JOURNAL OF VIROLOGY,May1982,p.456-466 Vol.42, No. 2 0022-538X/82/050456-11$02.00/0
Different
Forms
of Simian Virus 40 Large Tumor Antigen
Varying
in
Their Affinities for DNA
DAVIDGIDONI,1 ARNOSCHELLER,2 BETHBARNET,2 PETROS HANTZOPOULOS,2 MOSHE
OREN,1ANDCAROLPRIVES2*
Virology Department, Weizmann InstituteofScience, Rehovot, Israel,1 and Department of Biological Sciences, ColumbiaUniversity, New York, New York100272
Received19October1981/Accepted14January1982
Invariouspermissive monkeycell lines infected with simian virus 40thereare
two major forms oflarge T antigen which differ in their rate of sedimentation
through sucrose gradients. The lighter (5 to 7S) form sedimented slightly more
rapidly than the 4S tRNA marker, whereas the heavier (16S) form sedimented slightly more slowlythan the 18S rRNAmarker.Thesmalltantigen did not form complexes whichsedimented asrapidlyasthose formedbythe largeTantigen.
The 16S Tantigenformwasconvertedto theslowly sedimenting5 to7S form in
thepresenceof1.0 M NaCl. The majorityoflarge Tantigen synthesized in cell-freeprotein-synthesizing systems primed by mRNA isolated frominfected cells
sedimented as the 5 to 7S form even when premixed with excessquantities of cellularTantigen. The formation of the16Sform ininfected cells did notrequire ongoingviral orcellular DNAreplication because considerablequantities of this T antigen class were producedin thepresenceof DNAsynthesis inhibitors,such as
cytosinearabinoside. Both 5 to7Sand16Sformscould beisolatedseparately and, therefore, each could beanalyzed as to its individual properties. The 5 to7S T
antigen form boundmore efficientlyandtightlytoDNAandhadspecific affinity forsequences at the viraloriginofreplication, whereasthe16Sformbound less efficientlyto DNAandexhibited verylittlespecificity fororigin-containing DNA sequences.It isthereforelikelythat theactiveDNA-bindingspecies of T antigen isolated from infected cells is the5 to 7Sform.
The early region of the genome of the DNA tumorvirus simian virus 40(SV40) encodes two
polypeptides, the large T and small t antigens
(forareview see reference 42). Genetic studies have shown that large T antigen is required for initiating viral DNA replication (36) and viral L-strand-specific RNA transcription (4) and for initiation (12) and maintenance (2, 16, 25, 37) of the transformed phenotype in nonpermissive cells. Inaddition, this geneproduct exerts nega-tivefeedbackcontrol over early mRNA synthe-sis (31, 39). Small t antigen has noapparent role inthelytic cycle of thevirus but appears to play
a role, by slowing tumor growth, in virus-in-duced malignant transformation (17). Although little is known about the properties of small t antigen, considerable information has been ac-quired regarding the biochemistry of the large T antigen. Studies utilizing either authentic T anti-gen fromlytically infected or transformed
cells,
ortheclosely related D-2 protein, have shown it
tobe aphosphoprotein (38) exhibiting both tight
(specific) (23, 34, 40) and loose (nonspecific)
affinities for DNA (3, 23), as well as having ATPaseactivity (41). In addition, both earlier(3,
13, 15, 24, 27) and more recent (5, 6, 18, 28)
papers have described theability of theT anti-gen protein to form rapidly sedimenting com-plexes. Asbothbiological andbiochemical evi-dence supports the idea that T antigen is a multifunctional protein, we sought to isolate different forms of the Tantigen which may have different functions. Morespecifically, we isolat-ed and characterizisolat-ed twoformsof the Tantigen
frominfected cells which differ in their sedimen-tation properties and affinities for DNA.
MATERIALSANDMETHODS
Cells and viruses. BSC-1 or CV-1 lines of African greenmonkeykidney cellswereused,both for infect-edcellextractsandtogrow SV40 virusstrains 777 and 776.
Labeling and extractions of cells. Extraction and labeling of infected cells were done by previously publishedprocedures (29). Generally, cultures of4x 106cells werelabeled with[35S]methionine (100FiCi/
ml) in methionine-free medium from 42to46or 44 to 46 hpostinfection with SV40. All subsequent opera-tionswereperformedat4°C. Cellswerewashed three times inphosphate-buffered saline and then lysed in 0.6 mlof HIPbuffer(pH 8.2)containing0.14 MNaCl, 0.02 M N-2-hydroxyethylpiperazine-N'-2-ethanesul-fonic acid(HEPES),0.001MMgCI2, and0.5%
Noni-456
on November 10, 2019 by guest
http://jvi.asm.org/
DNA BINDING OF SV40 T ANTIGEN OLIGOMERS
det P-40. Phenylmethylsulfonyl fluoride and 1-1-to-sylamide-2-phenylethyl chloromethyl ketone (TPCK) wereadded to a final concentration of 0.25mg/mlof extract immediately before lysis. Nuclei and debris were removed by centrifugation at 2,000 rpm for 2 min.
