0022-538X/79/03-0888/11$02.00/0
Nucleosome-Like Structural
Subunits of Intranuclear Parental
Adenovirus
Type 2 DNA
ALAINSERGEANT,t MICHAELA. TIGGES,ANDHESCHELJ.RASKAS*
Department of Pathology and Department of Microbiology and Immunology, Washington University, School
of
Medicine,St.Louis,Missouri 63110Received for publication 17 February 1979
The intranuclear structure ofparental adenovirus 2 DNAwas studied using
digestion with micrococcal nucleaseas aprobe.When cultureswereinfected with 32P-labeled virions, at a multiplicity of 3,000 particles per cell, 14 to 21% of parental DNA penetrated thecelland reached the nucleus. Of thisparentalDNA, 60% could besolubilized byextensivedigestion with micrococcal nuclease. The nuclease-resistant fraction contained viraldeoxyribonucleoproteinmonomersand
oligomers.Thesenucleosome-likestructurescontained DNAfragmentswhichare
integral multiples ofa unit-length DNA ofapproximately 185 base pairs. The monomeric DNA is similar inlengthto
the4unit-length
DNAcontained in cellular nucleosomes.However, the viraloligomersareslightlysmallerthantheircellularcounterparts. DNA-DNAhybridization demonstrated that all segments of the viralgenome,including those expressedasmRNAonlyatlatetimes, are
repre-sented in the nucleosomal viral DNA. The amount ofearly intranuclear viral chromatin was proportional to multiplicity of infection up to multiplicities of
4,000particlespercell.However,viraltranscriptionalactivitydidnotincrease in directproportiontotheamount ofviral chromatin. Maximum accumulation of intranuclear viral chromatinwasachievedby3hafter infection.The intranuclear parental viral chromatin remained resistant to nuclease digestion even atlate times ininfection,after viral DNAreplicationhadbegun.
Adenovirustype 2replicationrequiresaseries of steps which result in the transfer of virion DNAtothe nucleus of theinfected cell. Studies of virus penetration anduncoating have dem-onstrated that the final steps in this process occur in the nucleus, approximately 1 to 2 h
after infection (24). Although purified virion DNA can be infectious (13, 27) and thevirion DNA has beenreportedtobe free of viral
pro-teinsafteruncoating (24),itseemsunlikelythat naked viral DNA normally serves astemplate
fortranscription. Itisnowwell established that
eucaryotic genes are packaged in chromatin
structures identified as nucleosomes (28).
Nu-cleosomeoligomersand monomers canbe gen-eratedby controlleddigestion of nucleior chro-matin by micrococcal nuclease (18). Quantita-tiveanalysishasdemonstrated thateach
nucleo-some contains about 185 to 200 base pairs of DNAconsisting of a nuclease-resistant core of 140 base pairs wrapped around an octomer of
the four histonesH2A, H2B, H3, andH4. This
coreis connectedtotheadjacentcorebyaDNA
tPresent address:U 102deVirologie,InstitutNational de la Sante etde la Recherche M6dicale, 59045Lille Ledex, France.
segment of15to 100basepairs (18), the linker DNA, which is more susceptible to digestion thancoreDNA.
Similarnucleosome structureshave been de-tected inpapovavirus DNA, both in virions and inintracellularstructurespresentlate in produc-tiveinfection (15, 25, 35). A similarstructurehas beenproposed for the virioncoresofadenovirus
2DNA(7).Ithas been suggested that the virion DNA(23x 106daltons)consists of 180
"nucleo-somes," each containing about200base pairs of DNAwrapped around six copies of the arginine-rich virionpolypeptide VII. Additionaldata sug-gestthat late inproductiveinfection newly
rep-licated viral DNA is also packaged in a
chro-matinstructure (7).
Wehave studied thestructureof intranuclear
parentaladenovirus2DNA incultures infected
withvirions containing radioactive viralDNA.
Using micrococcal nucleasetostudyDNA struc-ture, we have demonstrated that the parental
intranuclear DNAis in achromatin-like struc-ture. The relationship of these structures to
multiplicityofinfection (MOI),thetimecourse
of infection, and viral transcription has been
examined.
888
on November 10, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS 2 DNA INTRANUCLEAR STRUCTURE 889 MATERIALS AND METHODS
Cell culture and virus purification. KB cells
weregrowninsuspensionculturesasdescribed
previ-ously (10). Stocks of3P-labeled adenovirus 2 were
preparedasdescribed (8), and viruswaspurifiedby
cesium chloride centrifugation (10). Viruswas used
within14days after purification. The concentration of 32P-labeled adenovirus 2 preparationswasdetermined
from the optical density at 260 nm based on the
equation, 1optical densityunitat260nm- 1 x 1012
virus particles (24).
Isolation of nuclei. Suspension culture KB cells
wereinfectedataMOI of3,000particlespercellwith
radioactive virusor100PFU/cellwithnonradioactive virus,exceptwhere otherwiseindicated. After 1 h of adsorption,cellswerediluted to 3x105cellspermlin Joklik's minimum essential medium supplemented with 5% horseserum.Unless indicatedotherwise, cul-tureswereusuallyharvested3 h after thebeginningof adsorption, and thecells werewashed twice with cold phosphate-bufferedsaline.
All steps inharvestingcellsorpurifyingnucleiwere
performedat0 to4°C.Thefollowing threemethods
wereusedtopreparenuclei.
(i) Isotonic buffer.Washedcellswereresuspended
ataconcentration of 3x 10' cellsperml in 0.01 M Tris-hydrochloride, pH 7.5, 0.15 M NaCl, 0.0015 M MgCl2, and0.05% Triton X-100(SigmaChemicalCo.). After 15 min at 0°C, cells were disrupted with 10 strokesofatightlyfittedpestleinaDounce homoge-nizer. The nucleiwerepelletedat600xgfor 5minat 40C.
