JOURNAL OFVIROLOGY, Feb. 1979,p.782-788 Vol. 29,No. 2 0022-538X/79/02/0782/07$02.00/0
Novel Bacteriophage Lambda Mutation
Affecting
Lambda
Head
Assembly
C. P. GEORGOPOULOS,I* R. BISIG,2 M. MAGAZIN,2 H. EISEN,2 AND D. COURT3 Departmentof Microbiology, UniversityofUtah, Salt LakeCity, Utah84132';DepartmentofMolecular
Biology, UniversityofGeneva, Geneva,Switzerland2;andDepartmentofMolecularBiology,National CancerInstitute,Bethesda,Maryland200143
Received for publication21June 1978
Anovel phage X
mutation,
called dc10, which interferes with properX head assembly hasbeen isolatedandcharacterized. PhageXcarrying
this mutation is (i) unabletoform plaquesat 30 or37°C but doesso at42°C and (ii) unable to form plaquesat42°ConpN-constitutive hosts. Bothproperties
aredue todclO since all phage revertants for one phenotype simultaneously lose the other phenotype and vice versa. The dclO mutationhas been mapped in the Bgene and has been shown to be dominant over the corresponding wild-type product. At 30°C the dclO mutation results in the formation of abnormal petit X heads made upof pE, pB,pC, andpNu3. UnderpN-constitutive conditions, the dclO mutationresultsin theformation of abnormalpetitXheadsmade up ofpE, Xl, andX2only.A modelto explainthe dataispresented.The bacteriophage A headis an icosahedron of about 60 nm in diameter. On the basis of genetic and biochemical analysis it has been shownthattheprotein products ofatleast nine phage genes (pA, pW, pB, pC, pNu3, pD, pE, pFI and pFII), possiblytwo moredefined by the defective mutations Nul andNu2, and the prod-uct of at least one bacterial gene (groE) are
required for proper head assembly (1). Ofthe headgeneproducts, onlythose of genesB, D, E, and FIIarefound unmodifiedin
purified
infec-tiousvirions. Inaddition, theproducts ofgenes B,C, and E,arefoundinmodified forms. One of these (B*) results from cleavage of intact pB duringassembly. Similarly,theproductof gene Eisfoundasseveralspecies,onepE thoughtto be theunmodifiedproduct,and two others (Xl andX2)whichresult fromcleavageof aprotein formed eitherbyfusionofthe Eand Cproducts or by fusion of cleaved E and Cproducts (6). Thegene Cproduct is foundonlyinthe form of Xl and X2. Theprotein products of the remain-ing head genes are not found in the finished virion. During A infection, a small head struc-ture, called petitA,
about 2/3 the diameter of the mature phage head, can be seen in the electron microscope. Recently, it has been shown that thepetitA can encapsidate A DNA in vitroandsubsequently be joined to a phage tail to give rise to an infectious virion (9, 12), suggestingthat petitA is aprecursor to forma-tion of the phage head. The various forms of petit A resulting from infection with various phage mutants in the head assembly pathwayhaverecently beendescribed, andaschemefor the formation of mature petit X has been pro-posed (7, 10, 12, 14).Briefly,itappears that the
Nu3gene
product
plays
therole ofascaffolding
protein, inasmuchasit isrequired forthe assem-bly of the B and C proteins into the petit A structureand issubsequently removedupon for-mationofXI and X2. Theprocessing of Cand E to form Xl and X2 does not require the presenceof theBprotein, whereas, the process-ing of pBtopB* dependsonthepresence ofan active C gene product. The hostgroE gene is essentialforall oftheseproteinprocesses totake place. Once petit X heads containing pE, pB, pB*, and Xl and X2 have been formed, DNA encapsidation can proceed with the final addi-tions of the D and FIIproteinstoformafinished head structure. The addition of afinished tailto this structure produces an infectious phage A unit.
In this paper we report on the isolation and characterization of a novel phage mutation, calleddc10, which interferes with proper A head morphogenesis. We show that dclO is a cold-sensitive mutation in the B structural gene, that itmakesphage growth sensitive to high levels of Nprotein, and that it is trans dominant over the wild-type B gene product.
