy chain (Table 1). No 7 to8S IgM molecules or freelight-chain moleculesweredetected in any ofthe clones analyzed here. The frequency of
immunoglobulin-positive clones and the pres-ence of, heavy chain in the absence of light
chain is consistentwithearlier observations(22). As shown in Table 2, no strain-specific
differ-ences were seenwhen the frequency of
immu-noglobulin and TdT expression was tabulated
accordingtotheA-MuLV isolate used to
trans-form the clones. In addition, when these data
werereanalyzed with respecttothemousestrain
A
12 34
B
12
34
W
~H
[image:6.504.44.237.75.176.2]TdT
FIG. 5. Immunoprecipitationanalysisfor intracel-lularimmunoglobulinand TdT in clonal
A-MuLV-transformedlymphoidcell lines. Individual clonesof
A-MuLV-transformedcells(lanes1to4)werelabeled
andpreparedasdescribed inFig. 1, then
immuno-precipitatedwith5,ulofrabbit anti-TdT(A) anda polyvalent rabbit anti-mouse immunoglobulin (B).
Sampleswereprocessedasdescribed inFig. 1. TdT
expression was analyzed on a 10%
SDS-polyacryl-amidegel,andimmunoglobulinexpression was an-alyzedona12.5%SDS-polyacrylamidegel.
TABLE 2. Frequency of A-MuLV-transformedbone
marrowclonesexpressinglymphoid markersa
Frequency in A-MuLV strain: Marker
P160 P120 P90
IL 15/22 6/11 2/3
TdT 20/22 9/11 2/3
aSingle-focus isolates of A-MuLV bone marrow
cells were adapted to grow in liquid medium and examined forexpression of immunoglobulinandTdT by SDS-gel electrophoretic analysis of
[nS]methio-nine-labeled immunoprecipitates(22, 23). The size of the A-MuLV-specificprotein was confirmed in each of the clones by electrophoretic analysis of labeled immunoprecipitates.on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.247.438.153.392.2] [image:6.504.248.438.531.583.2]usedasa sourceofbone marrowcells,nomouse
strain-specific differences in the frequency of
marker expression were evident (data not
shown).
DISCUSSION
Allof the A-MuLV-transformed cell lineswe
have examined synthesizeanA-MuLV-specific protein that contains M-MuLV gag
gene-de-rivedsequences.Allofthemoleculesare
nongly-cosylated phosphoproteins, and the size ofthe
different proteins is stable upon cloning. Al-though these proteinsvary in sizefrom 160,000 to 90,000molecular weight, they aresimilar in
theirstructure,beingmadeupofthe
M-MuLV-derivedp15, p12, andpartof thep30 protein in
additionto anA-MuLV-specific region. Because
theproteinscontain thesamegaggene-derived
determinants, theyprobablyvary inthe size of theirspecific region. If the
A-MuLV-specific regionof thesepolyproteinscontains the
transforming sequences, the A-MuLV(P90)
strainplacesamaximum limitonthe size ofthis
region. Because P90 resembles P120in structure,
about 30,000molecularweight of P90 is probably derived from the M-MuLV gag geneproducts
p15,p12, and p30 (26; Baltimoreetal.,inpress). Thus, theremaining 60,000 molecularweightof
P90, the A-MuLV-specific sequences, is
suffi-cient toencode for anytransformingfunctions thesesequencesdirect.Inaddition,the similar-ity of lymphoid markers expressed by clones transformed with all of these isolates shows that the addition ofsequences inA-MuLV(P160)and the loss ofsequencesinA-MuLV(P90) doesnot
affecttargetcell selection.
The P160 strain ofA-MuLV appears tobea
naturallyoccurring isolate thatprobablyis the
predominant component of A-MuLV stocks maintainedattheNational Institutes ofHealth bypassageoftumorextracts. R. Risser and D.
Grunwald, usingA-MuLV derived from the
Na-tional Institutes of Health pool ofvirus, have
identified a 160,000-molecular-weight protein
thatexpressesp12determinantsinsomeof their A-MuLV-induced tumors (R. Risser, personal
communication). This protein is probably the
same asthe P160 describedhere.