(ii) Hypotonic buffer. Washed cellswere
resus-pendedatadensityof 3 x 106cellsperml in 0.01M
Tris-hydrochloride, pH 7.4, 0.01 M NaCl, 0.0015 M MgCl2, and 0.05%TritonX-100. After 5minat40C, theswollencellswerelysedinaDouncehomogenizer with 20 strokes ofaloosely fitted pestle. The nuclei
werepelletedat600x gfor 5minat4°C.
(iii) Sucrose buffer. Washed cells were
resus-pendedataconcentration of107cellspermlinlysis buffer(0.3Msucrose,0.01 M Tris-hydrochloride, pH 7.9, 0.001 M CaCl2, and 0.5% Nonidet P-40 [Shell Chemical Co.]) (30). Cells were lysed at 4°C in a
Dounce homogenizer as in method ii. Nuclei were
collectedbylow-speedcentrifugation andwashed two times withlysisbuffer and twice withdigestionbuffer (0.3 Msucrose, 0.001 MTris-hydrochloride, pH 7.9, and0.1 mMCaCl2).Insomeexperimentswith isolated nucleidigestionbuffer contained1.0 mMCaCl2,which increasedthe rate ofdigestion bymicrococcal nuclease but didnot alter thepatternoffiagmentrelease with respect todegreeofsolubility.
Procedure iii was used routinely for isolation of nuclei before micrococcal nuclease digestion. Each batch of nucleiprepared bythe above methodswas
foundintact andessentiallyfree ofcytoplasmic
con-taminationby phase-contrastmicroscopy.All nuclear
preparationswereusedimmediatelyfor further
exper-iments.
Isolationofchromatin KBcellslabeledfor72h with[8H]thymidine (final concentration, 0.2 pCi/ml)
were washed with phosphate-buffered salineas
de-scribedabove (method iii). Chromatin was prepared fromthe nuclearpelletasdescribedby Hancock (17), except that 10mM Tris-hydrochloride, pH 8.0, re-placed phosphate buffer. The chromatin pellet was washed twice with 10 ml of 1 mM
Tris-hydrochloride-0.5 mM EDTA (pH 8.0) and twice with digestion buffer. After the final wash the chromatin pellet was disbursed in digestion buffer byshearingin a Dounce homogenizer fitted with a tight pestle. The DNA con-centration was adjusted to 300 to 500
Ag/ml
with digest buffer.Micrococcal nuclease digestion of nuclei. Nu-clei from cultures infected with32P-labeled viruswere prepared by method iii and then resuspended in diges-tionbuffer to give a DNAconcentrationof about 300
pg/ml, asdetermined by the optical density reading (at 260nm) of an aliquot diluted in 5 M urea and 2 M NaCl (30). Thedigestion was carried out at370Cwith micrococcal nuclease (Worthington Biochemicals
Corp.;0.1 U/pg of DNA). Aliquots were removed at
various times,and the amount of labeled DNA ren-dered acid soluble was determined by precipitation with 10% trichloroacetic acid. In some experiments solubilization in perchloric acid was determined as described byLohr et al. (22).
WhenDNAwasto beanalyzed byelectrophoresis, thedigestion was stopped by addition of EDTA, NaCl, and pancreatic RNase A (2). The material was di-gested for 30 min at 370C and then treated with lauryl
sarcosine (0.5% final concentration) and 160 pg of
proteinase K per ml,followedby phenol extraction
(33).When deoxyribonucleoproteins (DNP) were to be analyzed by electrophoresis, the digestion was halted by the addition of EDTA to a 10 mM final concentration. Thenuclear suspension was held on ice for 60 min,sheared,andclearedbycentrifugationfor 10min at 10,000 rpm(SorvallR2B,SS34 rotor). The
supernatantwasused as asource ofnucleosome mon-omersandoligomers(3).
Gelelectrophoresis ofDNA and DNP. Nuclear DNApurifiedafter micrococcal nucleasedigestionand
adenovirus 2DNAdigested withendo R.SmaI (26) wereresolved on either 1.4% agarose slab gels (0.3 by 18by 16 cm) or2.0% agarose tube gels (0.7 by 12 cm) asdescribedpreviously(10).Electrophoresiswas per-formedovernightat 50V at room temperature for slab
gels and 8 h at 1.5 mA pertube forcylindricalgels.
32P-labeled viral DNA wasvisualized by
autoradiog-raphyofgelscovered withplasticwrap and exposed onKodak X-omat R film (XR-5).Theslices (1 mm) fromcylindrical gelswerehydrolyzed at80°C in 0.4 mlof 30% H202containing 1% concentratedNH4OH foratleast4h,neutralized with 1 M acetic acid, and counted in 5 ml ofFormula-963 (New England
Nu-clear). DNP was analyzed in 3.5% acrylamide-0.5% agarose gels (34). Electrophoresis was conducted in thecoldat 20mAfor6h. Afterelectrophoresis, gels were prepared for fluorography (4, 20). 32P-labeled
DNA fagments generated by micrococcal nuclease
digestionof nucleifrom infected cells werehybridized toSmaIadenovirus 2 DNAfragmentsaccording to a method described previously (29), and the hybrids
werevisualizedbyautoradiography.
Invitro RNA synthesisbyisolated nuclei. Pu-rified nucleiwereassayed for RNA syntheticcapacity Voi. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
890 SERGEANT, TIGGES, AND RASKAS as previously described (37, 38), using 500 ,uCi of
[3H]UTP (New England Nuclear, [6,5-3H]UTP) per ml (35 Ci/mmol). After 15min of incubation at 37°C, the mixture was diluted with an equal volume of 0.05 M Tris-hydrochloride (pH 7.9), 0.2 M NaCl, 0.01 M EDTA, 2% N-laurylsarcosine, 200,ug of dextran sulfate perml, 0.002 M UTP, and 300 ,ug of proteinase K (E. Merck; fungal, chromatographically purified and ly-ophilized) per ml. The suspension was pipetted gently, using a large-bore pipette, andincubated overnight at 37°C. RNAs were purified by phenol extraction (39). RNA-DNAhybridization experiments wereperformed with adenovirus 2 DNAimmobilized on 6.5-mm cel-lulose nitrate membranes (type B6, Schleicher & Schuell Co.) (10).