Sucrosegradientsedhnentation and immunoprecipi-tation. Cytoplasmic extracts (200 p.1) were layered onto5-mlgradients of 5 to20%sucrose in HIP buffer containing 50 p.g offreshly added TPCK per mland 100,ug ofphenylmethylsulfonylfluoride perml. Gradi-entswerecentrifugedat4°Cfor 180minat48,000 rpm in a BeckmanSW50.1swinging bucket rotor. Ten 520-,ul ortwenty260-p,lfractions werecollectedper gradi-ent. After removal of15-p.l samples of eachgradient fraction for direct analysis by polyacrylamide gel electrophoresis (PAGE), the remaining portion was immunoprecipitated with 10,ulofanti-SV40Tserum for1h at 4°C.Immune complexeswereprecipitated by theaddition of 50
IL1
of a10%suspension of formalde-hyde-fixed Staphylococcus A bacteria for 15 min. Immunoprecipitates were washed, suspended in elec-trophoresissample buffer, and subjected to PAGE as previously described (29). Gels were fluorographed, dried, and exposed to Kodak XR-5 film.mRNA preparation and translation. Cytoplasmic polyadenylic acid-containing RNA was purified from SV40-infected cellsandtranslatedin thenuclease-free reticulocyte lysate system according to previously publishedprocedures (30). mRNA (20 ,ug) was added per 500Illof translation reaction mixturecontaining 400
IuCi
of[35S]methionine.Translation products were layeredonto 5 to20%o sucrose gradients, centrifuged throughsucrose gradients,andimmunoprecipitatedas above.DNA-bindingexperimepts. Binding of T antigen in sucrose gradient fractionsto DNA-cellulosecolumns wascarried out byprocedures described by Oren et al. (23). Samples (200 ,ul) of[35S]methionine-labeled
infected cell extracts were centrifuged through five sucrosegradients run in parallel, separately fractionat-ed, and stored. One set of gradient fractions was immunoprecipitated with anti-T serum, subjected to PAGE, and then autoradiographed to identify peak tubes of T-antigen-sedimenting forms, which were thenpooledseparately from the other individual gradi-ents. Samplestobebound to DNA wereadjustedto pH 7.0 with2-1lincrementsof 0.1 N acetic acid and loaded onto a 1-ml calfthymus DNA-cellulose column orF-161SV40multioriginvariantDNA-cellulose col-umn(23)which had beenpreequilibratedwith 10 mM potassium phosphate-0.1MNaCI-109o(vol/vol) glyc-erine-0.5% NonidetP-40, pH 7.0. Afterallowing 30 minforthesampletobind,thenonbound fractionwas
collected,and the columnwaswashedwith
equilibra-tion buffer.Boundproteinswereelutedstepwisewith bufferscontaining0.4 Mand 1.0 MNaCI.Nonbound and boundelutedfractionswerethenadjustedto0.15 MNaCI, immunoprecipitated with anti-T serum, and subjectedtosodiumdodecyl sulfate-PAGEand
auto-radiographyasabove.
DNAbindinginsolutionwasperformed byrecently
describedprocedures (A. Scheller,L.Covey-Nichols,
B.Barnet, C. Prives, submittedforpublication).
Su-crosegradientfractionswereadjustedtopH6.5 with
2-01
samples of0.1 Nacetic acid. Five nanograms ofBstNl-digested SV40DNAlabeledwith 32Pby
nick-translation (32), as well as 5 ,ug ofsheared salmon sperm DNA, was added to each fraction for 1 h followed by immunoprecipitation of the T antigen DNA complexes with 10 pl of anti-T serum and StaphylococcusAbacteria as above. After the com-plexes had been washed three times, DNAfragments were released from the immune complexes with 50Il1
of buffer containing 0.01 M Tris, 0.001 M EDTA, and 0.5% sodium dodecyl sulfate (pH 7.4), subjected to electrophoresis through 2.0%'o agarose gels in Tris-boratebuffer,thendried,andautoradiographed.
Materials. [35S]methionineand
[32P]deoxynucleoside
triphosphateswerepurchasedfromtheRadiochemical Centre, Amersham, England. TPCK, phenylmethyl-sulfonyl fluoride, hydroxyurea, 2'-deoxy-2'-azidocyti-dine, aphidicolin, and cytosine arabinoside were boughtfrom SigmaChemical Co., St. Louis, Mo., and BstNl restriction endonuclease was purchased from New EnglandBiolabs, Beverly, Mass. Staphylococ-cus Abacteria were purchased from Calbiochem, La Jolla, Calif., as Pansorbin. Anti-SV40 T serum was obtained from the Research Resources Branch, Na-tional Cancer Institute, NaNa-tional Institutes ofHealth, Bethesda,Md.RESULTS
Existenceof Tantigen as two
major
sediment-ing forms in infected monkey cells. To extendstudies describing different oligomericforms of
T antigen, extracts of
[35S]methionine-labeled
BSC-1 cells were subjected to sedimentation through sucrose gradients. Individual
gradient
fractions
werecollectedandimmunoprecipitat-ed with anti-T serum, followed by PAGE and
autoradiography. Undertheconditionsof
infec-tion,labeling,
extpWption,
andcentrifugationde-scribed herein, tpe
large
T antigen appears astwo major sedimenting classes (Fig. 1). The
lighter class cosedjienting with 5S RNA and slightly less
rapidly'
than aldolase (molecular weight, 159,000), wastermedthe 5 to7S form.The heavier class,
se4imenting
lessrapidlythanitS
rRNA or thyroglobulin(molecular
weight,670,000),
was termed the 16Sform. Itwas notfeasibletoassign definitive sedimentation coeffi-cients or molecular weights for these
forms,
isolated as they are from the crude extracts of infected cells. In many
experiments
a45,000-molecular-weight
polypeptide
was immunopre-cipitated from allgradient
fractions. This maywell
bethemajorcapsidprotein
VP-1which has this molecular weight, reactswith many anti-Tsera(35),andforms
rapidly
sedimenting
assem-blycomplexes
(26).
The small t
antigen,
although
sedimenting
somewhat more
rapidly
thanexpected
for a17,000-molecular-weight
polypeptide,
was notdetectable in
gradient
fractionscontaining
the 16SformoflargeTantigen
andtherefore doesnot
participate
in therapidly
sedimenting large
Tantigen
complexes.
The sedimentationproper-ties oflarge Tand smallt
antigens
wererepro-VOL.42,1982 457
on November 10, 2019 by guest
http://jvi.asm.org/
J. VIROL. 458 GIDONI ET AL.
18$
4, :l
a
_S. - s1..
b 3s r
" _ ,,., - ...