MATERIALS AND METHODS
Bacterial strains. The Escherichia coli strains M72 (sup"), TC600 (sup+2) and Ymel (sup+3) were usedas indicator bacteria. A series of W3101 (sup') bacteriacarrying various i434cI+dgalprophages were 782
on November 10, 2019 by guest
http://jvi.asm.org/
LAMBDA HEAD MORPHOGENESIS 783
akind gift of A.Campbell. StrainDC510 isaK-12 his-strr(chiDpgl)Asupoderivativecarrying more than one prophage of the type
iecI857Nsus7sus53nin5
(sus [suppressorsensitive] is identical toam[amber]) and wasusedinisolating strain DC331 which carries the dclO mutation.Phage strains. The various lambdoid phage strains used in this work were from our collection exceptphagecarrying the susa8 mutation in the Nu3 gene, which was a gift of Helios Murialdo. T4 wild type andT4rII169 were agift of Richard Epstein.
Media. T-broth(TB) is 1%tryptone-0.5%NaCl; T-agar is TB plus 1% agar. L-broth (LB) is 1% tryptone-0.5% yeast extract-0.5% NaCl; L-agar is LB plus 1.5% agar. Low-sulfur M9, for labeling with 35SO42 (NewEnglandNuclear),contains per liter:7g ofNa2HPO4, 3 g ofKH2PO4, 0.5 g ofNaCl, 1 g of NH4Cl,11mgofCaCl2,7mg ofMgSO4.7H20,1mg of
thiamine-HCl, 4 g ofmaltose, and all amino acids except methionine and cysteine as described before (4).Adil is10-2MTris(pH 7.5) and10-2MMgCl2. H-buffer is10-2 M Tris(pH 7.7),10-3 MMgSO4,and10-4 Mdithiothreitol.
Bacterial andphageplatings. Platingswere as
previously described (4).
Curing of DC331of residentprophages.Phage i2lcI+b2 at 107/ml was spotted on top ofa lawn of DC331 bacteria and incubatedovernightat30°C. Bac-teria whichsurvived the infectionatthecenterof the spot were grownand tested for ability topropagate phage iAcIat30°C.About 10% of thesebacteriawere
abletoplate phage iAcIat300Candwerejudgedtobe cured of their residentprophagessincetheywerealso unable toplatephagesieOsus29andi21Esus4.
Electron microscopy of lysates. TC600 bac-teriagrowing in TBsupplementedwith 0.2%maltose
at 4 x 108 cells/ml were infected with phage
ixcI60Nsus7sus53nin5Ssus7dc10
atamultiplicity of5perbacterium and grownateither30 or420Cfor2h. The culturewasconcentrated20-fold, lysed with chlo-roform, andcentrifugedat10,000xgfor5min, anda
portion of the supernatantwaslayered on a carbon-coated grid and negatively stained with 1% uranyl
acetatesolution. Sampleswereobserved inaPhilips
300electronmicroscope at anaccelerating voltageof
60kV.
In vitro complementation. Cultures of
M72-(iWcI857Esus4)
andM72(iAcI857Jsus27)weregrown in TB at 300C to about 3 x 108 cells/ml, induced by heatingto420C for20 min, andsubsequentlygrownat370Cuntillysed. Portions of eachlysateweremixed with eachother and withalysate of TC600 bacteria with phage iXcI857Nsus7sus53nin5dcl0 described above for electronmicroscopyandincubatedat370C for 2 h. Production ofviable phage was scored on TC600 bacteria at30°C.
Phagecrosses.Forphagecrosses, bacteriaTC600
were grown in TB-mal to 2 x 108 cells/ml, concen-trated20-fold inAdilandinfectedatamultiplicityof 5phagesof eachtype perbacterium. After20min at roomtemperature forabsorption,the infected cultures
were diluted 10,000-foldinTB andgrownat300Cfor 90min,atwhich timechloroformwasaddedto assure
lysis. Forcomplementationorrecombinationstudies
inwhich oneparentresidesasaprophagein theform
M72(iMc1857),bacteria were pregrown at 30°C in TB-mal,infected at amultiplicity of 5 with phage of the other type, shifted to 420C for 8 min to induce the resident prophage, diluted 10,000-fold into TB and grownat370C for 90 min. For measuring complemen-tation, the progeny phage was assayed on TC600
(W'cI+)
at 420C to detect the iAdclO parent and on TC600 (iAcI+) at 300C to detect the i2l parent. For measuring recombination, the i21sus' recombinants wereassayed on M72(iAbiol0cI857AH1) at300C.The iAdc+ recombinants from thecrosseswith thei434cI+dg prophageswere assayed onTC600 (i43cI+) at 30°C. Preparation ofpetit X heads. TC600 bacteria were grownin M9-lowsulfur at300C to about 2x108 cells/ml, concentrated 20-fold in 10-2 MgCl2 and in-fected withphageat amultiplicity of5per bacterium. After20minat roomtemperatureforadsorption, the infected bacteriawere diluted 20-fold into fresh M9-low sulfursupplemented with 0.2%glucose, 0.5 mCi of carrier-free35SQ42 wasadded, and the cultures were incubated at the desired temperature for 2 h. Cells wereconcentratedbycentrifugationat10,000xg for5min,resuspended in0.25mlof H-buffer, lysed with chloroform, treated with 5 ,ug of DNase I for 5 min at 370C, and centrifuged at 10,000 x g for 10 min to remove cellular debris. Samples of the supernatants were layered on top of a 10-ml 10 to 30% glycerol gradient in H-buffer underlaid with 0.5 ml of CsCl solution ofdensity 1.62in10-2 M Tris (pH 7.5). Gra-dientswerecentrifugedat36,000 rpm for 75min in a BeckmanSW40 rotor, and 20 fractions were collected from the bottom and counted for radioactive material. Under these conditions the petit X head preparations
areusually found in fractions 7 to 12.