RESULTS
Preparation of nuclei containing
paren-tal viral DNA.Threemethods of nuclear
prep-arationwerecompared (Table 1)before
analyz-ing the physical state of parental viral DNA
earlyininfection. Forallthreemethods, nuclei
weretreated with nonionicdetergentsto remove
cytoplasmic material. Nuclei prepared in iso-tonic(i)orhypotonic (ii) buffers,usedroutinely
forpreparation ofnuclei for in vitro RNA
syn-thesis (9, 38,40), contained the same amountof parental DNA as did nuclei prepared in
su-crose-calcium buffer for micrococcal nuclease digestion (iii). Comparable amounts of viral
RNA were synthesized in vitro with the three
typesof nuclearpreparations (Table 1).
Since nuclei prepared for micrococcal
nu-clease digestion exhibited typical amounts of viral transcriptional activity, we used this methodtostudythe fateandstructureof
paren-tal viral DNA. The subcellular distribution of
parental
32P-labeled
adenovirus2DNA3hafterinfection is shown in Table2.Thepercentageof
parentalDNAfound in nucleirangedfrom14to
21%. To determine if the 32P-labeled DNA in nuclei3hafter infectionrepresentedviralDNA,
nuclear DNAwasanalyzedbyCsClequilibrium
density gradient centrifugation. The results of
such an analysis are shown in Fig. 1. Greater than95% of the32P-labeledDNA had the buoy-antdensity of viralDNA(1.714g/cm3).Asimilar result was obtained when parentalviral DNA
was examined 18 h after infection (data not
shown).Thus, 5%is the upperlimitof viral DNA
moleculesthatmight be covalently attachedto
cellularDNA(16)ortrappedwithcellularDNA, or,alternatively,whichweredegradedand
rein-corporatedintocellularDNA.
Kinetics ofdigestion of intranuclear ad-enovirus 2 DNA by micrococcal nuclease.
The rate at which DNA is renderedacidsoluble wheneucaryoticcell nucleiaredigested by
mi-crococcal nuclease islinear earlyinthe course
TABLE 1. Recovery of32P-labeledadenovirus2 DNA and viraltranscriptional activity in nuclei prepared for micrococcal nucleasedigestion (iii), as
comparedtocrude nuclearpreparations (i,ii)a Intranuclear Viral tran-adenovirus 2 Total scriptional DNA [3H]- activity (% UTP in- hybridiza-Nuclear prepn Ge- corpo- tion of
nomes %of rated [3H]RNA per input (cpm/108) to
adenovi-nu- cells rus 2
cleus DNA)
Isotonic-Tri- 500 15 423,000 1.90 ton (i)
RSB-Triton(ii)b 700 22 490,000 2.60 Sucrose-calci- 600 19 327,000 3.00
um-Noni-det P-40(iii)
aKB
cells
wereinfected with 3x 103particles
percellusing32P-labeledadenovirus 2. After 3 h of infec-tion, nucleiwereisolatedbythe indicated methods,
the amount of intranuclear32P-labeledadenovirus 2 DNA was determined,and the nuclei were incubated invitro for RNA synthesis as described in the text.
[3H]RNAwasextracted from nuclei and hybridized to 10
jig
of adenovirus 2 DNA.RSB,Reticulocyte standard buffer.
TABLE 2. Subcellular distributionofparental
adenovirus2DNA in culturesharvested early in infectiona
% of infective 'P-la-Fraction beledadenovirus2b
CelIsc. 37-58 (47.5)
Postnuclearsupernatant.16-33 (24.5)
Nuclearwashesd . 1.6-4 (2.8)
Nuclei ... 14-21 (17.5)
aKB
cells
wereinfectedwith32P-labeled
adenovirus 2atanMOI of 3,000particles per cell. Cultures were harvested 3 h afterinfection,and nucleiwere fraction-atedbythesucrose-calcium method.bThe numbers give the range of values obtained from sixexperiments. Numbers inparentheses repre-sentaveragevalues.
cKBcells after two washes with cold phosphate-buffered saline.
dNuclear washes include two washes with Nonidet P-40 buffer and two washes with the micrococcal of digestion and gradually reaches a plateau when 50 to 75% of the DNA isdigested (1, 6, 30).
Digestionoftheintranuclear parental
adenovi-rus2 DNAbymicrococcalnuclease in a digest
buffercontaining1.0 mMCaCl2 yielded kinetics of degradation similar to those described for
eucaryotic chromatin; a plateau was reached when about 60% of the viral DNA wasrendered
acid soluble (Fig. 2). The digestion kinetics of
purified adenovirus 2 DNA mixed with
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.272.463.94.240.2] [image:3.503.274.465.356.434.2]ADENOVIRUS 2 DNA INTRANUCLEAR STRUCTURE 891
0.2
0
`0
oi
b
I
I
I
U
0L
(N4
0.1 -10
-1-68
-5
-1.66
10 20 30 40 50
FRACTION NUMBER
FIG. 1. CsCIdensitygradientanalysis of intranu-clear32P-labeledparentaladenovirus2DNA. Nuclei
containing30 pg ofDNA wereprepared 3 h after infection. The nuclear DNAwascentrifugedto
equi-librium in 10-mlCsClgradients (32). Centrifugation
was at100,000Xgfor69hat15°CinaSpinco50Ti rotor.Gradientfractions(0.2 ml) werecollected,and
densitywasdeterminedby refractiveindex.Symbols: *, 32P-labeledadenovirus 2DNA;0, opticaldensity at 260 nm(OD260)ofcellularDNA; , CsCldensity.