'S,8 ,"110 4..
tg :pI :._". #W. .:
Ju. 1. Sucrosegradient sedimentationofSV40T
antigen. [35S]methionine-labeled cell extracts were
subjected tocentrifugation through5to20%sucrose
gradients asdescribed in the text. Twenty fractions
were collected, and 10-,ul samples of each fraction
weredirectlyanalyzed by PAGE andautoradiography
(a).Theremainderof eachsamplewas immunoprecip-itated withanti-SV40Tserumandsimilarly analyzed
(b).Twoparallelsucrosegradientswererun, contain-ing [3H]uridine-labeled rRNA markers in one and purified preparations of aldolase andthyroglobulinin the other. Peak tubes of 5S, 18S, and 28S rRNA markers were nos. 4, 11, and 17, respectively, and
peaktubesofaldolase(molecular weight, 159,000) and thyroglobulin (molecular weight, 670,000)werenos.5 and11,respectively.
duciblyobserved in several SV40-infected
mon-key lines, including BSC-1, CV-1, TC-7, and
AGMK cells. The relative proportionof the 5to
7S and 16S large T antigen form varied among
differentmonkey cell lines.Generally, therewas
more [35S]methionine-labeled 5to7S form than
16S formimmunoprecipitatedfromsucrose
gra-dientfractions. However, therewas more
rapid-ly sedimentingthan slowly sedimentingT
anti-gen when this protein was measured by the
enzyme-linked immunosorbent assay,
per-formedasrecently
published
(11,43),
indicating
that muchoftheaccumulatedTantigen
existsas heavier complexes (data not shown). As theexperiments in this study measured the
[35S]methionine-labeled
T antigen, it should be kept in mind that all the datapresented
refertothat population ofT antigen molecules
synthe-sizedduring thelabelingperiod of2to4 hbefore extraction.
Tantigensynthesizedinvitro.Studiesinother laboratories (5, 6, 13), as wellas our own
(28),
have shown that tsA mutant Tantigendoesnotform rapidly sedimenting complexes at the
re-strictivetemperature,
suggesting
that the gener-ation of the 16S form is related to abiological
function ofT antigen. To further explore this idea, weexaminedthesedimentationproperties
ofT antigen
synthesized
in acell-freeprotein-synthesizingsystemprimedwithSV40E-strand
mRNA.This Tantigenwouldserve as an
exam-ple of nonfunctional protein in that it has no opportunity to act in viral and cellular DNA
replication or RNA transcription. Moreover, it may notundergo theidentical
post-translational
modificationscharacteristicof Tantigen isolated
from infected cells.
When Tantigen synthesized inthe nuclease-treated reticulocyte
lysate
protein-synthesizing
systemwascentrifuged throughasucrosegradi-ent, noevidenceoftherapidlysedimentingform was detected (Fig. 2). Even afterco-incubation
and co-sedimentation of the cell-free
reticulo-cyte extractwith unlabeled extracts of infected
or uninfected cells, the in vitro-synthesized T antigen existed only as the 5 to 7S form. The results shown inFig. 2, in which 10 fractions per
gradient were collected, were confirmedin
ex-periments with 20 fraction gradients, such as those normally used in otherexperiments.This supports the idea that the 16S form of T antigen isaproperty of native,functional protein. Either
a specific post-translational modification or some aspect of the biologicalfunction of the T antigen protein is responsible for theability of this viral protein to form rapidly sedimenting complexes.
16S
Tantigenformation.Neither tsAmutant T antigen at restrictive temperature nor in vitro-synthesized T antigen forms oligomers. Both are nonfunctioning T antigens, the former due to its conditional defect, the latterbecause of its lack ofopportunitytofunction or lack of proper post-translationalmodification orboth. Another link betweenthese twonon-oligomerizingTantigens isthe absence of DNAreplication. tsA mutant-infected cells failed to initiate viral or cellularDNAreplication at thenonpermissive tempera-ture, andclearly there was no DNAreplication
on November 10, 2019 by guest
http://jvi.asm.org/
DNA BINDING OF SV40 T ANTIGEN OLIGOMERS 459
i
a
2 4 6 8 10
r
.. Ams Wa 0T-.
ep3q.
C
.. .*a a
C
t 4 6 8 10
. .. 6
o..:.. !1
_ *+* a | j
i- :s.
e
:M.'{:BiE'-t
..::Mr
******
X .F
!
tv :
__....-
_ J2 4 6 8
T- n,
[image:4.505.115.404.69.515.2]d
FIG. 2. Sedimentation of T antigen synthesized in vitro. SV40Tantigenwassynthesized inthereticulocyte
translation system (0.5-ml reaction volume) in response to mRNA from SV40-infected cells. Translation products (100 gxl)weremixed with 100 Il of HIP buffer (a); 100 II of infected unlabeled cellextract(b); 100 ILl of uninfected unlabeled cellextract(C) for 1 hat4°C followed bysucrosegradient centrifugation,
immunoprecipi-tation, and analysisasdescribed in thelegendtoFig.1.[35S]methionine-labeled Tantigen (100 ILI)from SV40-infectedextractwassimilarly analyzedon sucrosegradient (d).Ten fractionswerecollectedfrom eachgradient. That 5 to 7S and 16S T antigen forms could be distinguished under these sedimentation and fractionation conditions is seen in (d). Peaktubes of5S, 18S, and 28S rRNA markers run in parallelwere 3, 6, and 9, respectively.
in reticulocyte lysates. This led us to question whetherornotthere is arequirement for
ongo-ing DNA replication in order for the oligomeric T antigen to form. To test this, extracts of [35S]methionine-labeled cellswereprepared 10h postinfection, at a time considered to precede
the onset of virus-induced cellular and viral DNA replication. After centrifugation ofthese earlyextractsand anti-Timmunoprecipitation of gradient fractions, we observed no detectable difference in the relative proportions of16Sand 5to7S forms when compared with thoseof the
10 VOL.42, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
460 GIDONI ET AL.
I
I,
IIi %I, i*
b-o
Top
X
II
II
I It
'O
'I
tb~
IQ,IXe Bottom
10,s D0
zL0--2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 2u
FRACTION NUMBER
FIG. 3. Effect ofcytosinearabinosideonTantigensedimentation. Cultures of BSC-1 cells to whichcytosine arabinoside(20Fg/ml)had been addedatthe time of infection withSV40werelabeled with[35S]methionine42to 44hpostinfection,extracted andcentrifugedthroughsucrosegradientsasdescribedin the text and in thelegend to Fig. 1. Anti-Timmunoprecipitates of 20gradientfractions weresubjectedto PAGE andautoradiography, using Kodak XR-5 film.Eachslot,representingtheimmunoprecipitateofagradientfraction,wasscannedwitha Gilforddensitometer,andareasundercurvesrepresenting largeTantigen gelbandswerecut out andweighed.In parallel cultures treatedwithcytosinearabinosideasin thisexperiment,DNAsynthesiswasinhibitedby95%. 5S and 18S rRNA marker peaktubeswere4 and10,respectively. Symbols:0,culture treated with cytosine arabinoside; 0,control untreated culture.
typical 44-h late extract. However, as thecells
werenotsynchronized and therefore theremay
havebeen someDNAreplication induced even
at10 hpostinfection, several inhibitors ofDNA synthesis were used to further ensure the ab-senceof viral and cellular DNAreplication. All compounds tested, including hydroxyurea,
2'-deoxy-2'-azidocytidine (1), aphidicolin, and cytosine arabinoside yieldednoreduction in the relativeamountof labeled 16S T antigen, under conditions where DNA synthesis was reduced
by 90to95%.Indeed, inmany casestherewas a
significant enhancement of the 16S form of T antigen. Aparticularly dramatic example of this increase isseenin Fig. 3. One possible
interpre-tationoftheseresults is that T antigen oligomer-ization may, in fact, be an alternative to its
functioningin DNA replication.