SDS-polyacrylamide gel electrophoresis. The procedure for preparing the samples and for
discontin-uous sodium dodecyl sulfate (SDS)-polyacrylamide slabgel electrophoresis were previously described (15).
RESULTS
Isolationof theDC10morphogenetic mu-tation.
(i)
Selection of bacterial strain DC331. Bacterial strainDC510
(iANsus7-sus53cI857nin5)n
>Iwasgrown at320C
in TBtoabout5 x 108cells/ml. The culture was induced for 15 min at
430C
and plated at400C.
Most cells die at this temperature(40°C)
due to de-repression of the prophagelyticgenes.However, temperature-resistant survivorswerefoundat a frequency of 3 x 108. This selection was origi-nallyused in thehope ofidentifyingamongthe temperature-resistant survivors bacterial mu-tantswhichinterfered with theexpression ofthe Q geneproduct of phageX.Suchmutantswould not be able to express phage late functions in the absence ofN andQfunctional geneproducts. Strains carrying suchphages
are thought to survive at a high temperature with the phage existingas aplasmid (3).Atotal of50survivors from 10 independent cultures were tested for abilitytoplate phagei434cI.
Only onesurvivor, calledDC331, provedunabletopropagatei434cI
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
784 GEORGOPOULOS ET AL.
phageat any temperature.
Since phage i43cIis derived fromphageAwith the exception of the immunity region (11), the block exerted by DC331 bacteria on phage growthis eitherat the level ofphageadsorption or at asubsequent intracellularstep.
(ii) Plating
properties
of DC331. Table 1shows the
plating
properties
of variousphages
onDC331bacteria andaDC331 derivative cured of the resident
prophages.
The fact that bothi21b2
andijhAcI
phageplate
well on DC331bacteria suggests that the block exerted
by
DC331 is not atthephage adsorption level but at a subsequent intracellular event. The fact that phage T4rII169 does not propagate on
DC331 bacteria suggests that the lambda rex geneproduct isexpressed constitutivelyat370C. Furthermore, the
inability
of some lambdoid phagederivatesto propagate onthishost isdueTABLE 1. Plating propertiesofvariousphageson strainDC331a
Efficiency ofplating"at38°Con: Bacteriophage
DC331 DC331 cured
iAcI60 <10-6 1.0
i'vir <10-6 1.0
i4.UCI <10-6 1.0
i21cI
0.8 1.0i21cIPsus3 1.2 x10-4 <1o-6 i2`cIOsus29 0.12 <10-6
i21Ssus7 0.5 <10-6
q80cI
0.9 1.0hAi80cI
0.65 1.0T4 1.0 1.0
T4rII169 <10-6 1.0
aStrains DC331 and DC331 "cured" (cured from
prophageA)werepregrownin TB-malat42°Cbefore platingat37°C.
bEfficiency of plating denotes the number of plaques formedonDC331 relativetoDC331 cured. In the case ofphagei'cIOsus29 andi21cIPsus3 the effi-ciency of plating is relative to that on strain TC600(sup'2).
to a prophage-specific mutation since DC331
bacteria cured of their A prophages support
phageXgrowthnormally (Table 1).
(iii) Rescuing of the dclO mutation from DC331 bacteria. To study in detail the
pro-phage mutation(s) which interferes with A
growth, we tried to rescue a phage derivative
carrying this mutation from DC331 bacteria. DC331bacteriawereinfected with phage i43cII
at420Cand thesmallnumber ofphageprogeny
wereplatedonTC600(i43cI+) bacteria at420C. Twotypesofplaquewerefound from thiscross,
small and large. Upon further testing it was
found that the large plaqueswere
indistinguish-able from the iAcI857Nsus7sus53nin5 parent strain. Tenout oftensmall plaques examined, surprisingly, proved unable to propagate on
TC600, Ymel, or M72 bacteria at 30 or 370C.