Under theseconditions, the desitiesofadenovirus2 DNA and KB nuclear DNA are,respectively, 1.714 and1.698g/cm3.
infected KBcell nuclei weresignificantly
differ-ent. After30 s ofdigestion, as muchas 55% of the
32P-labeled
viral DNA wasdigested, as com-pared to 4% of the intranuclear parental viralDNA (Fig. 2). The digestion of purified viral DNA reached a plateau when about 90% had
been rendered acid soluble. As afurthercontrol
purified
32P-labeled
viral DNA wasmixedwithH-labeledKB cell chromatin andsubjected to digestion. Under these conditions viral DNA was degraded at least as fast as in the absence of chromatin. Forexample,after 1min ofdigestion (micrococcalnuclease, 1
U/,ug
ofDNA), 18%of the[3H]chromatin
countsperminute weresol-ubilized as compared to 70% of the
3"P-labeled
viralDNA.Relationship betweenMOI and the
intra-nuclear concentration of nuclease-resist-antviral DNA. To determine the effect of MOI on intranuclear viral DNA, three parameters
weremeasuredatvarious multiplicities(Fig.3). The amountof intranuclear viral DNA increased linearly up to 4,000 particles per cell. As
meas-uredin isolated nuclei, the amount of viral
tran-scriptional activity increased uptoMOIs of 3,000 to 4,000 particles per cell (Fig. 3A). However, the increase was not directly proportional to MOI.
Nucleipurified from cultures infected at var-iousmultiplicities were digested with micrococ-calnuclease (Fig. 3B). The kinetics of digestion were the same irrespective of the intranuclear concentration ofparental viral DNA. We con-clude that asimilar percentage of the viral DNA is in anuclease-resistant structure irrespective of theMOI.
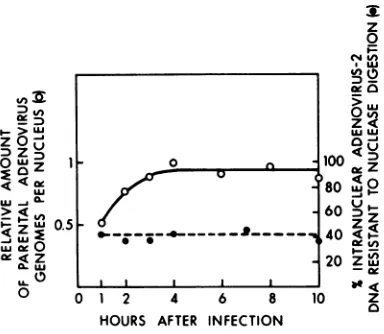
Nuclease resistance of parental viral DNA atvarious times during the course of infection. The fate ofparental viral DNA dur-ing the course of infection was analyzed by har-vestingcultures andpurifying nuclei at various times in infection (Fig. 4). Parental viral DNA
was detectable in nuclei by 1 hafterinfection,
Z 100 In E 80 0 z
c 60
-D 40
6
e 20
u
4
0 10 20 30 40 50
TIME OF DIGESTION (Min) 60
FIG. 2. Kineticsofmicrococcalnuclease digestion of32P-labeledparentalviralDNA inpurified KBcell
nuclei. Nuclei were digested with micrococcal
nu-cleaseasdescribed in thetext.Aliquotswereremoved fromthedigestattheindicatedtimes, and theamount
of32P-labeledadenovirus 2DNArenderedacid sol-uble in 10% trichloroacetic acid was determined.
Symbols: 0, infectious intranuclear3P-labeled
pa-rental viralDNA;0,deproteinized 3P-labeled
ade-novirus 2 DNA mixed with mock-infected KB cell nuclei.
I I a
.
.
aI
VOL. 29,1979
IF
4
9
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.49.234.52.378.2] [image:4.503.252.446.373.545.2]892 SERGEANT, TIGGES, AND RASKAS
10
8
6
0
x
c-uJ z
o
4 - -4 ae
j2I
0 2 4 10
ADENOVIRUS-2 PARTICLES/ CELLx10-3
4c
uJ )Z Zo 4c
a C~4
'.-I;
ZD)
uJ>
,£O
i
_JuJ
00
U)< 0
47
be
Adenovirus-2 Particles/Cell
o400 * 3,000 O 4,000 a10,000
0 20 40 60 80 100
[image:5.503.122.406.70.247.2]TIME OF DIGESTION(Min)
FIG. 3. EffectofincreasingMOIontheamountofintranuclear viralDNA, virus-specificRNAsynthesis in
vitro,and nuclease-resistant viral DNA. Nucleiwerepurified from32P-labeledadenovirus2-infectedcellsby methodiii. The MOIwaseither400, 3,000,4,0X0,or10,000adenovirus 2 virions percell. (A)*, Recoveryof intranuclear3P-labeledadenovirus 2DNA;0,synthesis ofvirus-specificRNAs in nuclei incubated in vitro.
(B)Kinetics of micrococcalnucleasedigestionofintranuclear adenovirus 2 DNA atdifferentMOIs.
1 2 4 6 8
HOURS AFTER INFECTION
x z
0
F-C~4VI
10 0
>z 80
CI-A
400
20 z2
20 ZL
0
FIG. 4.Micrococcal nuclease digestion of intra-nuclearparental adenovirus2DNAatdifferenttimes duringproductive infection.Nucleiwereisolatedby the sucrose-calciummethodfrom KB cellsinfected with 32P-labeled adenovirus2particles.Cultureswere
harvestedattheindicatedtimesduring the produc-tiveinfection. Symbols: 0, Recoveryofintranuclear
3P-labeledparental adenovirus 2 DNA; 0, limit
digestion of intranuclear3P-labeled parental ade-novirus2DNAby micrococcalnuclease.
although maximal transfer to nuclei was not
achieved until4h. Theamountofintranuclear viral DNA remained constant until the latest timesexamined, 10 hafter infection. For each time sample the percentage ofparental 3P-la-beled DNA resistant to micrococcal nuclease digestion (after limit digestion)wasdetermined. Forallsamples tested thepercentwasconstant
(Fig. 4), and the kinetics of digestion were the same (not shown).
Electrophoresis of DNA fragments
gen-erated by micrococcal nuclease digestion.
When nuclei from humancellsaredigested with
micrococcal nuclease, the DNA fragments are
integralmultiplesofamonomeric unit of about
185basepairs, consisting ofacoreof
nuclease-resistant
materialabout140base pairs in length and a heterogeneous nuclease-sensitive linker withameanlength of about45base pairs (19,21, 23). The sizes of cellular and viral DNAs
resistanttodigestionwere compared. KB cells
werelabeled with[3H]thymidinefor72hbefore
infection with3P-labeled adenovirus 2. At3h
afterinfectionnucleiwerepurified and prepared
for micrococcal nuclease digestion. Samples
weretakenfrom the digestion mixtureatvarious
times, and the DNA was extracted and then
fractionatedon2%agarosecylindrical gels (Fig. 5).