Sensitivity of 16S T antigen form to increased ionic strength. Toelucidate the properties ofthe bonds forming the oligomers of T antigen, we
tested different extraction and centrifugation procedures. Pulse-labeling and pulse-chase
ex-periments showed that the 5 to 7S form is
labeled beforethe16Sform(datanotshown),as
has been reported by others (5, 6). Therefore,
the relative proportions of [35S]methionine-la-beled T antigen 5to7S and16Sformschanged withthe duration of the labeling period, consis-tentwiththeobservation thatmostofthe
accu-mulated T antigen measured by the
enzyme-linked immunosorbent assay exists in rapidly sedimenting complexes. Under similarlabeling,
extraction, and centrifugation conditionsas
de-scribedforFig. 1, the relativeamountof the16S
form wasnot appreciably affected by the pres-ence or absence of0.5% Nonidet P-40, 5 mM MgCl2 or CaCl2, or 5 mM dithiothreitol or
,B-mercaptoethanol. Moreover,the amountofthe
16Sform wasnotdiminishedwhencellextracts
were incubated with quantities of micrococcal
nuclease sufficient to digest 40 ,ug of rRNA markers. Thisindicates that the T antigen in the
16Sform isnot associated with long, nuclease-sensitive stretchesof nucleic acid.
However,sometreatmentdidaffectthe integ-rity of the 16S form. When extractsof infected cells were adjusted to 1.0 M NaCl and
centri-20 19~ -18 x
E
o 17
.17
12
-i12
CZ
D 10
_cc.)_
u0
z
<
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.100.408.52.334.2]DNA BINDING OF SV40 T ANTIGEN OLIGOMERS
fuged through sucrose gradients containing this salt concentration, the oligomeric forms of T antigen vanished (Fig. 4). Indeed, the 5 to7ST antigen form was also seen to sediment even moreslowly than normally observed, suggesting that this form may itself beheterogeneous, per-haps consisting of monomers and dimers, the latter beingsimilarlydissociated at 1.0 MNaCI.
Thus, T antigen oligomers are unstable at higher salt concentrations and must be generated through bonds sensitive to increase in ionic strength. The 45,000-molecular-weight protein tentativelyidentified as VP-1 is also apparently sensitive to the increased saltconcentrations.In
addition,whenextracts were adjusted to pH 6.0 andcentrifugedthrough sucrose gradients at this
pH,there was a considerablereduction but not
an entire disappearance of the amount of the
[35S]methionine-labeled
16S Tantigen
form(data not shown). This pH effect was not ob-served above pH 6.5.
Stability
of 5 to 7S and16S T antigen forms invitro. Several lines of evidence point to the
possibility that SV40 T antigen is a
multifunc-tionalprotein whose various biological activities may reflect different domains or forms of the molecule. As the protein exhibits heterogeneity
with respect to its sedimentation properties, it was of interest to test whether these forms differed with respect to their biological and
2 4 6 8
go ww ,
-
T-biochemicalproperties. To assess any such
dif-ferences,assurance was sought that these forms were notinterconvertible in vitro. For example, as oligomerization of many proteins can be a
concentration-dependentevent, it might be sus-pected thatdilution of the 16S form may cause itsreconversiontothe 5 to 7S form. To test this
possibility,aswell as to test the stability of the 5 to7S form, extracts ofSV40-infected cellswere
subjected to sucrose gradientcentrifugation as
describedinFig. 1. The peak fractions of the 5 to 7S and 16Sformswerethendiluted twofold and threefold, respectively (to dilute the sucrose
concentrationtobelow5%),and each was sepa-rately subjected to a second centrifugation
through sucrose gradients(Fig. 5). Both5 to7S and 16S T antigen forms, even after dilution,
retained theiroriginalsedimentation properties;
5 to 7S T antigen remained as such, whereas virtually no 16S Tantigen was converted to the more slowly sedimentingforms. Nor did there appear to be any conversion ofthe 5 to 7S to
16S form in vitro: coincubation of isolated
[35S]methionine-labeled
5 to 7S peak fractionwith an unlabeled 16S peakfraction did not yield any labeled 5 to 7S T antigen forming heavier
complexes upon re-centrifugation (data not
shown).
DNA-bindingproperties of different
sediment-ing
forms
of T antigen. Many ofthe roles ofT10 12 14 16 18 M
U
94 6845
do 30
7 el-.-.
..1...;.i.
[image:6.505.135.392.401.598.2]4_-12
FIG. 4. SedimentationofSV40Tantigenathigh ionic strength.Extracts of[35S]methionine-labeled infected cellswerepreparedasdescribed in thetext.Afteradjustingthe concentration ofNaClin the extract to 1.0 M, 200 p. wascentrifugedthrough5to20%sucrosegradientsin buffer containing 1.0 MNaCl,0.02 M HEPES buffer, and 0.001 MMgC12 (pH7.4).Nonidet P-40wasomitted fromgradientsolutions due to co-precipitation with 1.0 M salt in the20%osucrose-containingsolution. However, the absence of Nonidet P-40 from sucrose gradients containing0.15 MNaCl didnotmeasurablyaffect theproportionsof 5 to7Sand16ST antigen forms (see the text). Thepeaktubes of 5to7S and16S Tantigenformsruninparallelunder identical conditions as in Fig. 1 were 4and8,respectively.
VOL.42, 1982 461
on November 10, 2019 by guest
http://jvi.asm.org/
462 GIDONI ET AL.J.Vo.
&~~~~~~~~~~~~~~~~~~~~~~~~~~~...
b ~~~~~~~~~~~~~~~~...2.