One of those plaques, called iAcI857Nsus7-sus53nin5dclO,waschosen for furtherstudy.
Characterization of the dclO mutation.(i) Platingpropertiesof the dclO mutation. To facilitate phage growth and studies, we
at-tempted to cross the dclO mutation into a
N+iYcI60
background. In doing the crossiXcI60N+ x iXcI857Nsus53nin5dcl0 we found
out that it was impossible to isolate the iAcI60N+dclO recombinant at anytemperature. It appears, then, that the combination of the
dclO mutationwithawild-type Ngeneis lethal.
Table 2 demonstrates this fact. Phage iAcI60Nsus7sus53nin5dclO, from the above
cross, is unable to propagate on strain M72
(iAbiolOcI857AH1) at 42°C, at which tempera-ture the N gene product is produced
constitu-tively. It appears that both the cold-sensitive phenotype and the N-sensitive phenotype are
due to the same mutation, inasmuch as all
(50/50) "revertants" on TC600 at 300C simul-taneously become abletogrowat420Con
M72-(iWbiolOcI857AH1).
The plating properties ofone such revertant, called Rl, are also shown
in Table 2.Furthermore,all (50/50) plaque
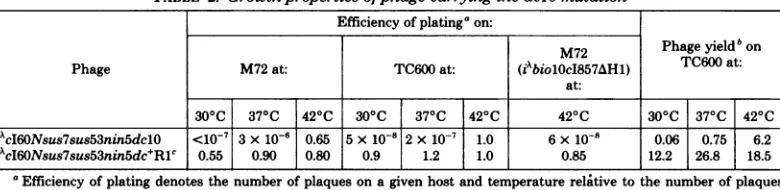
for-TABLE 2. Growthpropertiesofphage carryingthedclOmutation
Efficiency of plating'on:
M72 Phage
yield'
onPhage M72at: TC600at: (iAbiolOcI857AHl) TC600at:
at:
300C 370C 420C 300C 370C 420C 420C 300C 370C 420C
iAcI60Nsus7sus53nin5dclO
<10-7 3x10-6 0.65 5x10-8 2x1O-7 1.0 6x10-8 0.06 0.75 6.2iAcI60Nsus7sus53nin5dc+R1c 0.55 0.90 0.80 0.9 1.2 1.0 0.85 12.2 26.8 18.5 aEfficiency of plating denotes the number of plaqueson agivenhost andtemperature
relative
tothe numberofplaques formedonTC600at420C.bPhage yield denotes the number of viable phageprogeny perinfected bacterium.Themultiplicityof infection was 0.1 phage
per bacterium.
'Isolation asplaqueformerofiAcI60Nsus7sus53nin5dclOonTC600at300C.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.61.451.539.634.2]mersat42°ConM72(iAbiolOcI857AHl)bacteria
simultaneously become able to propagate at 30°Constrain TC600. A reconstruction experi-ment, lysogenizing M72
(supo)
bacteria with phage iAcI857Nsus7sus53nin5dcl0, produced alysogenic strain whichwasunabletoform colo-niesat42°C.Inthisrespectitdiffers from strain DC331andweareforcedtoconclude that in the originalDC331strainatleastanother, and
pos-siblymore,prophage mutation exists in addition
todclO, whichallowsthe bacteriatosurviveat
42°C.Apparently, this additional mutation(s) is not carried by phage iXcI857Nsus7sus53-nin5dclO.
(ii)delOphenotype is dominantoverwild
type. Theresultspresented inTable 1suggest thatatleastone prophage mutation present in
strain DC331 caninhibit the growth of phageX
in trans. Tofindoutif thisproperty is also due tothepresenceofthe dclOmutation,wetested
the ability of dclO mutation carrying phageto block lambdoid growth intrans. Thiswasdone
by mixedly infecting bacteria with phage iAcI60Nsus7sus53nin5dclOandavarietyofother
lambdoid phages. Table 3 shows that the effect of the dclO mutation is more pronounced in
mixed infection experiments with
i'
and i434 phages, whose Ngeneproductsareinterchange-able, and less in mixed infection experiments withi2' phage, whose N function hasadifferent
specificitythan that ofiAN function (2). (iii) In dclO infection tail activity butno
head donoractivity is found. Since bacteria infected with
iecI60Nsus7sus53nin5dclO
phage lyseatall temperatures, itappeared likely that thedclOmutationspecificallyaffectsalatestepduring the phage cycle, atthelevel of heador
tail morphogenesis orboth. This was testedin vitro by mixing lysates of dclO-infected cells
with lysates of iAcI857Esus4 (tail donors) and iAcI857Jsus27 (head donors). Complementation occurred only with the iAcI857Jsus27 lysate (data notshown), suggesting that the block
ex-ertedby the dclOmutationon phage growthis
atsomestepduring head assembly, either at the
level of petitXformationor atthe DNA pack-aging step.