Asthe digestion proceeded, thesizes of both
cellularandviraloligomersdecreased.The
high-molecular-weight fragments diminished, and
DNA shiftedtothe mobilitiesofmonomers and
material smaller than 140base pairs.The viral
oligomers were somewhat smaller than
corre-spondingcellular units. Also, the viral nucleo-somes shiftedto smaller sizes at a more rapid
rate than the cellular chromatin. The most
prominent difference between cellularand viral patterns was the rate of accumulation of sub-monomerfragmentsattheexpenseofmonomer.
These submonomers migrated faster than the dyefront (slice 80). Higher-resolutiongels (not
0
K
U)
LU
.-z
U)
uJ
0
0 4
03)
Inv. >..J
z X
0 Z ILL
c
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.70.262.308.475.2]ADENOVIRUS 2 DNA INTRANUCLEAR STRUCTURE 893
m A
a I Mon.
i~~~~~~~
w
.
to nuclease digestion were also displayed as
DNP. Nucleiwereprepared fromunlabeled KB
8.0 cells infected for3h with [3H]thymidinelabeled adenovirus 2. A sample was prepared after 10 6.0 minof digestion; electrophoresiswasperformed
before deproteinization (Fig.6).Aswasthecase
4.0 with viralDNA from infectednuclei,viral DNP
dimers and trimers migrated in the gel more
2.0 rapidly than cellular forms, reflecting the
re-duced DNAcontentofthesestructures (34).
Viral 'nucleosomes" are not formed by association of nuclear proteins with DNA
during digestion. To evaluate the possibility
°.0 that cellular nuclear proteins had exchanged
withviral genomes during the course of diges-5.0 tion, 3H-labeled cellular chromatin wasmixed
4.0 2.0 °
x
0-8.0
6.0
-
TC
Tv
X
4.0
2.0
-Dc
in D
35 15mi.
8.0
3.0-x 2.56.
~,2.0
1.5 ~~~~~~~~~4.0
1.01 2.0
0.5-20 40 60 80 100
SLICE NUMBER
FIG. 5. Micrococcal nucleasedigestionpatternsof
DNA from3H-labeled KB cells 3 h after infection
with 32P-labeledadenovirus2. NucleifromKB cells
infectedwith 3,000virusparticlespercellwere
pre-pared and digested with micrococcal nuclease as
described. Atthe indicated times, sampleswere re-movedfor phenol extraction andelectrophoresis in 2% agarosecylindrical gels.(A)1-mindigest,1%acid soluble;(B)3-mindigest,10% acidsoluble;(C)5-min digest, 15% acidsoluble;(D)15-mindigest,20%acid soluble.
shown) showedthatthese fragments ranged in
size from 20 to 80basepairs.
The parental viral DNA fragments resistant
[image:6.503.52.224.54.525.2]Mq-
X-Mc
FIG. 6. Electrophoretic analysis ofDNP released fromadenovirus2-infectednucleidigestedwith mi-crococcal nuclease. KB cells were infected with [3H]thymidine-labeledadenovirus 2(3,000 particles
per cell). Nuclei were isolated anddigestedas de-scribed in thetext.Electrophoresiswasinacomposite
3.5% acrylamide-0.5% agarose gel. M,, Mc:
Nucleo-somemonomers,viralandcellular;D,T:nucleosome
dimersand trimers. 6.0
4.0 2.0
8.0 0
E 6.0 I 4.0
1
2.0
8.0
6.0
4.0
2.0
-I
-
1-VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.251.438.248.578.2]894 SERGEANT, TIGGES, AND RASKAS
withphenol-extracted, "2P-labeledadenovirus 2
DNA and then digested with micrococcal
nu-clease. The DNA was extracted andsubjected
toelectrophoresis. To maximize the possibility of detecting resistant DNA, the ratio of 32p-labeled DNAto 3H-labeled DNA was 20times
greaterthan thatpresentinisolated nuclei. The results areshown in Fig. 7A through C. Atno
time during the course of digestion did there
appear 32P-labeled DNA fragments of discrete sizes in thegelregionscorrespondingto nucleo-somal DNA.In a separateexperiment(Fig.7D), purified 3H-labeled cellular DNA was mixed withpurified 32P-labeled adenovirus2 DNA. A
similar resultwasobtained.Thus,it is clear that
thenucleosomemonomersandoligomersof viral
DNA inthenucleusare notformedbyexchange
duringdigestion.
Allregions of the adenovirusgenome are
represented in viralnucleosomes. To deter-mine ifall regions of the adenovirusgenomeare
represented in viral nucleosomes, sequences in
nucleosomes were analyzed by DNA-DNA
hy-bridization.Amicrococcalnuclease digestof
in-fected nuclei containing 32P-labeled parental DNA was fractionated on an agarose slab gel. Ethidium bromide staining of the gel showed the typical pattern of cellular DNAfragments
migrating asintegral multiplesofamonomeric unit (Fig. 8, top). The viral nucleosomal DNA was then hybridized tothe setofadenovirus2 DNAfragments producedbycleavage wth endo R-SmaI. The nonradioactive viral DNA frag-ments werefractionatedon aslabgel, denatured,
and then transferred to a nitrocellulose sheet
(29).Thefractionatednucleosomal DNA in the slab gelwas annealed directly tothe DNA on
thenitrocellulose sheet. Thepattern of
hybridi-zation wasdisplayed by autoradiography.
Intra-nuclear viral DNA present as monomers and
monomerrepeatsaslargeaspentamers hybrid-ized toalltheSmaIfragments.The same results were obtainedaftera limitdigestion of the
nu-clei, when the DNA resistant to micrococcal nuclease migrated as a single monomer band
(notshown).