The P160 protein probably contains about
130,000 molecular weight of A-MuLV-specific
sequences;basedontheabilityofA-MuLV(P90)
totransformcells,overhalfofthesesequences
must notberequiredtotransformcellsin vitro.
Whetherthese sequences are beneficial to the
virusgrowthandoncogenicityinvivo isunclear.
However, the predominance ofA-MuLV(P120)
in another tumor extract virus pool indicates thatA-MuLV(P160) is notalways selected byin
vivo passage.Whether differences in the passage history of the Boston and National Institutes of
Health pools of A-MuLV are important in the
derivation of two A-MuLV strains cannot be ascertained at thistime.
The genomes of the new A-MuLV strains have notbeenfullycharacterized. However, the
3.6-kilobase central region of the A-MuLV(P120) genome is large enough to encode 130,000
mo-lecular weight of protein. Thus, if the
A-MuLV(P160)genome is the same size as the
A-MuLV(P120) genome, P160 would account for
the entire codingcapacity of the genome.
The origin of theshorter A-MuLVproteins is
unclear. Thepossible role of fluorodeoxyuridine
treatment in their origin must be considered. although neither P100 or P90 appears to be directly relatedtothemorphological variations observedin thefluorodeoxyuridine-treated cells,
20 random Ab-NRK clones did not synthesize smaller forms of P120 (0. Witte, unpublished
data). The Ab-NRK cells were originally iso-lated using virus from ANN-1 cells. We have
shown that this strain of A-MuLV is stable through repeatedcloningin mousecells(26) (see above).
Thelymphoidmarker studies indicate that all
thestrains of A-MuLV transform similartypes
of cells. In the case of A-MuLV(P160), which
was originally detected in cell lines ofpeculiar
differentiationtypeforA-MuLV,it isclear that
the P160 protein is not responsible for these
alteredphenotypes. Our dataareconsistent with thoseof Raschkeetal. (11),whoshowed that
A-MuLV produced byRAW309Cr, amacrophage
cellline, produced typical Abelsonlymphomas.
Thepercentageof A-MuLV clonesexpressing
,uchain is similartothefrequency reported by
Sidenetal.(22)usingthesamemethodtoisolate
thetransformed clones. The
IL-only
phenotypeis the most common immunoglobulin phenotypeexpressed by A-MuLV-transformed lymphoid
cellsisolated in vitrousingfocal transformation
assays.Therelationships,ifany,of these,u-only cellstonormallymphocytesis unclear. No
nor-mal ,u-only cell has been detected, although
suggestiveevidence of,u-onlycells exists (10;E.
Siden and D. Baltimore, personal
communica-tion). Sucha cellmightbe theprecursor of the
cytoplasmic IgM-positive pre-B cells. The
abil-itytostimulatecomplete IgMsynthesisinsome
A-only
A-MuLV-transformed lymphoid cell lines byusing the B lymphocytemitogenlipo-polysaccharide (19) suggests that such a
path-way may exist.
TdT expression by A-MuLV-transformed
cellshasbeenpreviouslyreported(4,23),butno
prospective surveys have beenconducted. This
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
study shows that almost all
A-MuLV-trans-formed cells express TdT. Many of the
TdT-positive clones also synthesized,i chain. These results support thehypothesis suggested by Sil-vertone et al. (23), that TdT expression is not
limited to committed pre-T-lymphocytes, but may characterize alltypesofimmature
lymph-oidcells.
The lymphoid marker survey also indicates that thecellssusceptibletoA-MuLV in the bone marrowofBALB/cAn, C57BL/6J, andC57L/J
mice areprobably similar. Thisresult is
signifi-cant because these three strains differ in their
susceptibility to A-MuLV-induced tumors, BALB/c being susceptible and C57BL/6J and
C57Lbeingresistant (15).Whereasprevious
re-sultshave shown that bone marrow cells from these three strainsare susceptible to A-MuLV in vitro (16), the results describedhere demon-strate that similar A-MuLV-sensitive target
cells are present in all three strains. Thus, the resistance ofC57BL/6J and C57Ladult miceto
A-MuLV is probably not mediated by altera-tions in target cellpopulation.