(iv)Electron microscopy of dclO lysates. Electron microscopic examination of lysates fromdclO-infectedcellsconfirmed andextended
the above conclusion. Itwasfoundthatat300C the infected cells contained a normal
comple-ment of tails and that petit A heads were the
only class ofAheadsseen (datanotshown).At
420C, however, in addition to tails and a few
normal heads, abnormal head structures and polyheadswerevisible (datanotshown). Inthis respect the delO lysate resembles infection of
gro+
supo
bacteria by Bsus (13) phage orinfec-tion ofgroE bacteria byAwild-type phage (4).
From these observations it appears likely that
the step during head assembly affected by the dclOis at the level ofproperpetit A assembly
andnot atthesubsequent DNA packagingsteps. (v) Recombination studies with the delO mutation. The above-mentioned conclusion
wasstrengthened by the recombination studies
shown in Table 4.The recombinationfrequency between the dclO mutation and BsuslO was
lower than the frequencies obtained in crosses
with ambermutantsinthe othergenesrequired
for headmorphogenesis.Furthermore, the wild-type allele of the dclO mutationwas rescuable
from an i43'dg deletion which ends between
genesBand C(dg514) butnotfroma deletion
which endssomewhere ingeneB(dg635).
(vi) Complementation studies with the delO mutation. The above conclusionwas
ver-ified inaseries ofcomplementation experiments
between iAcI6ONsus53nin5dclO and various i2l
derivativephage carrying mutations in the head assembly pathway. Itwasfound (Table 5) that
dclOphage complemented allknownhead mu-tants except mutantsin geneB. The lowlevels
ofphage produced in the successful complemen-tationtestsareduetothe partialdominance of
the dclO mutationunder theexperimental
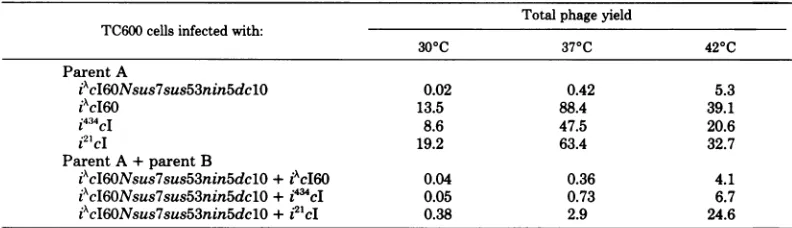
[image:4.505.64.460.548.662.2]con-ditions.Thus,itappearsthatthe dclOmutation TABLE 3. Mixedinfections
Totalphage yield TC600cells infected with:
300C 370C 420C
Parent A
iAcI60Nsus7sus53nin5dclO 0.02 0.42 5.3
iAcI60 13.5 88.4 39.1
i434cI 8.6 47.5 20.6
i21cI 19.2 63.4 32.7
Parent A+parentB
iAcI60Nsus7sus53nin5dc1O+iAcI60 0.04 0.36 4.1
iXcI60Nsus7sus53nin5dclO+i44cI 0.05 0.73 6.7
idcI60Nsus7sus53nin5dclO+
i21cI
0.38 2.9 24.6on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 4. Recombination studieswith the delO mutation
I Parent A Parent B i"sus+recombinants(%)
iXcl60Nsus7sus53nin5dcl0
i2"Asus32
2.3;2.4iecI60Nsus7sus53nin5dc10
i2"BsuslO 0.08;0.19iAcI60Nsus7sus53nin5dclO i21Csus42 0.18;0.25
iAcI60Nsus7sus53nin5dclO i21Nu3susa8 1.7;1.1
iecI60Nsus7sus53nin5dc10