Parental viral DNA retains a chromatin-like structure after viral DNA synthesis begins.It was ofinteresttodetermine whether
the chromatin-like structure of parental viral
DNA wasalso observed after the onset of viral DNA replication. To examine this possibility, 3H-labeled KB cells infected with 32P-labeled adenovirus 2 were continued in cultureuntil 18 hafterinfection;the cellswerethenharvested,
and nuclei were prepared and digested. The DNA wasextractedandsubjectedto
electropho-resis (Fig. 9). As withnuclei 3 h after infection
(Fig.5), achromatin-like structureofviral DNA
J. VIROL.
[image:7.503.286.452.51.537.2]20 40 60 80 100 SLICE NUMBER
FIG. 7. Micrococcal nuclease digestpatterns of DNAfrom 3H-labeled KBcell chromatin mixed with phenol-extracted,32P-labeledadenovirus 2 DNA. The isolation of chromatin anddigestion by micrococcal nuclease are described in the text. Electrophoresis wasin2%oagarosecylindrical gels. (A)1-mindigest, 3Hcpm,0.1%acid soluble;32pcpm,22%oacid soluble. (B)3-mindigest, 3Hcpm,3%acidsoluble; 32pcpm, 37% acidsoluble. (C) 5-mindigest,3Hcpm, 8% acid soluble; 32P cpm, 47% acid soluble. (D) Phenol ex-tracted, 3H-labeled KB cell DNA mixed with
32p-labeled adenovirus2and digested to 25%acid-soluble 3Hcpm and 65%32pcpm.
on November 10, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS 2 DNA INTRANUCLEAR STRUCTURE 895
Direction of Migration
MDT Multimers
I I
Ethidiurm Bromide Staining
I Hybridization to DNA fragments
M D T
Smo I Adenovirus-2
[image:8.503.102.392.63.469.2]_W,..
FIG. 8. Hybridizationtransferof DNA in nucleosomestonitrocellulose-bound endo R.SmaI fragmentsof adenovirus 2 DNA. Nuclei isolatedfrom3P-labeledadenovirus 2-infected KB cells (3 h after infection, 2,000 particlespercell)weredigested for5min withmicrococcal nuclease(500pgof DNAperml; 30Uofenzyme
perml). DNA fragments purified from the digestwere run on a1.4%agaroseslab gel (top). 'P-labeledDNA
intheslabgelwasthenhybridizedtononradioactive adenovirus2SmaI fragments (A-J) which previously hadbeenseparatedon a1%agaroseslab gel andtransferredtonitrocellulose (20). For the hybridizationstep
theradioactivegelwasorientedsuch that thedirection ofmigrationwasatright anglestothe direction of migration of the DNA which had been transferredtonitrocellulose. (Bottom)Pattern of theautoradiogram after the hybridizationstep.M, D, and T designate nucleosomemonomers,dimers,andtrimers, respectively.
wasobserved.Inthesepreparationsalso,a
sub-monomerfractionwasobserved. DISCUSSION
Basedonthe studiespresentedhere,we con-cludethatparentaladenovirus DNA molecules
enteringthe nucleusearlyininfection (3 h) are organizedinapatternsimilar to the nucleosomal
structureofcellular chromatin(Fig. 5); parental
DNA found in the nucleusatlate timesalsohas achromatin-likestructure (Fig. 9). Digestion of nuclei from infected cellswithmicrococcal nu-clease releasesviraland cellularDNPcontaining
DNAsequenceswhicharemultiples ofa
mono-mericunit. It is clear that at least 40%ofthe
viral DNA exists in thisorganization; after 15 min of digestion 40% of the 32P-labeled viral
countsperminute aresmaller thanfull-length
Sma I
Fragments
A _
B C
D
--E
-F
F
G-H VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
896 SERGEANT, TIGGES, AND RASKAS
T0
x
I
SLICE NUMBER
FIG. 9.Micrococcalnucleasedigestionpatternsof DNAfrom 'H-labeled KB cells 18h after infection with3P-labeled adenovirus2. Theinfection, diges-tion,andelectrophoresisconditionsaredescribedin thelegendtoFig.5.(A) 1-min digest, 4% acidsoluble; (B) 3-min digest, 15%acidsoluble; (C)5-min digest, 17% acidsoluble;(D) 15-min digest,20% acid soluble.
adenovirus 2 DNA but equaltoorlarger than
cellularmonomers. Anintracellular adenovirus chromatin structure was notobserved in
elec-tron microscopy studies of nuclear replicative DNAs extractedlate ininfection (18). However,
ourstudy has focusedexclusivelyonthe fate of parental DNA, which is a minor component at late times and might have a structure different from that of themajority of the DNA molecules. The viral nucleosomal products do differ in size from the cellular nucleosomes. Viral
oligo-merDNAs are smaller than the corresponding cellular DNAs and submonomer fragments ac-cumulate more rapidly, suggesting that viral chromatin is moresensitive tomicrococcal
nu-clease both in the linker regionand withinthe
core (Fig. 5 and9). It isperhapsnotsurprising
that viral DNA sequencesare moresensitiveto
nuclease than bulk cellular chromatin. It has
beenshown thattranscriptionallyactiveregions
of hen oviduct chromatinareselectively released
bymicrococcal nuclease(3),and,intermsofthe proportion of the genome engaged in transcrip-tion, viral sequencesaremuchmoreactiveboth
earlyand late than cellulargenes.
DNA inthe submonomerregionwasalso gen-erated when deproteinized DNA was digested (Fig. 7D). Because DNA fragment mobilities tended to be compressed inthis region of the
gels, itisimpossibletodistinguishbetweentwo
alternativestructuresfor the submonomer
frac-tion. Discretefragmentsthat aremultiplesof 10
basepairsaregenerated bymicrococcal nuclease
digestion ofchromatin (31).Alternatively,
ran-domly sized products would be expected from
thedigestionofprotein-freeDNA.Experiments
to characterize further the submonomer DNA arein progress.
Thequantitation of viral chromatin at
differ-ent MOIs has suggested differences between
amounts of chromatin and viraltranscriptional
activity (Fig. 3). Theamountof viral chromatin increased in proportion to MOI up to at least 4,000particlespercell. Viraltranscriptional
ac-tivity, asmeasured in isolatednuclei, appeared
toreachmaximal levelsatsomewhatlower
mul-tiplicities. Thus, quantitation of chromatin
structureisnotnecessarilyadirect reflection of viraltranscription. Viral templates may exist in chromatinstructuresbutnotbeutilized for viral
transcription.Infact,it may bethatonlyasmall
fraction of the viral chromatin isactively
tran-scribed. Although limitations in cellular RNA polymerase concentrations might be responsible forlimitedtemplateutilization,previous studies suggest that other factors may influence
tem-plate-enzyme interaction and thereby determine thetemplates actively transcribed (37).
