Vol. 28, No. 2 JOURNALOFVIROLOGY, Nov.1978, p.499-517
0022-538X/78/0028-0499$02.00/0
Copyright©1978 American Society forMicrobiology Printed in U.S.A.
Recombinants Between
Herpes
Simplex
Virus Types 1 and 2:
Analyses of Genome Structures and Expression of Immediate
Early Polypeptides
V. G. PRESTON,* A. J. DAVISON, H. S. MARSDEN, M. C. TIMBURY, J. H. SUBAK-SHARPE, AND N. M. WILKIE
Medical Research Council Virology Unit, University of Glasgow, Glasgow Gll5JR, Scotland Received for publication 17 April 1978
Recombinants between temperature-sensitivemutantsofherpessimplexvirus
types 1(HSV-1) and2(HSV-2)wereconstructed.Using restrictionendonucleases,
weanalyzed thegenomecomposition of17intertypicrecombinants and detected
crossovers in everyregion of the genome. Thevirion DNA ofonerecombinant
appearedtobelargely "frozen" intwoof the four possiblegenomearrangements of HSV. Knowledge of the genome structures ofrecombinants enabled us to
physically map immediate early polypeptides. We present evidence that the
immediateearly polypeptideVmw IE 110of HSV-1and itsfunctionally equivalent polypeptide,Vmw IE118,of HSV-2may mapinthe repetitive sequences bounding
thelong unique region of HSV.
The genomes of bothherpes simplex viruses
type 1(HSV-1)andtype 2(HSV-2) containtwo
unique (U)sequences,ofapproximately9 x 106
daltons (theUssequence) and 68 x 106daltons
(the UL sequence), which are bounded by
ter-minal and internal invertedrepetitions (TR and IR, respectively) ofapproximately 4 x 106
dal-tons (TRs and IRs) and 6 x 106 daltons (TRL
andIRL), respectively (4, 25, 31).Themannerin
which these sequencesarearranged onthe ge-nomeisillustratedatthe bottom ofFig.3.Asa
consequence of this unusual structure, the
unique regions can invert with respect to one another, and fourmajor genome arrangements
have been identified in virion DNA(4, 6, 11,25, 26).
HSV-1 and HSV-2 share homologyin about 50% of their DNA sequences (15), and both complementation and recombination between
the two serotypes have been reported (7, 29).
Studies on virus-induced polypeptides and an-tigens confirm that true recombinants result fromgeneticcrossesbetween HSV-1 and
HSV-2 (10). The restriction endonuclease maps of HSV-1 and HSV-2 DNAs (4,5a)arevery
differ-ent, and this property allows the location of HSV-1 and HSV-2 DNA sequences in the
ge-nomesofintertypic recombinants (19, 33).
Analysis by sodium dodecyl
sulfate-polyacryl-amidegelelectrophoresis(SDS-PAGE)resolves
about50virus-inducedpolypeptides incells
in-fectedwithHSV-1 orHSV-2 (12, 17,20). Most
HSV-1-induced polypeptides can be
distin-guished from HSV-2-induced polypeptides
solely on thebasis of differences inmobilityin SDS-PAGE (21). By analysis of polypeptides inducedby recombinants of knowngenome
com-positions, it ispossibletodetermine thephysical locations of the genesfor several polypeptides.
Thestrategyisessentiallythesame asthatused
to determine thephysical map coordinates for adenovirustemperature-sensitive (ts) mutations andpolypeptides (9, 18, 24,34).
Inthiscommunication,thegenome structures
of17recombinants between HSV-1 and HSV-2, including1whichappears tobelargely"frozen" intwooutof thefourpossiblegenome
arrange-mets, are analyzed. Knowledge of the genome
structureshasenabledus todetermine themap
coordinates for polypeptides belonging to the immediateearly (IE) class (22) of HSV-induced polypeptides.SincesomeIE RNAis encodedby
therepeatregionsof HSV-1 (5),wehave
exam-ined IE polypeptides induced by recombinants containingoneHSV-1 L repeatandoneHSV-2
L repeat to elucidate the expression of genes
within theserepetitivesequences.
MATERUILS AND METHODS
Cells. BHK-21 clone 13(C13) cells(16)wereused
in allexperiments except in the isolation of certain
HSV-1/HSV-2 recombinants, in which Vero cellswere
used. BHK-21 (C13) cells were cultivated in Eagle
medium containing twice the concentration of
vita-mins and amino acids, 10% (vol/vol) tryptose phos-phatebroth, and10% (vol/vol)calfserum.Verocells
weregrownin this mediumexceptthat 10% (vol/vol)
fetal calfserum (Gibco-Biocult) wasused instead of calfserum.
499
on November 10, 2019 by guest
http://jvi.asm.org/
500 PRESTON ET AL.
Virus. Two HSV-1ts mutantsfrom strain17
(Glas-gow),tsB syn+(nonsyncytial plaque morphology) and
tsD syn (syncytial plaque morphology) (3),andone
HSV-2tsmutant,ts1,from strainHg52(28),together
with the parental wild-type viruses,wereused. Virus stocks. All virus stocksweregrownin
BHK-21 (C13) cellsasdescribedpreviously (3).
Virus titration. Virus titrationwasperformedas
describedbyMarsden etal. (17). Infected cellswere
incubatedat31°C, the permissive temperature, for3
daysor at38.5°C, the nonpermissive temperature, for
2days.
Isolation of recombinants. Recombinants 2Sa,
15 Sa, 34 Nsd, and 34 sd were isolated in BHK-21 (C13) cells from thegeneticcross(ts D synxts1)as
described byTimbury andSubak-Sharpe(29). In this and subsequent papers these recombinants will be referred to as Dx1(2), DX1(15), Dx1(34-1), and Dx1(34-2), respectively.Similarly, recombinants 2444, 2641, 2853, 3134, 3145, and R13, isolated from the
geneticcross (tsB syn+ x tsl),willnowbeknownas
Bx1(24), Bx1(26), Bx1(28), Bx1(31-1), Bx1(31-2), and Bx1(13),respectively. Note that inWilkieetal. (33) theearliernomenclature has been used.
Recombinants Bx1(24), Bx1(26), Bx1(28),
Bx1(31-1),andBx1(31-2)wereisolated in Vero cells
as described in Fig. 1. Recombinants Dx1(31),
Dx1(32), Dx1(43), Dx1(48), Dx1(51), Dxl(53),
Dx1(57), and Bx1(13)wereisolated in BHK-21(C13) cellsessentiallyasdescribedforBx1(24).
SDS-PAGE.Sampleswereanalyzed by
discontin-uous SDS-PAGE. Two types of gels were used: a
gradient gel (5.5% to 12.5% polyacrylamide with a
stacking gel of 5% polyacrylamide) as described by
Marsden et al. (17) and a single-concentration gel
(7.5%polyacrylamidewithastacking gelof4.5%).The
ratio ofN,N'-methylenebisacrylamide to acrylamide
in thelatter type ofgelwas1:40 (wt/wt). The same
buffer (0.05 M Tris,0.055 Mglycine, 0.1%SDS) was
used in both the upper and the lower buffer tanks. Afterelectrophoresis, thegelswerefixed and stained
asdescribedpreviously(17).
Autoradiography. The gelswere dried, and
au-toradiographsweremade with Kodak KodirexX-ray filmat roomtemperature.
Virus-induced IEpolypeptides. A temperature
of38.5°Cwasusedthroughoutvirus induction of IE
polypeptides. Cells in30-mmpetri disheswere prein-cubatedfor 1.5h before virus infection inEagle
me-diumcontaining200pgofcycloheximideperml. Cells
were infected at amultiplicityof infection of about 20
PFU percell, washed, and incubated forafurther5h, all in the same medium. Actinomycin D was then addedto afinal concentration of2.5jug/ml.After15
min, this cycloheximide-containing medium was re-moved, and plates were washed twice with
methio-nine-reducedEagle medium containing 2% calfserum and actinomycin D (2.5pg/ml). [35S]methioninewas thenaddedat aconcentrationof 100lCi/ml,andthe cellswereincubated forafurther2h.Control cultures
were treated in thesame mannerexceptthat cyclo-heximidewasomitted.