i2'Dsusl5 2.8;1.7Parent B
II ParentA iAdc+progeny
dg deletion endpointin gene
iAcI60Nsus7sus53nin5dclO i4dg651 A 2.1x 102
iAcI60Nsus7sus53nin5dcl0 i4dg615 A-B 1.5x 102
iAcI60Nsus7sus53nin5dclO i4dg635 B 6.7x 10'
iAcI60Nsus7sus53nin5dclO i4dg514 B-C 3.4X 105
iAcI60Nsus7sus53nin5dc10 i4dg614 C 6.4X 105
[image:5.505.58.452.90.245.2]iAcI60Nsus7sus53nin5dclO i4dg652 D 3.9x 105
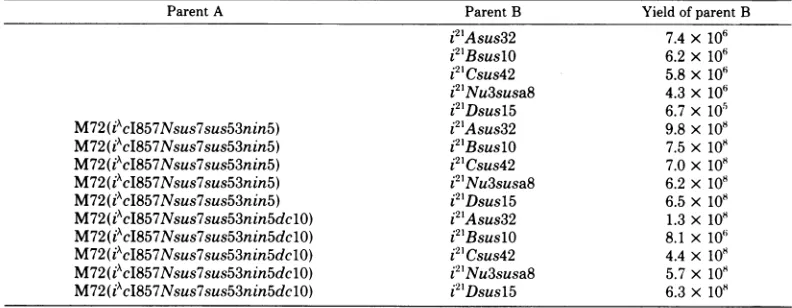
TABLE 5. Complementation studies with thedelO mutation
ParentA Parent B Yield of parent B
i2'Asus32 7.4x 106
i2'BsuslO 6.2x 106
i2lCsus42 5.8x 10"
i2'Nu3susa8 4.3X 106
i21Dsusl5 6.7 x 105
M72(iAcI857Nsus7sus53nin5) i2'Asus32 9.8x 108
M72(iAcl857Nsus7sus53nin5) i21BsuslO 7.5x 10"
M72(iAcI857Nsus7sus53nin5) i2lCsus42 7.0x 108
M72(iAcI857Nsus7sus53nin5) i21Nu3susa8 6.2x 108
M72(iWcI857Nsus7sus53nin5)
i21Dsusl5 6.5x 108M72(iWcI857Nsus7sus53nin5dclO)
i21Asus32 1.3x 108M72(iAcI857Nsus7sus53nin5dclO) i21BsuslO 8.1 x 10'
M72(iAcI857Nsus7sus53nin5dclO) i2lCsus42 4.4x 10"
M72(iAcI857Nsus7sus53nin5dclO) i21Nu3susa8 5.7 x 10"
M72(iWcl857Nsus7sus53nin5dclO)
i21Dsusl5 6.3x 108is in the Bstructuralgene.
(vii) Abnormal
petit
X madeduring
infec-tion
by
dclOphage.
PetitXfractions,
labeledwith 5SO4-2, were prepared from bacteria in-fected with phage carryingthe dclO mutations under various experimental conditions. The re-sults are shown in Fig. 1 and 2 and can be summarizedasfollows. Underconditions where the amount ofN gene product isreduced, e.g., infecting TC600 (sup+2) bacteria with iAcI60-Nsus7sus53nin5dclO, the majority of the petit A heads contain both B and C product in the expected amounts. Some of the C product is processed toXl and X2, and someof the B to B* cleavage takes place at both 37 and 42°C. Surprisingly, under N-constitutive conditions, e.g.,infectinganM72(iAbio10cI857AH1) lysogen pregrownat42°Cfor 4h, very littleBprotein is found associated with the petit N. However, C protein notonly wasfound associated with the petit X structures but wasprocessed to Xl and X2in asubstantialportion of them. This
obser-vationdefinitely accounts for the inability of the phagecarryingthedclO mutation to grow on N-constitutive hosts, although the mechanism of action by which normal amounts of N gene product result in exclusion of the B protein from petitAstructureremains unknown.
The composition of petit X heads made by phage iAc160Nsus7sus53nin5 after infection of TC600 or M72(ibiolOcI857AHl) at 30, 37, or 42°C was essentially that of wild-type petit A heads, i.e., normal amounts of E, B, B*, Xl, and X2proteinswerefound.