Analysis of viral chromatin has provided
ad-ditionalevidence for the utilizationofparental genomes in virusreplication. Maximum transfer
of parental DNA to the nucleus required 3 h
from thebeginning ofvirusadsorption, in
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.503.67.254.46.523.2]ADENOVIRUS 2 DNA INTRANUCLEAR STRUCTURE 897
mentwithpreviousstudiesofviralpenetration
(24).Theviral chromatinwasshowntobe pres-ent in nuclei as late as 18 h after infection; at all timesexamined, a constantproportion of paren-tal DNA was resistant to nuclease digestion. Thus, the chromatin-like structure ofparental
DNA waspreserved at least 11 to12hafter viral DNA replication had begun. An earlier study
haddemonstratedthepreservationofa
substan-tial amount ofparentalDNAuntillate in infec-tion(36).Since thelatephase ofviral
transcrip-tion is initiated at the time DNA replication
begins (14), ourfindings demonstrate that this
late transcriptional patterndoesnotrequireor
coincide with theeliminationofchromatin struc-turescontainingparentalDNA strands.
Although allregionsof the viral genomeare
represented inchromatin before the beginning
of viralDNAreplication(Fig. 8),it iscertainly
possiblethatregions containingearlygeneshave
astructuredifferent from those genesexpressed onlyatlate times(11,12,27).Thepolypeptides
responsible forthechromatin-likeorganization
ofparental viral DNA in the nucleus havenot
been identified. Structural analysis of virions
suggestsarole fortheinternal
proteins
VIIandV (7). However, the ability toinfect cells with
nakedDNA (13, 25) indicates that virion
pro-teins are notobligatory forearlyexpression of
the viralgenome. Thus, arole for cellular
his-tones in the early viral chromatin cannot be
excluded.
ACKNOWLEDGMENTS
Weappreciatethetechnicalassistanceof MarkTelle. Com-puter time fordataanalysiawasgenerously provided byM. McDaniel.
Thisstudywaasupported byPublic HealthServicegrant CA-16007 from theNational Cancer Institute and American CancerSocietygrantVC-94.Cell culture mediawereprepared
in a Cancer Centerfacilityfundedbythe National Cancer Institutes. Thisstudywasalso supported bythe following companies Brown&WillismnTobaccoCorp.;Larus and BrotherCompany, Inc.;LiggettandMyers,Inc.; Lorillard,a
Division ofLoewsTheatres,Inc.; Philip Morris, Inc.; RJ. Reynolds Tobacco Co.; UnitedStates TobaccoCo.;and To-bacco Associates, Inc. Thisinvestigation wassupportedin partbyNationalInstitutesof HealthResearchService Award GM 07067,TrainingPrograminCellular and Molecular Biol-ogy,fromtheNationalInstitute ofGeneral MedicalSciences.
LITERATURE CITED
1. Axel,R. 1975.Cleavageof DNA innuclei and chromatin withstaphylococcal nuclease. Biochemistry 14:2921-2925.
2. Axel,R.,W.Melchior,Jr.,B.Sollner-Webb, andG. Felsenfeld.1974.Specificsites of interaction between histonesand DNA in chromatin. Proc.Natl.Acad.Sci. U.S.A. 71:4101-4105.
3. Bloom,K.S.and J. N. Anderson.1978.Fractionation of hen oviduct chromatin intotranscriptionallyactive and inactiveregionsafter selectivemicrococcalnuclease digestion.Cell 15:141-150.
4.Bonner, W.M*,and A.Laskey.1974. Afilmdetection
method for tritium-labeledproteins and nucleic acids in polyacrylamidegels. Eur.J.Biochim.46:83-88. 5. Compton, J. L,AL Bellard,and P. Chambon. 1976.
Biochemicalevidenceofvariabilityin the DNA repeat lengthin the chromatinofhighereukaryotes. Proc. NatLAcad.Sci.U.SA73:4382-4386.
6.Compton,J. I., R.Hancok,P.Oudet,and P. Cham-bon. 1976. Biochemical andelectron-microscopic evi-dence that thesubunitstructure of Chinese-hamster-ovary interphase chromatin is conserved in mitotic chromosomes.Eur.J.Biochem.70:555-68.
7. Corden,J.,HLM.Engelking,and G. D.Pearson. 1976. Chromatin-like organization of the adenovirus chro-mosome. Proc.Natl. Acad. Sci.U.SA.73:401-404. 8.Craig,E.A.,andH. J.Raskas. 1974. Two clasesof
cytoplasmic viralRNAsynthesizedearlyinproductive infection with adenovirus2. J.Virol.14:751-757. 9.Craig,E.A.,andE.J.Raskas.1976. Nucleartrmncripts
largerthan the cytoplasmic mRNAs are specified by segments of the adenovirus genecodingforearly func-tions. Cell8:205-215.
10. Craig, E. A., S. Zinmer, and H. J. Raskas. 1975. Analysis of early adenovirus2 RNAusingEcoR-R1 viralDNAfragments J.Virol.15:1202-1213. 11.Flint, S. J., andH. AL Weintraub. 1977. An altered
subunit configuration associated withtheactively tran-scribedDNAofintegrated adenovirus genes. Cell 12: 783-794.
12. Garel, A., and R. Axel. 1976. Selective digestion of tanscriptionallyactive ovalbumin genesfromoviduct nucleiProc. Natl.Acad. Sci.U.S.A.73:3966-3970. 13. Graham, F.L,and A.J. Van der Eb.1973. A new
techniquefor the assay ofinfectivity of human adeno-virus5DNA.Virology52:456-467.