Samples were harvested as follows. The medium
was removed from the petri dishes, and denaturing
buffer (final sample buffer [17]) (0.35 ml) wasadded
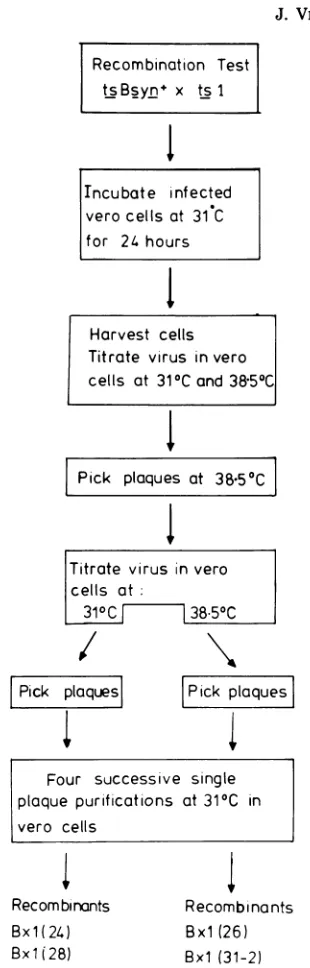
Recombinotion Test tsBsyn x ts I
I~
Tncubate infected vero cells at 31 C for 24 hours
1
Harvest cells Titrate virus in vero cells at 310Cand 38.500
1
Pick plaques at
38.50C
Titrate virus in vero cells at:
3100 38.500
Pick plaquesl
1
Pick
plaques
Four successive single plaque purifications at 310C in
vero cells
Recom binants Bx1(24)
Bxli28) Bxl(31-1)
Recombinants Bxl (26)
Bx1 (31-2)
FIG. 1. Isolation ofHSV-1/HSV-2 recombinants from thegeneticcross(ts Bsyn+xts1).
to each dish. Thedishes wereheated to 70°C for 15
min.Thecontents werethentransferred to vials and furtherheatedfor2 min at 100°Cto reduce the vis-cosity of the samples. The amount of radioactivity incorporatedintotrichloroaceticacid-precipitable ma-terialwasdeterminedsothatsamples with the same
amountofradioactivity couldbeloadedontogels.
Labeling and isolation of viralDNA. Samples
of 107 BHK cells in 10 ml ofphosphate-free Eagle J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.499.286.441.55.539.2]VOL. 28, 1978
mediumsupplemented with 2% calfserum wereseeded
onto 90-mm petri dishes and left for 24 h at 370C. Thesecells werethen infectedat31°Catamultiplicity
ofinfection of about 5. After 1 hof absorption they
werewashed andincubatedat 31°C,all inthesame
medium. Two hours postabsorption, carrier-free 32p;
(Amersham)wasaddedtoafinal concentration of100
,uCi/ml, and thecells wereincubated forafurther 24
hat31°C. The culture mediumwasremoved,and
cell-released viruswaspelletedat25,000rpminaBeckman
Ti 50rotorfor1h. The DNAwasextractedfrom the
virionsaspreviouslydescribed by Wilkie (30).
Alter-natively,virions wereextracted frominfected-cell
cy-toplasm as described by Wilkie (30) and combined
with tissue culture medium containing cell-released virus. Viruswaspelleted and viral DNAwasextracted
in the same way asdescribed for cell-released virus.
Phagelambda DNA. Lambda DNAwasobtained
by phenol-SDS extraction ofCsCl-banded virus from
a heat-inducible lysogenic host strain, Escherichia
coli 805(CI 85757).
Restrictionendonuclease analysis. Restriction endonucleasesEcoRI, HindIII,Bgl II, Xba I, Hpa I, andKpnIwerepurifiedessentiallyasdescribed
pre-viously (4, 33; R. N.Yoshimori, Ph.D. thesis, Univer-sity ofCalifornia, San Francisco, 1971). One unit of
enzymeis definedasthatquantity ofenzyme neces-sary to produce a limit digest with 1 ,ug of lambda
DNA after incubation for 3 hat370C inavolume of
50
pl
of0.01 MTris-hydrochloride (pH 7.4),0.006 M B-mercaptoethanol,0.006MMgCl2,and0.1%(wt/vol) bovineserumalbumin.Labeled HSV DNA samples containing less than
0.01 ,ugof viral DNAwere digested under the same
reaction conditions with1,ugoflambda DNA and1to 2 U of enzyme. Reactions were terminated by the
addition of 0.1 volume of 25% Ficoll-0.1 M EDTA containing bromophenol blue. DNAfragments were
separated by electrophoresis onslab gels of 0.3, 0.5,
0.75,or1.0%agaroseaspreviouslydescribed(32).The
gelsweredried, and autoradiographsweremade with
Kodirex KD54Tfilm atroomtemperature.
DNA-DNA hybridization.Unlabeled HSV DNA
was prepared essentially as described before (30).
HSV-1 and HSV-2 DNAs, further purified by rate
zonal centrifugation (32), were nick translated with
32P-labeled deoxyribonucleosidetriphosphates (Radi-ochemical Centre) by the technique described by Rigbyetal.(23). UnlabeledHSV-1 and HSV-2 DNAs
were also cleaved with Hpa I and Xba I, and the
fragmentswere purified byhydroxylapatite
chroma-tography as describedpreviously (31).Isolated
frag-mentswerealsonick translated. Thenick-translated
DNAshad specific radioactivities intherangeof4x
107to2x 108cpm/Ag.
Unlabeled DNA from Bx1(28) was cleaved with
Kpn I plusXbaI,and thefragmentswereseparated
by electrophoresis on slabgels of0.5%agarose (31).
DNAfragmentsweredenatured with alkali in situ and
transferredtosheets of nitrocellulosebythetechnique ofSouthern(27). After alkalidenaturation,
nick-trans-lated[32P]DNAwashybridizedto
DNA-fragment-con-tainingstripsfromthe nitrocellulose membrane sheets
asreportedpreviously (31). Afterwashingthe
nitro-cellulose membrane strips in 2x SSC (0.30M NaCl
HSV INTERTYPIC RECOMBINANTS 501 plus 0.03 Msodium citrate) at 55°C, DNA fragments hybridized to the32P-labeledprobewerevisualizedby
autoradiography.
RESULTS
Biological properties of recombinants. Most of therecombinants grew as well as
wild-type HSV-1 andHSV-2 viruses, but therewere some which grew poorly atthe nonpermissive temperature,withvariable and lower efficiencies of plating at 38.5°C/31°C. Table 1 shows effi-ciencies of plating at 38.5°C/31°C for the re-combinants and the controls. Recombinants Bxl(31-1) and Bxl(31-2), which are clonally related,weretsinBHK-21 (C13) cells andwere
never observed to form plaques at 38.5°C. In Vero cells, however, in which they were origi-nallyisolated, they generally formed plaques at 38.5°C and had muchlower and more variable efficiencies of plating at 38.5°/31°C than did wild-type HSV-1 and HSV-2.
Analysis of crossoverpointsin
recombi-nants. To analyze the crossover points in the
DNAof therecombinants, the physical maps for HSV-1 and HSV-2 DNA have to be aligned. Results from the hybridization of separated HSV-1 [32P]DNAfragmentstounlabeled sepa-rated HSV-2 fragments and vice versa by the
TABLE 1. Relativeefficienciesofplating(EOPs) of recombinant viruses(38.50C/31°C)
Relative EOP Temperature Virus (38.50C/310C) sensitivity'
Dxl(2)b 1.5X 10-2 Reduced
DX1(15)b 7.3x 10-3 Reduced
DX1(34_1)b 4.4 Wildtype
DX1(34-2)b 6x 10-' Wild type
Bxl(13) 1.2 Wildtype
Bxl(24) 1.3x 10-' Reduced
Bx1(26) 2.3x10-2 Reduced
Bxl(28) 9.5x 10-' Wildtype
BXl(31-1) <1.9Xlo--5 ts
BXl(31-2) <3.4X 10-6 ts
DX1(31) 1.9 Wildtype
Dxl(32) 1.5 Wildtype
Dxl(43) 1.4 Wildtype
DX1(48) 1.6 Wild type
DX1(51) 6.9x 10-1 Wildtype
Dxl(53) 2.1 Wild type
Dx1(57) 4.4 Wildtype
Hg52 5.8x 10-1 Wildtype
Strain17(syn+) 7.3x10-1 Wildtype
Strain17(syn) 1.1 Wildtype
ts 1 <1X10-6 ts
ts D (syn) <2x 10-6 ts
tsB (syn+) <5x 10-6 ts
aWildtype, EOP>2 x
10-1;
ts, EOP< 1 x10-4;reduced,EOP<2x 10-1and>1 x10-4.
bSee reference29.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.499.252.443.395.644.2]502 PRESTON ET AL.
Southern blottechnique (27) (N. M. Wilkie and R.Cortini,unpublishedobservations)showthat HSV-1fragmentshybridizetoHSV-2fragments with corresponding map positions on the
ge-nome,indicating thatsequencehomologyis
dis-tributed according tomap location throughout both the L and the S regions of the genome.
Thisinforination allowsalignment of the
HSV-1 and HSV-2 physicalmaps asshown in Fig.3
for sixrestriction endonucleases. Itcanbeseen from Fig. 3 that the restriction endonuclease
mapsof HSV-1 and HSV-2 differradicallyfrom each other.
The analyses of the DNAs from two
recom-binants, Dx1(53) andBx1(28),aregivenin de-tail toillustrate how the locations ofcrossover sites were determined. Figure 2 shows the re-striction endonucleasepatternsof DNAs fromts D,ts1,andrecombinant Dx1(53) isolated from (ts D x ts 1). Analysis depended on observing which DNA fragments from the recombinant DNA comigratedwith fragments ofone orthe other of theparentaltypes.Whenarecombinant fragment comigratedwith aparentalfragment, itwasassumed that the restriction sites which delimit the parental fragment were present in this recombinant DNA. In some cases novel fragments, which corresponded toneither
par-ent, were observed. Such fragments consisted
partly of HSV-1 and partly of HSV-2 DNA
sequences and hence contained the crossover
sites.Thus,acombinationmapofparental-type restriction endonuclease sites could be
con-structedfor the recombinant genomewhich al-lowedanestimate of thecrossoverpoints in the recombinant DNA.