DISCUSSION
A novel morphogenetic mutant, called dc10, located in gene Bofphage X has been isolated andpartiallycharacterized. The presence of the dclO mutationinterferes with properphage head morphogenesis both at low temperature and in the presence of constitutive levels ofN protein. Both properties are the result of the dclO mu-tation since reversion to the wild-type
pheno-786 GEORGOPOULOS ET AL. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.56.452.271.425.2]LAMBDA HEAD MORPHOGENESIS 787
typefor either sensitivityresults insimultaneous loss of the other character. The step in head morphogenesis where theB gene product acts is
T 6b A E
to
I3-0
E eL 0
2-U) on
5 9 13 5 9 13 5 9 13 FRACTION NUMBER
FIG. 1. Glycerol gradient profilesofpetit A
prepa-rations labeled with 35SO4-2 prepared as
previ-ously described (4). TC600 bacteria infected with iAcI60Nsus7sus53nin5Ssus7dcl0phage at: (A) 30°C;
(B)37°C; and (C) 42°C. M72 (iAbiolOcI857AHl),
pre-grown at 42°C for 10 generations, infected with
iAcI60Nsus7sus53nin5Ssus7dcl0 phage at: (D)300C; (E)37°C; and (F) 420C.
A B C
anearly one,involvingtheaddition ofthe B and Nu3 products to the petit X head (14). It is knownthattheC gene product can be processed inpetitX heads to Xl and X2 in theabsenceof functionalBproduct. Onthe other hand, theB gene product can be transported to the petitX headstructureintheabsence of C, but no pBto pB* processingor pNu3 exit fromthe petit Acan take place (7, 10, 14). In this respect, the com-positionof thepetitA indclOinfectionsis inter-esting. Under pN-limiting conditions, e.g., growth ofiAcI857Nsus7sus53nin5dcl0phageon
a sup+2host (TC600), themajority of thepetit AmanufacturedaremadeupofpE, pB,pC, and pNu3. Some processingof pB to pB* and pC to Xl and X2takes place, especially at 42°C, but the processing is never complete. Thus, it ap-pearsthatalthough the dclOmutationisinthe Bgene, it somehow interferes also withcomplete processing of theC protein to Xland X2, espe-ciallyat30°C. UnderN-constitutive conditions, however, e.g., growth ofiAcI857Nsus53nin5dclO phage on M72 (iAbiolOcI857AHl) bacteria, the majority ofthepetitA heads manufactured are composed ofpE and Xl and X2. Surprisingly, no B protein is found associated with petit A
D
,& A ,& .
B-_
--,
*B
-fift -Xi
-X2
E F
jAA AA
-C
- ~- E
-Xl
- ~~~~~X2
Nu3-
-Om -1 -Nu3
-_a
IPw
11I 1 1 1 1 1 II 1
5 7 9 4 6 8 67 8
I I I I I11
[image:6.505.53.247.124.269.2]5 7 5 7 7 9
FIG. 2. SDS-polyacrylamide gel electrophoresis ofthe""S-labeledpetitApreparationsshown in Fig. 1. The percent acrylamidewas10. Wild-typeAphage preparationswereusedtomonitor thepositions oftheB,B*, E,Xland X2proteins, andpetitA heads derivedfrom groEA44
(iMcI857Ssus7)
lysogens fortheC andNu3 proteins.29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.55.450.358.622.2]788 GEORGOPOULOS ET AL.
structures atanytemperature. Under these
con-ditions, quantitative processing of pCtoXland X2 takesplace. Thus itappearsthat the BdclO
protein, when not present in the petit A struc-ture, doesnotinterfere with thepCprocessing. In thisrespect, the dclO infection is analogous toBsus infection ofa
gro'sup°
host. Toexplainthe fact that underpN-limiting conditions the BdclO product is transported into the petit A
structurebut under N-constitutive conditions is not, the following model is proposed. The B product,tobe properly assembled into the phage prohead, hastobe acteduponbyahostfactor.
The nature of this action and the subsequent modification of the B protein, if any, is not
known. The host factor could be thegroEgene
product, which has recently been identified on
SDS-gelsas apolypeptide chain of65,000 (5, 8).
Thepresence of the dclO mutation in thegene
product somehow interferes with the proper
modification of the B protein by the host
com-ponent, but does not completely eliminate it. Thus, under N-limiting conditions the levels of Bprotein within thecellarerelativelylowand
the slowed modificationprocesscaneventually
handle the B "processing" so that enough B
geneproduct is assembled into the petitA.Under
N-constitutive conditions, however, higher BdclO levelsaremanufactured,notall ofwhich
canbeprocessed within the time of infectionso
that the BdclOproductcannotbe assembledto the petit A structure atall. This, in turn, may
imply that the B product is transported into petitXincomplexes which contain multiple pB subunits and that the unprocessed BdclO
pro-tein interferes with the functionorassembly of
theprocessed pBorBdclOprotein. This scheme
explains whynoBdclOprotein is found in petit
A under pN-constitutive conditions. The
pres-ence of the dclO mutation may have an
addi-tional effect in slowing down the processing of both the B and Cproteins aswitnessedby the
factthat underlimiting conditionsnotallthepB
to pB* processing and the pC to Xl and X2 processing takes place evenat 420C. The cold sensitivity associated with the dclO mutation could be duetosuchaneffectonthepB and pC
processing.