+
~~DNA.
Two DNA-binding assays wereem-ployed,
both ofwhichtestfortight
binding ofTantigen
toDNA. In thefirstassay,specific
andT
nonspecific
bindingwasexaminedbytestingthedifferential salt-sensitive bindingofT antigento
SV40
origin-containing
(SV40multiorigin
vari-ant[MOVIand
origin-minus
(calfthymus)DNA linkedto cellulose (23). When thismethod wasdeveloped,
itwas found that therearedifferentclasses ofT
antigen
whichexhibitvarying
affini-ty forDNA. To examine whether these classesmight
be related to differentsedimenting
formsofTantigen, the
peak
tubes ofthe 5 to 7S and16S form werecollected after sucrose
gradient
sedimentation, and each bound to and eluted from calf
thymus
orSV40MOVDNA-cellulose columns(Fig.
6). Ithas beenpreviously
shown that the amount of DNA onthe calfthymus orMOV DNA-cellulosecolumns isinconsiderable.
excess ofthe bindable T
antigen
isolated fromcomparable
quantities
of infectedcells (23, 29).Under the binding (pH 7.0; 0.15 M
NaCI)
and elution (pH 7.0; 0.4 M and 1.0 MNaCl)condi-tions used,itwasfound that the 5to7S formof
[image:7.505.60.245.61.338.2]large T antigen binds calf thymus and MOV
FIG. 5. Re-sedimentation of5Sand 16S Tantigen
forms. Sucrose gradient sedimentation of
[35S]methi-onine-labeledinfectedcellextracts was performedas
described in thelegendtoFig.1.Peak tubesof5 to75 and16STantigenforms(generallytubesnos.4and10) were diluted twofold and threefold, respectively, in HIP buffer and subjected to sedimentation through
sucrose gradients as before. Two parallel sucrose gradientsof diluted 55and16Speak tubeswererun, andthe corresponding fractions of the twogradients werepooledandimmunoprecipitated. (a) Re-sedimen-tation of 5to7S Tantigenpeak tube; (b) re-sedimenta-tion of 16S T antigen peak tube. Inparallel sucrose gradientsedimentationofrRNAmarkers,55, 185,and 28SrRNApeaktubeswere nos.4, 11,and16,
respec-tively,asindicatedbyarrows.
ci O c d e f
;,.M..gm
.: ... :.
-M.:....
T...- .:
g h i 1kI
4 O
antigen
in the lytic cycleimply
its interactionwith DNA,
specifically
with the viral origin ofreplication. Theabilityof Tantigento specifical-ly bind DNA sequences at the viral origin in vitro is well documented through several ap-proaches (34, 41). Ithaspreviouslybeen shown that there are different subsets of T antigen molecules which vary in their affinity for DNA (8, 23, 29).The existence of Tantigenastwoor
more
sedimenting
forms raised the question astowhether these forms may differ in their affini-ties for DNA and thus may be related to the DNA-binding heterogeneityof Tantigen.As the 5to75and 165 classes, when separately
isolat-ed,retained theirabilitytosedimentaslightand heavy forms, respectively (Fig. 5), itwas possi-ble to test their individual interactions with
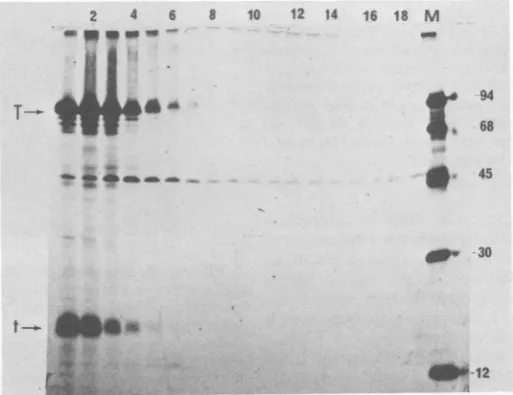
FIG. 6. Binding of55 and165Tantigenformns to calfthymus and MOV DNA-cellulose.
-Five
sucrose gradientsof200-pzl
samplesof[35S]methionine-labeled
infectedcellextractswere runinparallel.PeakS5to7S
(tube no. 4) and 16S (tube no. 10) fractions were pooled and adjusted to pH 7.0, and each poolwas
dividedintotwoportions. DNAbindingof 5to7S(a through f0 and 16S (g through 1) T antigen to calf
thymusDNA-cellulose(a, b,c,g,h, i)orF-161MOV DNA(d,e,f,j, k, 1)wastestedby bindingand elution
pH7.0inbuffercontaining0.1 MNACI (a, d, g,j), followedbyelution inbuffercontaining0.4 MNaCl(b,
e,h,k),then inbuffercontaining1.0 MNaCl(c, f,i, 1).
Allelution fractions wereadjustedto0.1 MNaClby dilution, immunoprecipitated with anti-T serum, and
analyzedbyPAGE andautoradiography.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.272.439.366.523.2]DNA BINDING OF SV40T ANTIGEN OLIGOMERS 463
DNA, whereas small t antigen does not. The 5 to 7S T antigen, which bound to DNA, could be eluted from calf thymus DNA with 0.4 M
NaCI,
butrequired higher ionic strength (1.0 M NaCI) tobe eluted from MOV DNA, consistent with earlierobservations of whole cell extracts ofT-antigen-containing infected cells (23). In con-trast, the 16S form bound very inefficiently to
calfthymus DNA and even more poorly to MOV DNA, in thatconsiderably more of this T anti-genclass did not bind to the column at all when compared to the proportions of bound and non-bound 5 to 7S T antigen. In contrast to the 5 to
7Sform,nodetectable tight (1.0 M
NaCl-sensi-tive) binding of the 16S form to MOV DNA was
observed.Thus, these two forms clearly differ in their interaction with DNA immobilized on cel-lulose columns.