14. Green,ML,J.T. Parsons,ML Pina,K. Fujinaga,H. Caffier,andLLandraf-Leurs 1970.Transcription of adenovusgenes in productivelyinfectedand intrans formedcells. ColdSpringHarborSymp. Quant. Biol. 35:803-818.
15. Griffith,J.D. 1975.Chromatin structure: deduced from aminichromosome. Science187:1202-1203.
16. Groneberg, J.,D. T. Brown, and W.Doefler. 1975. Uptake and fate of the DNA ofadenovirus type 2 in KBcells. Virology 64:115-131.
17. Hancock,R. 1974.Interphase chromosomal deoxyribo-nucleoproteinisolatedas adiscrete structure from cul-tured cells. J. Mol. Biol.86:649-663.
18 Keer, C.,0. Brian,F.Perrin, and J.Wilhelm. 1978.Structuralanalysisofviralreplicative intermedi-atesisolated from adenovirustype 2-infected HeLa cell nuclei. J. Virol.26:364-379.
19.Kornberg, R. D. 1977. Structureof chromatin. Annu. Rev.Biochem.46:931-954.
20. Laskey,R.A., andD. Mills. 1975.Quantitative film detection of3H and 4Cinpolyacrylamidegels by fluo-rography.Eur.J. Biochem.56:335-341.
21.Lohr,D.,J.Corden,K.Tatcheli R. T.Kovacic, and K. E. VanHolde. 1977.Comparative subunit structure of HeLa, yeast and chicken erythrocyte chromatin. Proc.Natl.Acad. Sci.U.S.A.74:79-83.
22.Lohr, D.,R. T.Kovacic,and K. E.VanHolde. 1977. Quantitatinganalysisof the digestion ofyeast chroma-tinbystaphylococcalnuclease. Biochemistry 16:463-471.
23. Lohr, D.,K.Tatchel,and K. E. Van Holde.1977. On the occurrence of nucleosomephasing in chromatin. Cell12:829-836.
24. Lonberg-Holm, K., and L. Philipson. 1969. Early eventsof vimscellinteraction in an adenovirussystem. J.Virol.4:323-338.
25. Louie, A. J. 1974. The organization ofproteinsinpolyoma and cellular chromatin. Cold Spring Harbor Symp. Quant. Biol.39:259-266.
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
898 SERGEANT, TIGGES, AND RASKAS 26. McGrogan, M., andH. J.Raskas.1977.Species
identi-fication andgenomemapping of cytoplasmicadenovirus type2RNAs synthesizedlate in infection. J. Virol.23:
240-249.
27.Nicolson, M.O., and R.M.McAllister.1972.Infectivity ofhuman adenovirus1DNA. Virology 48:14-21. 28. Oudet, P., M.Gross-Bellard,and P. Chambon.1975.
Electron microscopic and biochemical evidence that chromatinstructureisarepeating unit. Cell4:281-300. 29. Sato, S., C. A. Hutchinson m,and J.I.Harris.1977.
Athermostable sequencespecific endonuclease from thermusaquaticus. Proc. Natl. Acad. Sci. U.S.A.74:
542-546.
30. Sollner-Webb, B., and G. Felsenfeld. 1975. A
compar-ison ofthedigestion of nuclei and chromatin by staph-ylococcal nuclease. Biochemistry 14:2915-2920. 31. Sollner-Webb, B., W. Melchior, Jr., and G.
Felsen-feld. 1978. DNAse I, DNAse II and staphylococcal
nucleasecut atdifferent,yetsymetrically located, sites
onthenucleosomecore.Cell14:611-627.
32. Tibbets, C., K. Johanson, and L Philipson. 1973. Hydroxyapatite chromatography and formamide dena-turationofadenovirus DNA. J. Virol. 12:218-225. 33.Tibbets, C., V. Pettersson, K. Johansoon, and L
Philipson. 1974. Relationship of mRNA from produc-tively infected celistothecomplementary strands of adenovirustype2 DNA. J.Virol. 13:370-377.
34.Todd, R. D.,and W. T. Garrard. 1977. Two dimensional electrophoresis analysis of polynucleosomes. J. Biol. Chem. 252:4729-4738.
35. Varshavsky,A.J.,V. V.Bakayev,P. M.Chumackov, and G. P.Georgiev.1976. Minichromosome of simian
virus40:presenceof histoneHi.NucleicAcids Res. 3: 2101-2113.
36.Vlak,J.M.,T. H.Rozijn,and F.Spies.1976. Replica-tion ofadenovirus type5DNA inKBcells:localization and fate ofparental DNA during replication. Virology 72:99-109.
37. Weinman,R.,J. A.Jaehning,H.J.Raskas,and R. G. Roeder.1976. ViralRNAsynthesis and levels of DNA-dependent RNApolymerases during replication ofadenovirus2.J.Virol. 17:114-126.
38. Weinmann,R.,H.Raskas,and R. Roeder. 1974. Role ofDNA-dependent RNA polymerase II andIII in of the adenovirusgenomelateinproductiveinfection. Proc. Natl. Acad. Sci. U.S.A. 71:3426-3430.
39. Wilhelm,J.,0.Brisoni,C.Kedinger, and P.
Chan-bon. 1976. Characterizationofadenovirus type2 tran-scriptional complexesisolated frominfectedHeLacell
nuclei. J.Virol. 19:61-81.
40. Zimmer,S.G.,C. J.Goldenberg,D.P.Carlson,E. A.
Craig, and H. J. Raskas. 1978. Size distribution of polyadenylatedadenovirus 2 RNAsynthesizedin iso-lated nuclei. Biochemistry 17:4207-4213.
on November 10, 2019 by guest
http://jvi.asm.org/


![FIG. 6.perfrom[3H]thymidine-labeledcrococcalscribed3.5%somedimers Electrophoretic analysis of DNP released adenovirus 2-infected nuclei digested with mi- nuclease](https://thumb-us.123doks.com/thumbv2/123dok_us/1516684.104278/6.503.251.438.248.578/thymidine-labeledcrococcalscribed-somedimers-electrophoretic-analysis-released-adenovirus-infected.webp)