ACKNOWLEDGMENTS
We aregratefultoDanClarkforexcellent technical assist-ance and to David Baltimore for his support and helpful discussions.
Thisworkwassupported by Public Health Service CA-24220andProgramProjectgrantCA-24530from theNational CancerInstitute (N.R.),bygrantVC-4Jfrom the American CancerSociety (to DavidBaltimore),andby Public Health Service grantCA-14051 from the National Cancer Institute (coregrant toS. E.Luria).N.R.is arecipientofanAmerican CancerSocietyResearch Scholar Award(Massachusetts Di-vision). O.N.W.is aHelenHayWhitney Postdoctoral Fellow.
LITERATURE CITED
1. Abelson, H.T.,andL.S. Rabstein.1970.Influence of prednisoloneonMoloney leukemogenicvirus inBALB/ cmice.CancerRes. 30:2208-2212.
2. Bassin, R. H., N.Tuttle, and P. J.Fischinger. 1971. Rapid cell cultureassay technique for murine leukemia viruses. Nature (London) 269:564-566.
3. Bonner,W.M., andR. A.Laskey.1974. A film detection methodfortritium-labeled proteinsandnucleicacids in polyacrylamide gels. Eur.J.Biochem. 46:83-88. 4. Boss, M., M. Greaves, and N. Teich. 1979. Abelson
virus-transformed haematopoieticcell lines with pre-B cell characteristics. Nature (London) 278:551-553. 5. Fan,H., and M. Paskind. 1974. Measurement of the
sequence complexity of clonedMoloneymurine leuke-mia virus 60 to 70 S RNA: evidence for a haploid genome. J. Virol.14:421-429.
6. Kessler, S.W. 1975.Rapidisolation of antigens from cells with a staphylococcal protein-A-antibody adsorbent: parameters of the interaction ofantibody-antigen com-plexeswith protein A. J. Immunol.115:1617-1624. 7. Laemmli, U.K. 1970. Cleavage of structural proteins
duringthe assembly of the head ofbacteriophageT4. Nature(London) 227:680-685.
8. Parks,W.P., R. S.Hawk,A.Anisowicz, andE. M. Scolnick. 1976. Deletion mapping ofMoloneytype C
virus:polypeptideand nucleic acid expressionin differ-enttransforming virus isolates. J.Virol. 18:491-503. 9. Potter,M.,M. D.Sklar,and W. P. Rowe.1973.Rapid
viral induction of plasmacytomas in pristane-primed BALB/c mice. Science 182:592-594.
10. Raff,M. C., M. Megson,J. J. T. Owen, and M. D. Cooper.1976.Early productionof intracellularIgMby B-lymphocyte precursors in mouse. Nature (London) 259:224-226.
11. Raschke,W.C., S.Baird,P.Ralph, andI.Nakoinz. 1978.Functionalmacrophage cell lines transformedby Abelsonleukemia virus. Cell 15:261-267.
12.Rasheed, S.,M.B.Gardner, andE.Chan.1976. Am-photropichost rangeof naturally occurringwild mouse leukemiaviruses.J. Virol.19:13-18.
13. Reynolds,F.H., Jr.,T.L.Sacks,D. N.Deobaghar, andJ. R. Stephenson. 1978. Cells nonproductively transformed byAbelson murine leukemia virus express ahighmolecularweight polyprotein containing struc-turalandnon-structuralcomponents. Proc. Natl.Acad. Sci. U.S.A.75:3974-3978.
14. Reynolds, R. K., W. J. M. van deVen, and J. R. Stephenson. 1978.Translation oftypeC viral RNAs inXenopuslaevis oocytes: evidencethat the 120,000-molecular-weight polyproteins expressed in Abelson leukemiavirustransformed cellsis virus coded. J. Virol. 28:665-669.