Analysis ofDx1(53)DNA.HpaIfragments ofDx1(53) DNAcomigratedwith allthe
HSV-2fragments excepte. Fragments corresponding
to HSV-1 eandqwere alsopresent.The band inthe recombinantprofile correspondingtothe
1 M band q of HSV-1 was, however, 2 M in relativeamount,suggestingthat therewere two
fragmentsof this size in theHpaIdigestof the recombinant genome. The additional fragment
the size ofq can be accounted forbya crossover
eventwithin HSV-1 sandHSV-2 e.Therewas
also another novel fragment, which had a
slightlygreatermolecularweightthan HSV-1m.
Thisnovelfragmentcanbeexplained bya
cross-over eventwithin HSV-1fand HSV-2 e (refer
tophysicalmaps[Fig. 3]).
The region of the recombinant genome in
whichtheparentalnatureoftheDNA was
un-certain isrepresentedinFig.3bycrosshatched
lines. The sites which delimit this region give
the map coordinates between whicha crossover
has occurred. Thus, from the Hpa I analysis,
DX1(53) consisted ofHSV-2 DNAwith an
in-sertion of HSV-1 DNA with maximum map
coordinates of0.56 to 0.75 and minimum map
coordinates of0.60 to0.72.
The results of all the other enzyme digests
wereinagreementwith thisanalysis and further
defined the estimates of the map coordinates for
the crossovers. By combining all theresults, it
could be concluded that Dx1(53) contained an
insertion of HSV-1 DNA with maximum map
coordinates from 0.58to 0.74and minimummap
coordinates from 0.60 to 0.72.As Dx1(53) was
wildtypeingrowth properties,weassumed that
thetslesionts1hadbeenreplaced by wild-type information from HSV-1 DNA, and hence that
ts 1lay within themapcoordinates0.58 to 0.74.
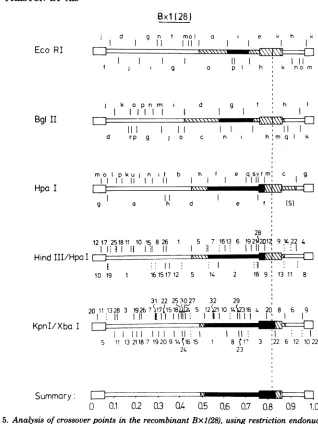
Analysisof the DNA of Bx1(28).The DNA restriction endonuclease fragment patterns of Bx1(28)weremuchmorecomplex than those of Dx1(53). Figure 4 shows autoradiograms of Bx1(28) [32P]DNAdigested with EcoRI, BglII,
HpaI,HindIIIplusHpa I, and KpnIplus Xba I.Individual fragmentswereidentified bya sim-ilar analysistothatgiven for Dx1(53). It could be deduced that Bx1(28) contained HSV-2
se-quencesfrommapcoordinates0tobetween0.49
and0.52,where therewas acrossover toHSV-1
sequences;theHSV-1sequencescontinuedto a
point in IRs (or
TRs)
andthen crossed backtoHSV-2 sequences. Most of the S region
con-tained HSV-2 DNA. Inaddition, the KpnI/Xba Idouble-digestdata strongly suggested that the population of DNAwasheterogeneous inthe S
region, and this will be considered inmoredetail
below. Figure 5 summarizes the restriction
en-donuclease analysis of the crossover points in
Bx1(28).
Previous studies (4, 11, 26) have shown that,
as a consequence of inversions of UL and Us,
restriction enzymes which cleave HSV DNA
only in Us and UL produce four terninal
frag-ments, with relative molarities of0.5, and four
fragments which span the "joint" region, with
relative molarities of0.25. Figure 4 showsthat
the terminalfragmentHSV-2EcoRI-f appeared
tobe overrepresentedin the DNA ofBx1(28). Conversely, thejoint-spanning fragments
HSV-2 EcoRI-b and -d, which contained EcoRI-f, appeared tobe underrepresentedin the digest.
In the Bgl II digest, joint-spanning fragments
HSV-2aand b werealsounderrepresented,and
these also contained the HSV-2 EcoRI-f
se-quences. The HSV-1Bgl II-ffragment,from the
opposite terminus toHSV-1 EcoRI-f, was also clearly underrepresented.Noinformation could
be obtainedonthe HSV-2 terminalBglII
frag-mentd,asthisfragment comigratedwith
HSV-2 fragments c and e. The submolar fragments J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
XbaI
Co x
ch
xI -A
)
I OM
S (I)
IC
N)
Hind
S
I -&
-S CA IC
Eco
RII O
< -A
(A)
S
U)
IC
N)
ab.
cw.
de.
a.#o_O abc
b_ od
C .
Owe
.1de .
f. = :.JO
st_ o9
9.
~ ~ O
m.i °h
9i
aOi
k.°
j
kI.9
*Al
°Me qb -A low& OM
Qi
_
.-_
°i
o
ofn
no.
os .
.0
nfl
0s0a
[image:5.499.50.440.46.636.2]oO
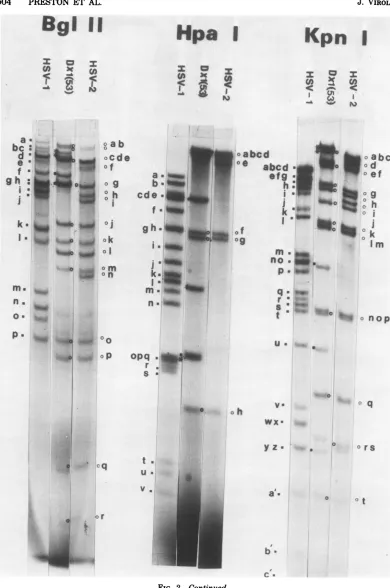
FIG. 2. Autoradiograms of [3PJDNAsof recombinant Dxl(53)and parental virusests1(HSV-2)andtsD
(HSV-1).TheDNAsamplesweredigested with restrictionendonucleases Xba I,HindIII,EcoRI,BglII, Hpa
I, andKpnL Theletters refertospecific HSV-1 orHSV-2 DNAfragments, thephysicalmap locationsof
which aregiven in Fig. 3. Symbols: U, HSV-1 DNA fragment; 0, HSV-2 DNA fragment, *, novel DNA
fragmentproduceduponrestrictionendonucleasedigestion of the recombinant DNA. 503
%*w
bcab. _ d
_t~ e
defga
h.
ia
%wo
°m
on
k.
d
I .%%W
69.
.;...on November 10, 2019 by guest
http://jvi.asm.org/
504 PRESTON ET AL.
BgI
II2: O h
cI
<J. VIROL.
Hpa
I
0 x
Kpn
I
:c =:
I wwl I
C-& %& Ct
oa 0e
cdo
f40~~0
~:f l_
opq .t
S.~~~~~~~~~~~~._
a
to
cq
V.
or
bcd
abcd
fefg
*.h:
!
ak a 00
me
no
.
401
P m
-q _
r.
t a
u.
np.. oh
WX*
yZ a
a'.
b
b
c."
abc
Q_o _od
o ef
ltb
0h
O j
f
o
0k
ImIo.
.'#
*vo
nopO q
0 o rs
[image:6.499.69.459.58.646.2]ot
FIG. 2-Continued.
a a
bc
: d*
I
e a
f
*ai
gh
:i
4
i
a.k. I
I.
m.a
nfl
0.
P
"W
;.
Go
ab ocdE ofg
o l.
o J
ok
oI om
on
- 00
°o ....liE 0
I.
b
on November 10, 2019 by guest
http://jvi.asm.org/
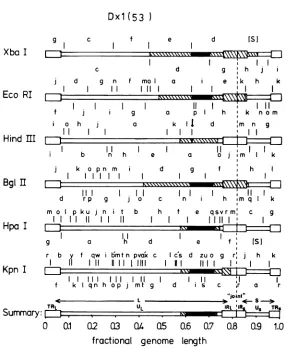
HSV INTERTYPIC RECOMBINANTS 505 Dxl
(53
)
9
Xba I
Eco RI
Hind III
Bgl H I
d
Hpa I
c f e d IS)
D ~
~~~~~~~~
|c d g h j
j d g n f mo a
r
e k h k11 11 I
11 11
f i g a p I h k n om
io h j a k II d im n g
II I
K\NWl
==:El
b n h e
k o pn m d
I I III
l l XXN
I11
rp g j 0
m o I p ku j n t b
I I 11
g a h
C
h
I
d r b y f qw lmtn pva'x c (
Kp 11UI1 11111
KpnfI
lI
f klIqnh op j mt g c
Summary: TRL
11
o J n I I k
g f h I
I. I1
n h im q I k
f e qsvrm, c g
11111
II e
I~f
IfISl
Scs d zu o g r: j h k
I I 11 I
.4JlZ
II I I
s c ,r a r
'joint"
4. S -IRL IR Us TRS
I -i
.L
z1.
[image:7.499.101.387.61.409.2]O 0.1 0.2 03 0.4 0.5 0.6 0.7 0.8 0.9 1.0 fractional genome length
FIG. 3. Analysisofthe crossoverpointsinrecombinantDxl(53), usingsix restrictionendonucleases.