We also tested the binding of T antigen to
various DNA sequences in an assay in which extractsfrominfected cellscontainingTantigen areboundto32P-labeledrestriction fragments of viral SV40 DNA in solution (19; Scheller et al.,
submitted forpublication). The T antigen-DNA
fragment
complex is then immunoprecipitatedwithanti-SV40 T serum(Schelleret al.,
submit-tedfor publication).Incubation of BstNl-digest-edSV40 DNA comprising 13 fragments,
includ-ing the 311-base-pair G fragment spanning the viral origin ofreplication, with extracts of
SV40-infectedcellsundersuitable binding conditions,
yieldedconsiderablequantities of the origin
se-quence-containing G fragment specifically bound to Tantigen.Totestwhether the different
sedimenting forms ofT antigen varied intheir
ability to specifically bind to origins, gradient
fractions were bound to 32P-labeled BstNl-di-gestedSV40
fragments
in solution(Fig. 7).This experiment differed from the column-binding experiments in two major ways: first,
the kinetics of binding in solution were more rapidthanwhenDNAwasimmobilizedon
cellu-lose; second, whereas the column-binding
ex-periment
provided
information about labeled (newlysynthesized)
Tantigen
molecules, thesolution-binding experiment measured the
spe-cific-origin-binding activity
ofthe totalaccumu-lated T antigen in each
gradient
fraction. Theresultsconfirmed and extendedthose obtained
with the DNA-cellulose binding assay.
Only
fractions in the 5 to 7S region of the gradient boundtoBstN1 fragment
G;
morerapidly
sedi-menting forms exhibited little or no
specific
affinity for the
origin-containing fragment.
It isnoteworthythattubeno.4showed
considerably
greater Gfragmentbinding
than did tubeno. 3.Consistentwith theresults of the NaCl dissocia-tion experiment
(Fig. 3),
whichsuggested
that the5 to7S classmaybeheterogeneous,it can besuggestedthat the 5to7ST
antigen
may consistof both monomers and dimers, of which the latter are the major origin-binding form.
DISCUSSION
Our experiments indicate that different forms
of T antigen vary in their affinity for DNA. Under the DNA-binding conditions that we used, the 5 to 7S T antigen form could bind to DNA with high affinity, whereas the 16S form could not. These data are somewhat at odds with those of Myers et al. (21), who provideevidence from electron microscopic analysis that prepara-tions ofpurified and concentrated D-2 protein
AM 2 4 6 8 10 12
I _
T 4..
lA IS 18 20
_
M2n5e , _
B M 2 3 4 5 -0 o 9C ! mv a 35 CO M
A-
D- C--
F-FIG. 7. Bindingof varioussedimenting forms ofT antigento DNAfragments in solution. Twoparallel
sucrose gradients of
200-1.l
portions of[35S]methio-nine-labeled and unlabeled infected cell extractswere run.Gradient fractionsof[35S]methionine-labeled ex-tractswerecollected andsubjectedto immunoprecipi-tation and analysisas described in thelegendtoFig.
1(A),orgradientfractions of unlabeledextract were incubated with 5 ng of32P-labeled BstNlSV40 restric-tionfragments, followed by immunoprecipitation of the Tantigen DNAcomplexas described in thetext (B). Tubes from fractions 6 andupwardsweredoubled in theDNA-bindingassay, asshown in thefigure,to
compensate for the greaterquantities of
[35S]methio-nine-labeled T antigen present in the more slowly
sedimenting fractions.
VOL.42, 1982
I ca
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.505.265.459.251.530.2]464 GIDONI ET AL.
consist mainly of oligomers which were
ob-served boundtosequencesattheviral origin of replication. To reconcile theirobservationswith
the binding studies reported herein, it can be
suggested that either highly purifiedD-2protein isfunctionally different from 16S T antigen
iso-lated from extracts of SV40-infected cells or
that, although the active binding form of T
antigen is the 5to7S form, the 16S oligomersare
generated through DNA binding. Possibly, T
antigen binds to DNA as monomer or dimer,
whichcould notbeseparated in these sedimen-tation conditions, and then forms tetramers
throughjuxtaposition at its binding sequences.
Although cessation of DNA replication did not
reduce the appearance of 16S T antigen, it is
possible that these oligomers are formed by
binding to cellularornonreplicating viral
chro-matin. Experiments are under way to test
whetherT antigen oligomers can be generated
through bindingtoDNA invitro.
Another area of investigation concerns the nature ofthe 16S oligomers. As the monomer
molecular weight ofTantigen is 82,000(7), the 5
to7Sformprobably consists ofmonomers and possibly dimers, whereas the 16S form, which sediments somewhat more slowly than native
thyroglobulin (molecular weight, 670,000), may
consistofoligomers ranging from tetramers to octamers.Ithas beenreportedthat themajority
of D-2 T antigen present in highly purified, concentrated protein preparations existsas
tet-ramers, as estimated by electron microscopic
analysis (21). Again, it is difficult toassess the
extent towhichpurifiedD-2tetramersresemble
the 16S T antigen observed in crude extracts.
Although it is possible that Tantigen
oligomer-ization in infected cells maybe the result ofits
complexing with other viralorcellularproteins, noevidenceofany specifically
immunoprecipi-tated [35S]methionine-labeled protein associated
with16STantigenwasobserved. It is alsonot inconceivablethatsome16STantigen is, in fact,
a protein-DNA complex. Our experiments do not rule out the existence of short, protected
sequencesof nucleic acid.Experimentsto
deter-minewhether thereareotherproteinsornucleic
acidfragmentsassociatedwith16STantigenare
inprogress.
SV40Tantigenhas beenshowntobindto(6,
9, 14, 18, 20) and stabilize (22)an
immunopre-cipitation-resistant complex with a
53,000-mo-lecular-weighthostprotein foundin transformed
cells. This complex was shown to sediment
morerapidlythan the 16STantigen form(18,20,
28). Althoughit has been suggestedthat 16ST antigenmaybeanintermediatein the formation
of the22S T-53,000-molecular-weight host
pro-teincomplex, thereare substantialquantitiesof
16S T antigen in lytically infected cells which
have considerably smaller amounts of the
53,000-molecular-weight
protein (10, 20).
Theproperties of theTantigenthatis
complexed
tothe
53,000-molecular-weight
hostprotein
aredif-ferent fromthose ofthe 16S form because the
formercanbindtoDNA sequencesatthe viral
origin (Scheller et al., submitted for
publica-tion), whereas the latter doesnot.The existence ofmultipleforms ofT
antigen
differing in their state of
oligomerization
has beenreported by several othergroups.Although
minordiscrepanciesamongsomeofthereported
studies exist, salientfeaturesofthe
phenomena
canbesummarized asfollows.(i)
Inlytic
infec-tion the Tantigenoligomers sedimentat approx-imately 16S, whereas intransformed cellsthere are still heavier 22S T antigencomplexes
con-taining the
53,000-molecular-weight
nonviral T antigen. (ii) Pulse-labeling studies suggest that themonomericor5to7S form isformed beforethe heavier forms and therefore may be their precursor. (iii) tsA mutant T antigens do not
form oligomers at the restrictive temperature.