There are three lines of evidence that give
partial support to the abovescheme, proposed toexplain the effect of the dclO mutation. First,
wehaveshown previously (4) thatsome of the
A mutants, called E, which can overcome the
groE block, mapin thephage Bgene,implying an interaction between this gene product and
the host, groE gene. Second, some evidence
exists that thegroE bacterial product and the B
gene product interact as witnessedby the fact
that they cosediment ina 25Scomplex (Helios
Murialdo, personal communication). Third,
some of thegroEbacterial mutants inour col-lection can be shown, under certain temperature conditions, tomimic the effect of the dclO mu-tation. That is,when infected withwild-typeX phage, the majority of the petitX which accu-mulatecontain normal amounts of Cproteinbut very little or no B protein. Furthermore, some groE bacterialmutants canbe found which
ac-cumulatepetitAwhere thepBtopB* and pCto
XlandX2
processings
areincomplete.
Thismayimply that the groEgeneproduct
plays
adirect rolein thecleavageprocesses.ACKNOWLEDGMENTS
Financial supportwasprovided by grant3-519-75from the Fonds National Suisse de la RechercheScientifique and by Public Health Service grant GM23917 from the National Institute ofGeneral Medical Sciences.
Wethank Sherwood Casjens for reading the manuscript and EthelEldredge for typing it.
LITERATURE CITED
1. Casjens, S.,and J.King.1975.Virusassembly.Annu. Rev. Biochem. 44:555-611.
2. Friedman,D.I.,G. S.Wilgus,and R. J. Mural.1973. GeneNregulator function of phageAimm2l:evidence thatasiteof N action differs fromasiteof N recogni-tion. J.Mol. Biol. 81:505-516.
3. Friedman,D.I.,and M. B.Yarmolinsky.1972. Preven-tion of thelethality of induced A prophage byanisogenic Aplasmid.Virology50:472-481.
4. Georgopoulos, C. P., R. W. Hendrix, S. R.Casjens,
and A. D.Kaiser. 1973. Hostparticipationin bacterio-phage lambda headassembly. J.Mol. Biol. 76:45-60. 5.Georgopoulos,C.P.,and B. Hohn. 1978. Identification ofahostprotein necessary for bacteriophage morpho-genesis (the groE gene product). Proc. Natl. Acad. Sci. U.S.A.75:131-135.
6.Hendrix,R.W.,and S. R.Casjens.1974.Protein fusion: a novel reaction in bacteriophage A head assembly. Proc.Natl.Acad.Sci. U.S.A. 71:1451-1455.
7.Hendrix,R.W.,and S. R.Casjens.1975.Assembly of bacteriophage lambda heads: protein processing and its genetic control in petit A assembly. J. Mol. Biol. 91: 187-199.
8. Hendrix,R.W.,and L.Tsui.1978.Role of the hostin virus assembly: cloning of the Escherichia coli groE gene and identification ofits protein product. Proc. Natl. Acad.Sci. U.S.A.75:136-139.
9. Hohn, B.,and T. Hohn.1974.Activityof empty head-likeparticlesforpackagingof DNA ofbacteriophageA in vitro. Proc.N0tl.Acad.Sci. U.S.A. 71:2372-2376. 10. Hohn,T., H. Flick,and B.Hohn. 1975.Petit A, a family
ofparticles from coliphage lambda infected cells. J. Mol. Biol. 98:107-120.
11.Kaiser,A.D., and F. Jacob.1957.Recombination be-tweenrelatedtemperatebacteriophages and the genetic controlofimmunity and prophagelocalization.Virology 4:509-520.
12.Kaiser, D.,M.Syvanen, andT.Masuda. 1975.DNA packaging stepsinbacteriophage A headassembly. J. Mol. Biol.91:175-186.
13.Kemp, C. L., A. F.Howatson, and L. Siminovitch. 1968.Electronmicroscope studies of mutantsof lambda bacteriophageI.Generaldescriptionandquantitation of viralproducts. Virology36:490-502.
14. Ray, P., and H.Murialdo.1975.Therole ofgeneNu3in bacteriophage lambda head morphogenesis. Virology 64:247-263.
15. Russel,M. 1973.ControlofbacteriophageT4 DNA po-lymerase synthesis.J. Mol.Biol.79:83-94.
J. VIROL.