HSV-1physicalmaps have beenalignedwith thoseofHSV-2. Only oneorientation of ULand Usisshown, the
orientationof UL beingthat whichpredominatesinBx1(28)virionDNA. The HSV-1physicalmapsareshown
above, whereas thoseoftheHSV-2areshown below, the genomestructureofthe recombinant. TheHSV-1
HindIII,XbaI,EcoRI, Bgl II,andHpa Imapsaretakenfrom publisheddata(5,26, 31),andthecorresponding
mapsofHSV-2arefrom Cortini and Wilkie (5a). Kpn ImapsofHSV-1 and HSV-2are takenfromthe
unpublisheddataofA.J. Davison andfromMorseetal.(19).The solidregionsofthegenomereferto
HSV-1DNA sequences, and the openregionsrefertoHSV-2 DNA sequences. The crosshatchedregionsindicate DNA sequences in whicha crossoverhasoccurred. Thearrowabove the HSV-1 cleavagesite in theHSV-1 HindIIIphysicalmapreferstothesingleHSV-1HindIIIcleavagesitepresent in the recombinant DNA.
from the S region were present in normal
amounts. These observations suggest that Us
inverted normallyinBxl(28),but the Lregion
appearedtobelargelyfrozen inoneorientation,
the same asthatdepictedin Fig.5. TheHpaI
fragmentpattern ofBxl(28)showed that
HSV-1HpaI-rwaspresent.Thus,theHpaIsite which
delimits Hpa I-mmust also have beenpresent
(Fig. 5). However, HSV-1Hpa I-mwasmissing
from thedigest, and this HSV-1site must
there-fore have remained largely within the IRL
se-quence, in keepingwith observations fromBgl
II andEcoRIdigests. Figure5 shows that inthe
HindIII/Hpa
I double digest the S region of Bxl(28) wascleaved in two places,as wasthe HSV-2 S region, unlike the HSV-1 S region, which was cleaved in three places. If Bxl(28) contained theHpaIcleavagesite whichdelimits HSV-1 Hpa I-m (fragment 12 in theHindIII/Hpa I digest of Fig. 5), then the
HindIII/Hpa Idigest ofBxl(28) DNA should have generated two submolarfragments, of10
x 106and 8.7x
106
daltons(HSV-1 fragment12plus HSV-2fragment8 [12,+82] and12, + 132 in Fig.5). Figure4 shows thepresence of these two fragments, marked with asterisks.
More-,.
_
VOL. 28,1978
_;v
I on November 10, 2019 by guest
http://jvi.asm.org/
506 PRESTON ET AL.
Eco
RI
I w -A >c
_ I1,
N)
_0
_ abc
ab. 4J6P ID V ."
. t
h
° -.m
a.
bc:
e
0
tI%*_.
a
b
cde f .9 h
.;O _A
1*X O'AaK k
o m
., n
n. _
0
....
...
k. ___
... 0 ..- n
p
a.
b-cde.
f.
,;Stiabcd
we
_
la .00
Om '%00
gh. a11 ;.UIf
n. _e
k
_i
Mn.
_Wf
op q
F. m.
.0.,
n.
oh
t.
U
V.
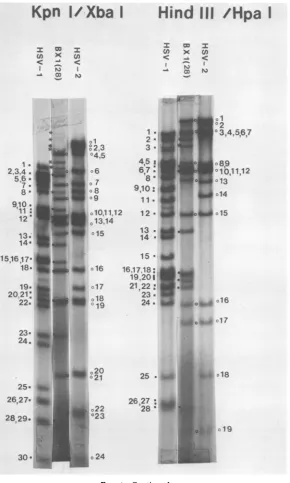
FIG. 4. Autoradiograms of [32P]DNAs ofrecombinantBxl(28)andparentalviruses ts1 (HSV-2)andts B
(HSV-1). The DNAsamplesweredigestedwith restriction endonucleasesEcoRI,BglII,HpaI,HindIIIplus
Hpa I, andKpnIplusXbaI. Theletters inthesingle digestsorthe numbers in the doubledigests referto
specificHSV-1 orHSV-2 DNAfragments,thephysicalmaplocationsofwhich aregiveninFig.5.Symbols:
U, HSV-1 DNA fragment; 0, HSV-2 DNA fragment; *, novel DNA fragmentproduced upon restriction
endonucleasedigestion ofthe recombinantDNA.
J. VIROL.
Bgl
11
I a, I
-r x cn
c: _l c
Hpa I
I W I
ozx n
_ m m
0
-Y i4jAW.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.499.69.456.75.593.2]HSV INTERTYPIC RECOMBINANTS 507
Kpn
I/Xba I
I W X
(n X u)
'C <h
_
- CO N)
*
* _ 1
$ 2.3
04.5
1.
2334. 6
91
o.
12 _ 13.14
7
13.
olJ_1
151617 I
19.
20.21:
22.
23*
24.
25. 26,27.
2829.
30a
C17
n18 19
_
o20
I 21
o22
023
o24
I CC
U) X <
-_Lr
1. 2
-3.
4.5 1
6,7:
8 a
9,10:
111
12*
I N
01
02
°3,4,5,6.7
10,101,12
013
1c.14
_°b O15
13 __a
14
of
15* 16.17.18:
19,201
-. 21,22 :23
24. w0o. 16
JA o* - o17
25 *a U 4* o18
[image:9.499.106.396.71.554.2]o 19
FIG. 4-Continued.
over,they comigratedwithtwofragments
pres-entinHindIII/HpaIdigestsoftwoother
recom-binants, Dxl(43) and Dxl(51), knowntohave
theHSV-1 HpaI-mcleavagesiteandan
HSV-2 Sregion (data notshown, but genome struc-ture isdepictedinFig. 10).Takentogether,the data were consistent with the conclusion that
the virion DNA ofBx1(28)wasoverwhelmingly
in oneorientation ofUL,whereasboth
orienta-tions of the Us region were present in equal
amounts.
Identification of submolar fragments in
Kpn
I/Hpa
I doubledigestsofBx1(28).Anunexpected featureof theKpn I/HpaI double digest ofBx1(28) was the excessive number of
submolar fragments present (Fig. 4). Figure 6
shows how thesesubmolarfragmentscouldhave
beengeneratedby recombination,assumingthat
Hind
III
/Hpa
I
VOL. 28,1978
on November 10, 2019 by guest
http://jvi.asm.org/
508 PRESTON ET AL.
Bxl(28)
d 9 n f mo a
I1I1 111
j k o p n m
_
_11I
d rp g j a
e k h k
't1\%'\\\ \
F71
11 I 11
9 a p h k no m
d 9 f h
I _ _
11
II~~~~~~~~~~~~~~~~~~~~~~~~~~~
c n Ih mq k
j
mo p ku j n it b
II II
II
h 9
1217251811 10 15 826
,,- I, I II
h f e q svrm' c g
11111
_x\\\f
d e f ISI
28
1 5 7 1613 6 1921A201g 9 1422
'_I 4
E I I
10 19 1 16151712 5 14 2 16 9: 13 11 8 31 22 2530 27 32 29
2011 1328 3 19267S17 1518)1fl 5 1128211014X16 4 20 8 6 9
II I Eli111 111 1'1 1. ,1
5 11 1321187 1920914 16 15 1 8 1 3 :22 6 12 1022
24 23
[image:10.499.90.408.62.486.2]0 0.1 0.2
0.3
Q4
0.5 0.6 0.7 0.8 0.9 1.0FIG. 5. Analysis ofcrossoverpoints in the recombinantBxl(28), using restriction endonucleases EcoRI,
BglII, Hpa I,HindIII plus Hpa I, and Kpn I plus XbaLHSV-1 physicalmaps areshown above, whereas
thoseofHSV-2are shownbelow, thegenomestructureof Bxl(28). The solid regions depict known HSV-1 DNA sequences, and the open regions depict HSV-2 sequences. The crosshatched regions indicate DNA
sequencesinwhichacrossoverhasoccurred. Inthe double digests, specific fragmentsaredenoted by numbers
insteadofletters. The shortvertical dottedlines in the HindIII/Hpa I digest refertoHindIIIcleavage sites.
Similarly,those inKpnI/Xba I digests refertocleavage sites ofXbaI.The orientations of ULand Usarethe
same asinFig.3.
(i) the L region was inverted occasionally (as
already discussed) and (ii) that there was an
inequalityintherepetitive regions bounding Us
such thatone of the HSV-2 Kpn I sites which
normally delimited fragment 22 was missing
(thereweretwoof thesecleavage sites in
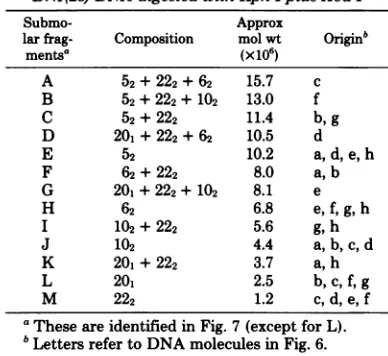
HSV-2DNA.) Table2lists the13submolar fragments
generated by this scheme (fragments A to M)
and their derivations, together with their
frag-ment compositions and expected molecular
weights. Some fragments concordant with the
scheme could beseeninthedigest shown in Fig.