(iv) Therapidly sedimenting T
antigen
is moreextensively
phosphorylated
than themore slow-ly sedimenting forms. Inexperiments
not de-scribed in this paper we have confirmed all of these observations except for thefourth,
in which wedidnotobserveasignificant
differencein the degree of
phosphorylation
of the twosedimenting forms ofT antigen. However, our
dataareconsistentwith the
pulse-labeling
stud-ies in thatwehavepreviouslyshown thatnewly synthesizedTantigen bindsmoreefficiently (23,
28) and tightly (23) to DNA than do older T antigen molecules. We are currentlyexploring
thepossibilitythatvariousmonoclonalantibod-ies whichreactwithsubpopulationsofTantigen
thatare active in DNAbinding (Schelleretal.) preferentially immunoprecipitate the 5 to 7S form.
Biological andbiochemical studies of SV40T antigen suggestthat this viral geneproductisa
multifunctional protein. Although the different functions may be due to different domains or
post-transcriptionally modified states (e.g.,
phosphorylation)of the T antigenpolypeptide,it is also possible that T antigen may exhibit het-erogeneity through various degrees of oligomer-ization. Theobservation that the 5 to 7S and 16S forms differ in their DNA-binding properties may be related to different functions of the protein. We can speculate that, although the newlysynthesized 5 to 7S T antigen form is the active viralorigin-binding form, the subsequent-ly formed more rapidsubsequent-ly sedimenting T antigen
oligomers may (i) bind nonspecifically to DNA and play a role in the cellular DNA replication function of Tantigen; (ii) have a function unre-latedto DNAbinding, such as association with
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
DNA BINDING OF SV40 T ANTIGEN OLIGOMERS 465
the53,000-molecular-weight nonviral T antigen,
or adenosine triphosphatase activity; or (iii) be
inactive forms of T antigen representing an
additional mode of autoregulation of the
avail-ability of functionalT antigen.Weare currently
carryingoutexperimentsto testtheseand other
possibilities.
ACKNOWLEDGMENTS
Studies carriedoutin Israelwerefundedbyagrantfrom the
U.S.-Israel BinationalFund to C. P.;those carried outin the
United Statesweresupported byU.S. PublicHealth Service grantCA 26905toC.P.
R. Pollack and Y. Gronerarethanked for critical reviewof
this manuscript.
ADDENDUMINPROOF
While this paper was submitted for publication, Bradleyetal.(M.K. Bradley,J. D.Griffin,andD. M.
Livingston, Cell 28:125-134, 1982) reported that the
slowly sedimentingform of Tantigencanberesolved
into 5.5Sand7Sspecies. Theyhave reportedthat of
thetwoforms onlythe7S speciesis activeinbinding toDNA.
LITERATURECITED
1. Bjursell,G. 1978. Effectsof2'-deoxy-2'-azidocytidineon
polyomavirus DNAreplication: evidence forrolling
cir-cle-typemechanism. J.Virol. 26:136-142.
2. Brugge,J.,andJ.Butel. 1975.Involvementof the simian
virus 40geneA function in the maintenanceof
transfor-mation. J.Virol. 15:619-635.
3. Carroll, R.B.,L.Hager, and R. Dulbecco. 1974. Simian
virus 40TantigenbindstoDNA. Proc. Natl.Acad. Sci.
U.S.A.71:3754-3757.
4. Cowan, K., P. Tegtmeyer, and D. D. Anthony. 1973.
Relationship of replication and transcription of simian
virus 40 DNA. Proc. Natl. Acad. Sci. U.S.A.
70:1927-1930.
5. Fanning, E., B.Nowak, and C.Burger. 1981.Detection
and characterizationofmultipleforms of simianvirus 40
largeTantigen. J. Virol.37:92-102.
6. Greenspan, D. S.,and R. B. Carroll. 1981.Complex of
simian virus 40 large tumour antigen and 48000-dalton
host tumourantigen. Proc.Natl.Acad. Sci. USA
78:105-109.
7. Griffin, J. D., S. Light, and D. M. Livigston. 1978.
Measurements ofthe molecularsize of the simianvirus 40
largeTantigen.J. Virol. 27:218-226.
8. Griff,J. D., G. Spangler,and D. M. LivIngton.1979.
Proteinkinaseactivityassociatedwith simian virus40T
antigen. Proc.Natl. Acad. Sci. U.S.A. 76:2610-2614.
9. Gurney, E. G., R. 0. Harrison, and J. Fenno. 1980.
Monoclonalantibodiesagainstsimianvirus 40 Tantigens: evidencefor distinct subclassesoflargeTantigenandfor
similarities amongnonviralTantigens. J. Virol.
34:752-763.
10. Harlow, E., D. C. Pim, and L. V. Crawford. 1981.
Complex of simian virus 40 large T antigen and host
53,000 molecular-weight proteininmonkeycells. J.Virol.
37:564-573.
11. Kilton,L.J.,M.Bradley,C.Mehta,and D. M. Livingston.
1981.Rapidand sensitivequantitative immunoassay for
thelargesimianvirus40 Tantigen.J. Virol.38:612-620.
12. Khmura,B.,and R.Dulbecco.1973. A temperature-sensi-tive mutant of simian virus 40 affecting transforming
ability.Virology52:529-534.
13. Kuchino,T.,andN.Yamaguchi.1975.Characterizationof
Tantigenin cells infectedwith atemperature-sensitive mutant of simian virus40. J.Virol. 15:1302-1307.
14. Lane,D.P.,andL. V.Crawford.1979. Tantigenis bound
toahostproteinin SV40-transformedcells.Nature
(Lon-don)278:261-263.
15. Livingston,D.M.,I.C.Henderson,andJ.Hudson.1974. SV40Tantigen:partial purificationandproperties.Cold SpringHarborSymp. Quant.Biol.39:283-289.