15. Risser,R.,M.Potter, andW. P.Rowe.1978.Abelson virus-inducedlymphomagenesis in mice. J.Exp.Med. 148:714-726.
16. Rosenberg, N., andD.Baltimore.1976. Aquantitative assay fortransformationof bone marrow cellsby Abel-sonmurine leukemiavirus. J.Exp. Med.143:1453-1463. 17. Rosenberg,N., andD.Baltimore. 1978.Theeffect of helpervirus onAbelson virus-induced transformation oflymphoid cells.J.Exp.Med. 147:1126-1141. 18. Rosenberg, N.,D.Baltimore, and C.D.Scher. 1975.
In vitrotransformation oflymphoid cells by Abelson murine leukemia virus. Proc. Natl. Acad. Sci. U.S.A. 72:1932-1936.
19. Rosenberg, N., E. Siden, and D. Baltimore. 1979. Synthesis of,u chains by abelson virus-transformed cells and inductionoflight chainsynthesiswith lipopolysac-charide,p. 379-386. In M.Cooper,D.Mosier,J.Scher, and E. Vitetta (ed.), Lymphocytes and the immune response. Elsevier, North Holland Biomedical Press, New York.
20. Rowe,W.P.,W. E.Pugh,andJ. W.Hartley. 1970. Plaqueassaytechniquesfor murineleukemiaviruses. Virology42:1136-1144.
21. Scher,C.D.,and R.Siegler.1975.Direct transformation
of 3T3cellsby Abelson murineleukemia virus. Nature (London)253:729-731.
22. Siden,E.J.,D.Baltimore,D.Clark,and N.
Rosen-berg. 1979. Immunoglobulin synthesis by lymphoid cells transformedinvitroby Abelsonmurineleukemia virus.Cell16:389-396.
23. Silvertone, A.E., N. Rosenberg, V.L. Sato,M. P.
Scheid,E.A.Boyse, andD.Baltimore. 1978. Cor-relating terminaldeoxynucleotidyltransferase andcell surface markersin thepathway of lymphocyte ontog-eny, p. 433-453. In B. Clarkson,J. E. Till,and P. A. Marks(ed.), Differentiation of normal and neoplastic hematopoietic cells. Cold Spring Harbor Laboratory, ColdSpring Harbor,New York.
24. Witte,0.N.,and D. Baltimore. 1978.Relationship of retrovirus polyprotein cleavagestovirion maturation studied withtemperature-sensitivemurine leukemia
vi-rus mutants.J. Virol. 26:750-761.
25. Witte,0.N.,N.Rosenberg, andD.Baltimore. 1979. Preparationofsyngeneictumorregressor serum reac-tive with the uniquedeterminants of the murine
on November 10, 2019 by guest
http://jvi.asm.org/
348 ROSENBERG AND WITTE
kemiavirus-encoded P120proteinatthe cell surface. J. Virol. 31:776-784.
25a.Witte,D.N.,N.Rosenberg,and D.Baltimore. 1979. Identification ofanormal cellular protein cross-reactive
tothe major Abelson murine leukemia virusgene
prod-uct.Nature(London)281:396-398.
26. Witte,0.N.,N.Rosenberg,M.Paskind, A. Shields,
andD.Baltimore. 1978.IdentificationofanAbelson murine leukemia virus-encoded protein present in transformed fibroblast and lymphoid cells. Proc. Natl. Acad. Sci. U.S.A. 75:2488-2492.
27. Witte, 0. N., and D. Wirth. 1979. Structure of the
murine leukemia virusenvelope glycoproteinprecursor.
J.Virol.29:735-743.
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG. 4.formedgoat[35S]methioninerumtionofactive(laneimmunoprecipitatedas106 unlabeled described P90 contains serological determinants re- with anti-AbT serum](https://thumb-us.123doks.com/thumbv2/123dok_us/1501736.102900/5.504.71.262.415.584/formedgoat-methioninerumtionofactive-laneimmunoprecipitatedas-unlabeled-described-contains-serological-determinants.webp)