4. To ascertain the identity of these submolar
fragments, 32P-labeled HSV-1 Hpa I-m and
HSV-2 Xba I-h and -iwerehybridizedto
unla-beledKpn I/XbaIdouble-digest-separated
frag-ments of Bx1(28) DNA, using the Southern
technique (27). Figure 7 shows the results and
thecontrol [amixture ofuncleaved HSV-1 and
HSV-2 [32P]DNAs hybridizedtounlabeled Kpn
i
Eco RI
Bgl II
Hpa I
HindIII/Hpa:
KpnI/Xba I
Summary:
rlxxx
*\.1 \ l V
=
J. VIROL.
-i
L--V
on November 10, 2019 by guest
http://jvi.asm.org/
HSV INTERTYPIC RECOMBINANTS 509
5 20+22+6 12 10 22
IlL
_4~~~~~~~~~~~~~
5 ,+ 10 12 6+22
20 5+22} 10 12 6+22*
~~~ ~~~~~~~~~~~
b I t-.C
20 5+22+6 12 10 22
5 20+2246 12 10 22
dZ
5 20+22+6 12 10 22
2 12 6 22
20 5+ 22+10 12 6 22
f 4
20 4415 + 6 12 10+22
9
[image:11.499.49.447.52.316.2]h L
FIG. 6. Explanation forsubmolarfragmentspresent in theKpn I/XbaI doubledigestof Bx1(28) virion DNA. Thegenome structureofBx1(28),showing KpnI andXba Icleavagesites in S and those in L which producetheterminalfragments, isdrawn induplicate atthetopofthefigure.Not allofL is drawn. Two
intramolecular recombinationevents canoccur, andthesearerepresentedbydotted lines. Event1shows that
intramolecular recombinationtotherightofthecleavagesite inTRswhichgenerates the HSV-2fragment22
inverts Usandthiscleavage site.Moleculeais thusgenerated,with this HSV-2cleavagesitenowpresentin IRs. Event 2 shows that intramolecular recombination totheleftofthecleavagesitegeneratingtheHSV-2 fiagment22inverts Usbutnotthiscleavagesite,which remains within TRs. The DNA moleculeeisthus
generated. By subsequent inversion ofL orSorboth regions in molecules a ande, eight different DNA
[image:11.499.46.240.426.604.2]moleculesareproduced.
TABLE 2. Composition of submolar fragments from
Bx1(28)DNAdigestedwithKpn Iplus Xba I
Submo- Approx
larfrag- Composition molwt Originb
mentsa (x106)
A 52+ 222+ 62 15.7 c
B 52+ 222+102 13.0 f
C 52+ 222 11.4 b,g
D 201+222 +62 10.5 d
E 52 10.2 a,d,e,h
F 62+222 8.0 a,b
G 201+222 +102 8.1 e
H 62 6.8 e,f,g,h
I 102+ 222 5.6 g, h
J 10% 4.4 a,b,c,d
K 201+222 3.7 a,h
L 20, 2.5 b,c,f, g
M 222 1.2 C,d, e, f
a Theseareidentified inFig.7(exceptforL).
bLettersrefertoDNAmolecules inFig.6.
I/Xba I
double-digest-separated
fragments
ofBx1(28) DNA] (slots 1 and 5). In the control
tracks, submolar fragments A, B, and C were very faint. Table2 shows thatthese fragments
contain the invertedformof the terminal
frag-ment52(seeFig. 6), again supportingthe
conclu-sion that the Lregion of BX1(28) is present in
mainly one orientation. Xba I-h (double-digest
fragments 62 + 222) hybridized more to A than to B, as would be expected from Table 2. As
expected,XbaI-h alsohybridizedstronglytoD,
F, and H, which contain 62, andto fragment I
andafragment(s) inan areaofthegelwhere K should have been present. Both I and K have
sequences from 222. Similarly,Xba I-i
(double-digest fragments 102 + 222) hybridized more to
fragmentBthantoA. Italsohybridized strongly
toG, I,andJ,allofwhich contain102,andtoD, F, H, andpossibly M, which all havesequences
from 222. Some hybridization to E (contains
sequences within IRL and TRL) was observed.
This was expected, as there are sequences in
common between the repetitive regions
bound-ingUL andthosebounding Us (8).Hpa I-m(Kpn
I/XbaIdouble-digest fragment-20, and41
con-tain sequences present in Hpa I-m) hybridized
strongly to D (20, + 222 + 62), G (20, + 222 +
102), and the Kregion(20 +222).Italso
hybrid-izedto amolarfragment(41in the double digest)
(Fig. 5). All of these fragments have sequences
VOL. 28,1978
on November 10, 2019 by guest
http://jvi.asm.org/
510 PRESTON ET AL.
Kpn I/Xba I DNA
fragments
HSV-2
HSV-w x x =
X cr er X0
-C
6 fo
7
1
2'
42
3
&44.A
9._
10,11!?1 *1 J
18,190.
. 3
i
.-i.
20,21o4
22?,23o* *M ?
240
in common with Hpa I-m (Fig. 5). Thus, the
results of these hybridizations identified the sub-molarfragments. As predicted from the scheme,
-1 fragment 20 in the double digest was not de-tectable inFig. 7. Thisagainsupports the con-ceptthatBx1(28)DNA ispredominately inone
orientationofUL.
Analysis
of IEpolypeptides
induced inselected recombinants. IE polypeptides are defined in this paper asthose earlyviral
poly-5 peptides whose mRNA's are synthesized in
in-fected cellsevenwhen cycloheximide has been
presentthroughout the time of infection. Upon
removal of cycloheximide, these mRNA's are
A translated in the presence of
actinomycin
D,
I
which preventsfurther
RNA synthesis.D Figure8showsSDS-PAGE of IEpolypeptides
E inducedby recombinants from thegeneticcross
t . (ts Bxts 1).The data are summarized in Table 1: 3,whichcomparestheIEpolypeptidesof HSV-4 1and HSV-2 with those inducedby the
recom-binants.Polypeptides VmwIE 175,110, and63of
. HSV-1probably correspondtoICPs4,0, and 27, respectively, describedbyHoness and Roizman
10 (13).
12 Recombinant Bx1(24) induced the Vmw IE
182 of HSV-2, where Bx1(26), Bx1(31-1), and
Bx(31-2) induced the HSV-1 Vmw IE 175.
14
Bx1(28),
however, induced a band containing1.
components whichcomigrated with theVmw IE16 182 of HSV-2 and what appeared to be further
componentswitha greater apparent molecular
weight than Vmw IE 182.
The
Vmw
IE 118/110region
wasofparticular
21 interest. RecombinantsBx1(26)
andBx1(31-1)
induced the HSV-1VmwIE 110,whereas
recom-I
binantBx1(24)
induced the HSV-2Vmw
IE118;
but the two recombinants Bx1(31-2) and 23
Bx1(28)
appeared to induce both the Vmw IE 110 and the Vm,, IE 118. This observationde-pendedonband identification in the gel,and an
4 alternative explanation couldbe that these two
recombinants synthesized as anIEpolypeptide
a polypeptide normally made later in infection whichcomigrated with the Vmw IE118 of
HSV-29 2 (the glycopolypeptide
Vmw
118 ofHSV-2,seenusuallyat 5hpostabsorption).Acomparisonof IE polypeptides induced by the recombinants
FIG. 7. Composition of submolar fragments in the
KpnI/XbaIdouble digest ofBx1(28) virion DNA.
Unlabeled separated KpnI/XbaIdouble digest of
Bx1(28)DNAwasblottedonto nitrocellulose filters
andhybridizedtothefollowing by the Southern tech-nique: equal amounts of HSV-2 and HSV-1 [2JP]
DNAs (slots 1 and 5), 32P-labeled HSV-2 fragment
Xba I-h(slot 2), 32P-labeledHSV-2 fragment Xba I-i * (slot 3), and 32P-labeled HSV-1 fragment Hpa I-m
(slot 4).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.499.64.250.49.650.2]HSV INTERTYPIC RECOMBINANTS 511
1 2 3 4 5 6 7 8 9 10 11 12 13
0(0
C4 CY CY
_
_T
_ mv-x c, x IA x
mC_ m -w m
U,)
co
un
HSV- 2 2 APPARENT
MW X 1
.
*
_182
.i- 157 A -138(144).134,132
-118
-67
-64
FIG. 8. IEpolypeptides induced by recombinants fromthe geneticcross (ts Bx ts1).Slots 1 and13are
mock-infected cellpolypeptide profiles. Symbols: U, HSV-1-induced polypeptide; 0, HSV-2-induced
poly-peptide;*,alteredpolypeptide.Allmolecularweightsareapproximateand basedonestimates made by using
gradient concentration SDS-PAGE. If the apparent molecularweightdiffered when single-concentration
SDS-PAGE was used, then an estimate of the molecular weight based on this gel system is given in parentheses.
with those labeledat5h postabsorption (Fig. 9)
excluded thispossibility,sincenodetectable late
polypeptide comigrated with Vmw IE 118.
More-over,proteolytic cleavagemapsofthe
polypep-tides Vmw 110 and 118ofBx1(28) are identical
tomapsof the VmwIE 110 ofHSV-1 andtheVmw
IE 118 ofHSV-2, respectively (C. M. Preston,
personal communications). This interesting
find-ingis discussed inmoredetaillater.