16. Mardn, R. G., and J.Y. Chou. 1975. Simianvirus 40 functionsrequiredfortheestablishment andmaintenance ofmalignanttransformation.J.Virol.15:599-612.
17. Mardn,R.G.,V.P.Setlow,A. B.
Chepelisky,
R.Seif,A.M.Lewis, Jr.,C. D.Scher,C.D.Sties,andJ.Avila. 1979. Rolesof the Tantigensintransformationby SV40.Cold SpringHarborSymp.Quant.Biol. 44:311-324. 18. McCormick, F.,and E.Harlow. 1980. Association ofa
murine53,000-daltonphosphoproteinwith simian virus 40
largeTantigenin transformedcells. J. Virol.34:213-224. 19. McKay,R.1981.Bindingofasimian virus 40 Tantigen
relatedproteintoDNA. J.Mol. Biol. 145:471-488. 20. Melero,J.A.,D.T.Stltt,W. F.Mangel,and R.B.Carroll.
1979.Identification ofnewpolypeptide species(48-54K)
immunoprecipitatedbyantiserumtopurifiedlargeT anti-genandpresentin SV40infectedand transformedcells. Virology93:466-480.
21. Myers, R. M., R. C. Williams, and R. Tjian. 1981.
Oligomeric structureofan SV40T antigenin freeform
andasboundto SV40DNA. J. Mol.Biol. 148:347-353. 22. Oren, M., W.Maltzman,and A. J.Levine. 1981.
Post-translationalregulationofthe 54K cellulartumourantigen
in normaland transformedcells. Mol. Cell Biol. 1:101-110.
23. Oren, M.,E.Winocour, andC.Prives. 1980.Differential affinities ofSV40T antigenfor DNA. Proc. Natl. Acad. Sci. U.S.A. 77:220-224.
24. Osborn,M.,and K.Weber.1975. SV40:Tantigen,theA
function andtransformation. ColdSpringHarborSymp.
Quant.Biol. 39:267-276.
25. Osborn,M.,andK. Weber.1975.Simianvirus 40geneA function and maintenance of transformation. J. Virol. 15:636-644.
26. Ozer, H. L., and P. Tegtmeyer. 1972. Synthesis and
assemblyofSV40 H.Synthesisofthemajorcapsid protein
and itsincorporationintoviralparticles.J.Virol.9:52-60. 27. Potter,C.W.,B. C.McLaughlin,andJ.S. Oxford.1969. Simian virus40-inducedTandtumorantigens. J. Virol. 14:574-579.
28. Prives,C.,Y.Beck,D.Gidoni,M.Oren,and H.Shure.
1979. DNAbindingandsedimentationpropertiesofSV40
tumour antigenssynthesized invivo and in vitro. Cold
SpringHarborSymp.Quant.Biol. 44:123-130. 29. Prives, C., Y. Beck, and H. Shure. 1980. DNA binding
properties of simianvirus 40 T-antigens synthesized in vivoand in vitro. J. Virol.33:689-696.
30. Prives,C.L.,and H.Shure.1979.Cell-freetranslationof simian virus 40 16S and 19S L-strand-specific mRNA
classes tomajor VP-1 and minor VP2 and VP3 capsid proteins.J.Virol.29:1204-1212.
31. Reed,S.I.,G. R.Stark,andJ.C.Alwine.1976. Autoregu-lationofsimian virus40 gene AbyTantigen.Proc.Natl.
Acad.Sci. U.S.A. 73:3038-3087.
32. Rigby,P. W.J.,M.Deckman,C. R.Rhodes,and P.Berg.
1977. Labelling deoxyribonucleic acid to high specific
activityin vitrobynicktranslationwith DNApolymerase.
J. Mol. Biol. 113:237-251.
33.
Rundeil,
K.,J.K.Collins,P.Tegtmeyer,H. L.Ozer,C-i. Lai, and D. Nathans. Identificationof simian virus 40 proteinA.J.Virol. 21:636-646.34. Shalloway, D., T.Kleinberger, D. M. Livingston. 1980. MappingofSV40DNAreplicationorigin region binding
sites for theSV40Tantigen byprotectionagainst
exonu-clease IIIdigestion.Cell20:411-422.
35. Smith, A.E.,R.Smith,andE.Paucha. Extractionand
fingerprint analysisofsimian virus 40largeandsmall
T-antigens.J.Virol.28:140-153.
36. Tegtmeyer,P.1972.Simianvirus40deoxyribonucleicacid synthesis:the viralreplicon.J.Virol.10:591-598. VOL.42, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
466 GIDONI ET AL.
37. Tegtmeyer,P.1975.Function ofsimian virus 40geneAin
transforming infection. J. Virol. 15:613-618.
38. Tegtmeyer,P., K.Rundell,J.K.Collins.1974. Modifica-tion of simian virus 40 protein A. J. Virol. 21;647-657. 39. Tegtmeyer,P., M. Schwartz, J. K. Collins, and K. Rundell.
1975. Regulation oftumourantigensynthesis by simian virus40geneA. J.Virol. 16:168-178.
40. Tjhan, R. 1978. The binding siteonSV40 DNA foraT
antigen-related protein. Cell 13:165-179.
41. T3ian, R., and R. Robbins. 1979. Enzymatic activities
associated with a purified simian virus 40 T antigen-related protein. Proc. Natl. Acad. Sci. U.S.A. 76:610-614.
42. Tooze, J. (ed.). 1980. DNA tumor viruses: molecular biology oftumourviruses, 2nded.,part2. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y. 43. Voller, A., A.Bartlett,D. E. Bidwell,M. F.Clark,and
A. N. Adams.1976.The detection of viruses by
enzyme-linked immunosorbent assay (ELISA). J. Gen. Virol.
33:165-167.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/


![FIG. 5.forms.onine-labeleddescribedandweregradientsHIPandsucroseweretationtiongradient28Stively, Re-sedimentation of 5S and 16S T antigen Sucrose gradient sedimentation of [35S]methi- infected cell extracts was performed as in the legend to Fig](https://thumb-us.123doks.com/thumbv2/123dok_us/1460538.98732/7.505.272.439.366.523/labeleddescribedandweregradientshipandsucroseweretationtiongradient-stively-sedimentation-sucrose-sedimentation-infected-extracts-performed.webp)