Knowledge ofthe genome structures of the
recombinants allowsthese IEpolypeptidestobe
mappedonto the genome, ascan beillustrated
with the Vmw IE 63 of HSV-1 and its HSV-2
counterpart polypeptide, Vmw IE 64. All the
re-combinants shown inFig.8induced theHSV-1
polypeptide, which thereforemust map within
the HSV-1 sequences of the recombinant
ge-nomes. Bx1(24) contained the smallest HSV-1
sequenceincommonwith theHSV-1sequences
ofthe otherrecombinants andthus located the
type-specific region ofthis polypeptide to the
map coordinates from 0.56 to 0.79. Figure 13
shows the physical map locations of
polypep-tidesVmw IE175, 110, 87, 68, and 63,derived in
asimilarmanner.
DISCUSSION
Analysis with restriction endonucleases has
demonstrated that 15outof17putative
recom-binants whichwereisolated fromtwocrossesof
VOL. 28, 1978
N _
I
X (A _( U,
:
z
ua
a
(A
i.
4i~~
nz
*t ^iI, lr 1
HSV-1 APPARENT MW X 10 3
175w
136(143). 110
-87
-68_
63W
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.499.49.443.49.438.2]TABLE 3. Summaryof Data on IEpolypeptides
polypeptide HSV-2 Recombinant
polypeptide"
Apparent Apparent
molwt Intensity' molwt Intensity' Bxl(24) Bxl(26) Bxl(28) Bxl(31-1) Bx1(31-2)
(X103)b (X103)b
175 + 182 + 2 1 (a) 1 1
136 (143) V 138 + NI 1 NI 1 NI
- 134/132 + NI ND NI ND NI
110 + 118 + 2 1 1+2d 1 1+2
87 V - 1 1 1 1 1
68 V 67 V 2 1 2 1 1
63 + 64 + 1 1 1 1 1
- 42.5e V 2 ND 2 ND 2
a1, HSV-1 polypeptide; 2, HSV-2 polypeptide; NI, no information; ND, no detectable band; A, altered
polypeptide.
b_,
Noequivalent polypeptidedetectable.+,Intense bandalways present; V, band of variableintensity whichwassometimesnotdetectable.
dSynthesized IE
Vmw
110and IEVmw118.eThe
Vmw
IE 42.5 ofHSV-2probably corresponds to theVmw
43 ofHSV-1, which is seen onlylater in infection.HSV-1 and HSV-2 ts mutants contain both
HSV-1 and HSV-2 sequencesin their genomes
(Fig. 10). Thetwoother recombinants, Dx1(15)
andBx1(26), have identical restriction
endonu-clease maps (with the six enzymes tested) to
thatoftheDNA of HSV-1 and therefore either
are revertants ofts D or tsB, respectively, or
are true recombinants with undetected small
HSV-2 DNAsequences.
Crossovereventshave beenidentifiedinevery
region of thegenome. Mostrecombinantsmust
have been formedbymore than one crossover
event,the greatest number detected in this study
being four[recombinantDx1(51)].
Odd numbers ofcrossovers in UL or Us will
produce recombinants with an HSV-1 and an
HSV-2 repeat bounding the unique region in
whichthecrossovershave occurred.The
recom-binantBx1(31-2), for example, appearstohave
oneHSV-1(L)repeatandoneHSV-2(L)repeat.
Ifthe dataforDx1(2) are included, five of the
recombinants shown in Fig. 10 have odd
num-bersofcrossoversinL, S,orL andS.Figure11
showsthatwherewehavedetectedanodd
num-ber ofcrossovers in one orin both unique
re-gions, then the actual orientation of UL and Us
can alter the minimum number ofcrossovers
necessarytogenerate therecombinant,
assum-ing that the entire LorSorbothregions invert.
However, ifthere are, in addition, crossovers
within the repeat regions, then, depending on
the position of the crossover(s) and the site of
intra- or intermolecular recombination which
leadstoinversion of theuniqueregion, the
min-imumnumberofcrossoversmay or maynotbe
affectedby the orientation of ULorUs,as
illus-trated in Fig. 12. It should also be noted that
intra-orintermolecularrecombinationeventsin such recombinantscangenerateDNAmolecules with differentarrangementsof1 and
HSV-2sequencesfrom that of theparental
recombi-nantDNA(refertoFig. 12).
The virion DNA ofonerecombinant,Bx1(28),
appears tocontainmainlyoneorientation of the
ULregion, althoughboth orientations of theUs regionarerepresentedinequalamounts. Assum-ing that the recombinant was cloned from a
single virus particle, Bx1(28) was formedby a
crossover event in the UL region and another
withinoneof therepetitivesequencesbounding Us. As a consequence, it appears to have one
HSV-1 LrepeatandoneHSV-2Lrepeat bound-ing UL (heterotypic for the L repeat region), whereas those bounding Us appear to be
par-tially heterotypic. After the initial
recombina-tionevents,sequencesinTRs which contain the
HSV-2 KpnI cleavage siteappear to be inter-changeable with the equivalent HSV-1
se-quencesinIRs,whichlackaKpnIcleavage site,
resulting in amixed population ofDNA mole-cules. The result could have arisen if the Sregion
inverts by intra- orintermolecular
recombina-tion on either sideof theKpnIcleavage site (see
Fig. 6).Alternatively,sequencescouldexchange
between TRs and IRs by a double crossover
eventwithintherepetitive region boundingUs.
Itis also feasible that intermolecular recombi-nation couldgenerate molecules which contain
theKpnIcleavagesite in bothTRsandIRsand
molecules which lack thecleavage site in both
TRs and IRs. Further tests on subclones of
Bx1(28)
are currently being carried out inourlaboratory.
Itis not known whythe DNA ofBxl(28) is
J. VIROL.
512 PRESTON ET AL.
on November 10, 2019 by guest
http://jvi.asm.org/
HSV INTERTYPIC RECOMBINANTS 513
1 2 3 4 5 6 7 8 9 10 11 12 13 14
HSV -2
APPARENT
MW X 10-3
IE 182
-IE 138(144) I E 134/132 =
118
-IE 118
IE 64
-IE
42-5-LU LU w LU LU Lu
_- z _-l
N N CV CI Cs z _ _U- o a x x x x
2 2 _- _. =: x (a ax ss to u
.,,w }
,., . A4
_04 -011 a*
FIG. 9. Comparison ofIEpolypeptideswith those made laterininfection (5hpostabsorption)in the absence
of cycloheximide. IEpolypeptidesinducedbyrecombinantsBx1(28) (slot 7)andBx1(31-2) (slot 9), by parental
viruses(slots3and13)andby wild-typeHSV-2 and HSV-1 (slots5and11)areshown. Slot1showsa
mock-infected-cellIEpolypeptide profile. Theremainingslots showpolypeptidesinducedat5hpostabsorption of
virus in the absenceofcycloheximideexceptslot2, which shows themock-infected-cellprofile.
found predominantly in only one arrangement
ofUL. EitheronearrangementofUL is generated
only atagreatly reduced frequency during the
normal replication cycle of Bxl(28) or is
strongly selected against,orthere isa
combina-tion of both possibilities. The DNA we have
analyzed in thisreportwasderived from virions,
and it is conceivable thatboth orientations of
ULwerepresentinnornalamounts in the nuclei
of infectedcellsbut thatonlyonearrangement
waspackaged.
Recombinant Bxl(31-2) also appears to be
heterotypic for the repeats bounding UL, but
with all four genome arrangements present in
virion DNA in equimolar amounts. Both
Bxl(31-2) and Bx1(28) show the minimum
number ofcrossovers when drawnin thesame
orientation of UL in which Bxl(28) is largely
frozen.Morseetal. (19) have also reported that
HSV-1/HSV-2 recombinants formed by odd
numbers of crossovers in UL also showed the
minimumnumberofcrossoverswhen drawn in
the same orientation of UL. This led them to
conclude that their "data were consistent with
thehypothesis that onlyonearrangementofthe
parental DNA participates in the generation of
ul
r-z
cr
I-U,
co m
U,, (A 4-. *-'
HSV-1 APPARENT MW X 10
- 175
- 13fi(143)
- 117
- 110
- 87
- 68
- 63 VOL. 28,1978
'w*x-,4... ,map.
If.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:15.499.35.431.65.461.2]514 PRESTON ET AL.
Dxl(2) T L
Dxl(34-1)
Dxl(34-2), L-' 11~~~
Bxl(24) ---~~~
B 1( )
-l
Bxl(31-2)
Bx1(31-2) . i
l---Bxl1(13) ~
Dxl(31)
Dxl(32)
Dxl(43)
Dxl(48)
Dxl(51)
Dx1(53)
--- --- - ---
[image:16.499.66.261.46.283.2]---o 0.1 02 03 h CB 06 07 G s 1.0 fractional genome length
FIG. 10. Summary ofthegenome structuresof15 HSV-1/HSV-2 recombinants based on datafrom di-gestsofsix restrictionendonucleases. Vertical dotted lines correspond to the endsofthe L andS repeat sequences. The sequences inarecombinant derived from HSV-1 andHSV-2 parents are shown by a thick continuous horizontal linesuperimposedontheupper andlower,respectively, oftwohorizontal dotted lines. A crossover region is shown by two vertical lines between the thick continuoushorizontal lines. The distance between the twovertical lines indicates the region in which the parental nature of the DNA is uncertain. Where the uncertainty is small, the
cross-overregionappears as a singleverticalline. In the
Dxl(2)genomestructure, the crossover in the L
re-peat sequences isquestionable. There are two inter-pretations ofthe data on this recombinant. This crossovermayormay not be present.
recombinants." Although Bxl(28) is partially fixedinthesameorientation ofUL,wefeel that thehypothesismustcontinuetobe treated with
somereservation for thefollowingreasons.
(i) Onlyasmall number of recombinants with odd numbers ofcrossovers in L or S or both regionshave been isolated. Combiningthe data
from Morseetal. (19) and from ourlaboratory,
a minimum number of 7 of 46 recombinants showanodd numberofcrossoversinL. If
recom-binantswithcrossovers close to, orwithin, the
jointregionareincluded, this givesafigureof 14 of 46 recombinants.
(ii) Thegenetic cross used toselect a
partic-ular recombinantmayinfluencethe genome
ar-rangement of theparental virusparticipatingin
the recombinationevent, andtodateonlyavery
limited number of genetic crosses have been
investigated.
(iii) Fine-structure physicalmaps have notyet been made of theIRL andTRL regions of these recombinants. It ispossible theremaybe
unde-tected crossovers within these regions which
causethe number ofcrossovers tobe unaffected
by the orientationofUL (refertoFig. 12). This could explainwhy theBx1(31-2) genome,which
appears to beheterotypic for therepetitive
se-quences bounding UL, unlike Bx1(28), inverts
UL normallytogivefourarrangementsof DNA molecules.
Using information about the physical struc-tureofwild-typerecombinants,tsmutationscan
be located onthegenome.Forinstance,
recom-binants derived from the crosses involving ts 1 allhave incommonthesamesequencesof
HSV-1DNA, and thesemapbetween coordinates0.69
HSV-1 and0.71. We thereforepredictthat this defines
--HSV-2 themaplocation of thetslesion ints 1.Similarly,
tsDlieswithinman coordinates0.83tO0.86and
0.96to0.99 orwithin theregion0.94 to 0.96.The
- L -4 S
TRI IRL IRs TRs
Onecross- _-over
Inversion of L
Two cross- m overs
Two cross
-overs
Inversion of L
4,
Three overs
cross
-Inversion of S
Two cross-overs
Inversion of L
Three cross--_=
[image:16.499.269.461.270.593.2]overs
FIG. 11. Diagrammaticrepresentation of how in-versionof the whole ofLand/orSofHSVDNA could affect the apparent number of crossovers required for thegeneration of a recombinant. Thesolid regions of thegenome refer to HSV-1 DNAsequences, and the open regions refer to HSV-2 DNA sequences.
'JJ
-i ., w---4U
:_ 4--- 7T -! -4 .. -1.
i 11-i
J. VIROL.
.I
Dxl(57)
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 28, 1978
Three crossovers
Three crossovers
Threecrossovers
Four crossovers
Threecrossovers
Three crossovers
Four crossovers
4-El-~~~~~~
b
T hreecrossovers
FIG. 12. Diagrammatic representation of how in-version of the whole of L (a)orinversion bya cross-over within intratypic homologouw regions of the L
repeat(b) could affect theapparentnumber of cross-oversgenerating the recombinant and thestructure
of the recombinant. The solid regions of thegenome
refertoHSV-1 DNAsequences,andtheopenregions
refertoHSV-2 DNAsequences. The horizontal ar-rowsrefertothe orientationof UL.
physicalmaplocations ofthesetwo tsmutations
confirm results frommarker rescuestudies (N.
D. Stowetal. [submitted for publication], who unambiguouslylocatethetsDmutation inTRs
and IRs; P. Chartrand, personal communica-tion). Crossovers alsooccurwhichareunrelated
to events which remove the ts lesion. For
ex-ample,recombinantsDx1(51) andDx1(43)are
formed by crossover events which affect the
short region, but DX1(51), in addition, has a
double crossover in the UL region. From the
genome structures ofother recombinants, this
double crossoverisunlikelytoberelatedtothe
positionof the lesion intsD.
Most recombinantsgrowas well aswild-type
HSV-1 and HSV-2 viruses atboththe
permis-sive and thenonpermissivetemperatures.Thus,
polypeptides fromone serotypeare able to
re-place the functions of polypeptides from the
other withoutimpairing growthof the
recombi-nant virus. Some recombinants, however [for example,Bx1(24)],donotgrowas well as
HSV-1 or HSV-2 atthe nonpermissive temperature.
In such cases, the interaction of polypeptides
fromonetypeofviruswith those from the other
type (or its nucleic acid) maybeimpaired.
Al-ternatively, thecrossovereventitselfmayhave
HSV INTERTYPIC RECOMBINANTS 515
altered the genome ofthe recombinant such that analteredpolypeptide, differing fromeitherthe HSV-1 or the HSV-2 counterpart, is produced which functions lessefficiently. A further possi-bility is that the recombinant genome itself may interact less efficiently with HSV-1 or HSV-2 polypeptides.
Analysis of the IE polypeptides induced by the intertypic recombinants permits physical map coordinates to be assigned to those poly-peptides by correlation with the genome struc-tures. This approach is outlined inmoredetail in anotherpaper(H. S. Marsden et al., J.
Virol.,
in press). It is implicit that HSV-1 and HSV-2 counterpartpolypeptidesarefunctionally equiv-alent. The assumption ofequivalence is based onsimilarityofmobilityofpolypeptidesin SDS-PAGE, kinetics ofsynthesis,amountmade, and post-translational processing of the polypep-tides. Moreover, in the case ofthose polypep-tideswhichundergoposttranslational modifica-tion,it ispossiblethat the
modifying
gene,and not the structural gene, is in fact mapped. It should be noted thatwedonotclaimtomap the entirecoding regionof anypolypeptide,butonly that part whichcauses achangeinmobilityonSDS-PAGE. The possibility that gene splicing takesplaceasinadenovirus and simian virus40
(1, 2) has innoway beenexcludedbyourresults. The summary of map locations of polypep-tides
Vmw
IE 175, 110,87, 68,and63is shown in Fig. 13. Thepolypeptidesmap in regions ofthe genome which contain relatively abundant classes ofIE RNAtranscripts (5, 14; R. J.Wat-son and J. B. Clements, submitted for publica-tion). We conclude that both the L and the S regions of the genome code for IE polypeptides and, at least in the case of
Vmw
IE 182, that the Srepeatregionhasbeenimplicated.Marsdenetal. (in press)have shown that late
polypeptides
arealso made in L and S;thus, neither IEnor
late polypeptides are restricted to the S or L region alone.
In somecases wehave found that the mobili-ties ofpolypeptides specified by recombinants do not correspond to those of either of the parental forms. Forexample, BX1(28) specifies
an IE polypeptide with apparent molecular weightgreater than IEVmw182ofHSV-2aswell
as components which migrate with the upper component of HSV-2IEVmw182.Evidence from
recombinants generated by intertypic marker
rescue(Marsdenetal., in press) suggests thatat
leastpartof thispolypeptide mapswithinTRs and/orIRs. SinceBx1(28)containsa crossover
in IRs (and TRs [see discussion above]) the
sequence ofthe altered polypeptide may con-form in part to HSV-1 and inpart to HSV-2.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:17.499.41.235.68.288.2]516 PRESTON ET AL.
liii
*
IlIllillhllIll liiimilniu
Il
w
*
C'
'~~~~~~~~
1175 175:
'110 _
l
!
:87:
68
.,
63
.~~~~~I0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
J. VIROL.
immediate-early transcripts
immediate-early
polypeptides
genome
[image:18.499.118.414.72.236.2]fractional length
FIG. 13. Summary of physicalmap locationsofIEpolypeptides, togetherwitha comparison ofIE RNA HSV-1transcripts.Thesolidareasofthe IE HSV-1transcripts refertoabundanttranscripts,and the hatched areasrefertoscarcetranscripts.
Alternatively, the polypeptide's mobility could havechanged as aresult ofdifferent processing
ortermination.
Recombinants Bx1(28) and Bx1(31-2)
syn-thesize both theVmw IE 110 of HSV-1 and its HSV-2counterpart, Vmw IE 118.Comparison of IEpolypeptideswith those made later in infec-tiongivesnosupport tothepossibilitythatone
of these isnot a true IEpolypeptide,but isone
normallysynthesized later in infection. This cir-cumstantial evidence isconfirmed by results of proteolytic cleavage ofpolypeptides Vmw IE 110
and 118 induced by the recombinant Bxl(28) (C. M.Preston, manuscriptinpreparation). The simplest explanation is that the polypeptides
map in IRL and TRL, since Bx1(28) is hetero-typic for these repetitive sequences bounding UL.Thisexplanation dependsonthe assumption that the HSV-1 and HSV-2 counterpart
poly-peptides are functionally equivalent. Bx1(24) synthesized only the Vmw IE 118, and, as this recombinant hasoneHSV-2L repeatandone L
repeat which contains HSV-1 and HSV-2
se-quences, this suggests that at least the
type-specific regionsof thepolypeptides Vmw IE 110
and 118map between map coordinates 0.78 to
0.82and0 to 0.04.
We conclude from these results that both repetitive sequences bounding UL can be
ex-pressed. As Bx1(28) is predominantly in one
arrangement of UL, it is possible that genes
withinthese sequences can beexpressed in both
IRLandTRLarrangements. Of course, it is
pos-sible that only one arrangement is expressed
from any one genome. Furthermore, if
expres-sion is from nuclearHSVDNA,whichexists as
concatemers orcircles, then mostrepetitive
se-quencesbounding UL-may atthetimeof
expres-sion beinternal.
ACKNOWLEDGMENTS
We thank R.Cortini formakingavailable restriction en-donuclease maps before publication and S. M. Brown for providingseed stocks of HSV-1 and forhelpfuladvice.
V.G.P. wassupported by a CommonwealthScholarship. LITERATURE CITED
1. Aloni, Y.,R.Dhar, 0. Laub,M. Horowitz, and G. Khoury. 1977. Novel mechanism for RNA maturation: the leader sequences of simian virus40mRNAare not transcribed adjacentto codingsequences. Proc.Natl. Acad. Sci. U.S.A. 74:3686-3690.
2. Berget,S.M., C.Moore,and P. A.Sharp.1977.Spliced
segmentsatthe 5' terminusof adenovirus 2 late mRNA. Proc. Natl. Acad. Sci. U.S.A. 74:3171-3175.
3. Brown, S.M.,D. A.Ritchie,and J. H.Subak-Sharpe.
1973.Geneticstudies withherpes simplexvirus type1: the isolation oftemperature-sensitive mutants, their arrangement intocomplementationgroups and recom-binationanalysis leadingto alinkagemap. J.Gen. Virol. 18:329-346.
4. Clements,J.B., R.Cortini, and N. M.Wilkie.1976. Analysis of herpesvirus DNA substructurebymeansof restriction endonucleases. J. Gen. Virol. 30:243-256. 5.Clements,J.B.,R.J.Watson,and N. M.Wilkie.1977.
Temporal regulationof HSV type 1transcription: lo-cation of transcripts on the viral genome. Cell 12:275-286.
5a.Cortini, R.,and N. M. Wilkie.1978.Physicalmaps for HSVtype2DNA withfive restrictionendonucleases. J.Gen. Virol. 39:259-280.
6. Delius, H.,and J. B. Clements.1976. Apartial denatur-ationmap ofherpes simplexvirustype1DNA:evidence for inversions of the unique regions. J. Gen. Virol. 33:125-133.
7. Esparza, J., M.Benyesh-Melnick,and P. A. Schaffer. 1976. Intertypic complementation and recombination betweentemperature-sensitivemutantsofherpes sim-plexvirustypes1and2.Virology70:372-384. 8. Grafstrom,R.H.,J. C.Alwine,W.L.Steinhart,and
W. C.Hill.1974.Terminal repetitionsinherpessimplex virus type 1 DNA. ColdSpringHarborSymp. Quant. Biol. 39:679-681.
9. Grodzicker, T.,C.Anderson,J.Sambrook,and M. B. Mathews. 1977. Thephysicallocations of structural genesinadenovirusDNA.Virology80:111-126. 10. Halliburton,I.W.,R. E.Randall,R. A.Killington,
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 28,1978
and D. H. Watson. 1977. Someproperties of recom-binants between type 1 and type 2 herpes simplex viruses. J.Gen. Virol. 36:471-484.
11. Hayward, G.S., R. J. Jacob, S. C. Watsworth, andB. Roizman. 1975. Anatomy of herpes simplex virus DNA: evidence for fourpopulations that differ in the relative orientationsof their long and short components. Proc. Natl. Acad.Sci. U.S.A.72:4243-4247.
12.Honess, R.W., and B. Roizman. 1973. Proteins specified by herpessimplex virus. XI. Identification and relative molar rates of synthesis of structural and nonstructural herpes viruspolypeptides in the infected cell. J. Virol. 12:1347-1365.
13. Honess,R.W., and B. Roizman. 1974. Regulation of herpesvirus macromolecular synthesis. I. Cascade regulation of the synthesis of three groups of viral proteins. J.Virol.14:8-19.
14. Jones,P.C., G. S. Hayward, andB. Roizman. 1977. Anatomyofherpes simplex virus DNA. VII. a RNA is homologoustononcontiguoussites in both the Land S components ofviral DNA. J. Virol. 21:268-276. 15. Kieff, E. D., S.L. Bachenheimer, and B.Roizman.
1971. Size,composition, and structure of the deoxyri-bonucleic acid ofherpes simplexvirussubtypes 1 and 2. J. Virol. 8:125-132.
16. Macpherson, I., and M.Stoker. 1962.Polyoma trans-formation inhamstercellclones-aninvestigation of genetic factors affecting cell competence. Virology 16:147-151.
17. Marsden, H. S., I. K. Crombie, and J. H. Subak-Sharpe.1976. Controlofprotein synthesisin herpes-virus-infectedcells: analysis of polypeptides induced by wild type and sixteen temperature-sensitive mutants of HSV strain17.J. Gen.Virol.31:347-372.
18. Mautner,V.,J.Williams,J.Sambrook,P. A.Sharp,
and T.Grodzicker. 1975. The location of the gene coding forhexonand fibreproteinsinadenovirus DNA. Cell5:93-99.
19. Morse,L.S.,T.G.Buchman,B.Roizman, andP. A. Schaffer.1977.Anatomyofherpessimplexvirus DNA. IX.Apparentexclusionof someparentalDNA arrange-mentsin thegenerationofintertypic(HSV-1xHSV-2) recombinants. J. Virol. 24:231-248.
20. Powell,K.L.,and R. J.Courtney. 1975.Polypeptides synthesised in herpes simplex virus type 2-infected HEp-2 cells.Virology66:217-228.
21. Powell, K.L.,R.Mirkovic,and R. J.Courtney.1977. Comparativeanalysis of polypeptides induced bytype 1and type 2 strainsofherpessimplexvirus. Intervirol-ogy 8:18-29.
22. Rakusanova,T.,T.Ben-Porat,M.Himeno,and A. S.
HSV INTERTYPIC RECOMBINANTS 517
Kaplan. 1971. Early function of the genome of herpes-virus. I. Characterization of the RNA synthesised in cycloheximide-treated, infected cells. Virology 46: 877489.
23. Rigby, P. W. J., M.Dieckmann, C. Rhodes, and P. Berg. 1977. Labeling deoxyribonucleic acid to high specific activity in vitro bynick-translation with DNA polymerase I. J. Mol. Biol.113:237-251.
24. Sambrook, J., J. Williams, P. A.Sharp,and T. Grod-zicker. 1975. Physical mappingof temperature-sensi-tive mutations of adenoviruses. J. Mol. Biol. 97:369-390.
25. Sheldrick, P., and N. Berthelot. 1974. Inverted repeti-tions in the chromosome of herpes simplex virus. Cold SpringHarborSymp.Quant. Biol. 39:667-678. 26. Skare, J., and W. C.Summers. 1977. Structure and
functionof herpes virus genomes. II. ECoRl, Xba I and Hind III endonuclease cleavage sites on herpes simplex virus type 1 DNA. Virology76:581-595.
27. Southern, E. M. 1975. Detection ofspecific sequences amongDNAfragmentsseparatedby gel electrophore-sis. J.Mol. Biol.98:503-533.
28. Timbury,M.C.1971.Temperature-sensitivemutantsof herpessimplex virus type 2. J. Gen. Virol. 13:373-376. 29. Timbury,M.C.,and J. H.Subak-Sharpe. 1973. Genetic interactions between temperature-sensitive mutants of types 1 and 2herpes simplexviruses. J. Gen.Virol. 18:347-357.
30. Wilkie,N. M. 1973. The synthesis and substructure of herpesvirus DNA: thedistributionof alkali-labile single strand interruptions in HSV-1 DNA. J. Gen. Virol. 21:453-467.
31. Wilkie,N. M. 1976. Physical maps forherpes simplex virus type 1 DNA for restriction endonucleases Hind III, Hpa-1 and X.bad.J. Virol. 20:222-233.
32. Wilkie,N.M., and R.Cortini.1976.Sequence arrange-ment inherpes simplex virus type 1 DNA: identification of terminal fragments in restriction endonuclease di-gests and evidence for inversions in redundant and unique sequences. J. Virol. 20:211-221.
33. Wilkie,N.M.,N. D.Stow,H.S.Marsden,V.Preston, R. Cortini, M. C. Timbury, and J. H. Subak-Sharpe.1977.Physical mappingofherpes simplexvirus coded functions andpolypeptides bymarker rescue and intertypic recombinants. In G.de The,W.Henle,and F.Rapp (ed.), Proceedingsof the 3rdSymposiumon Herpesviruses and Oncogenesis. I.A.R.C., Lyons, France.
34. Williams,J. F., T.Grodzicker, P. A.Sharp,and J. Sambrook. 1975.Adenovirusrecombination:physical mapping ofcrossover events.Cell 4:113-119.
on November 10, 2019 by guest
http://jvi.asm.org/