City, University of London Institutional Repository
Citation:
Friston, K. J., Bastos, A. M., Pinotsis, D. A. & Litvak, V. (2014). LFP and
oscillations - what do they tell us?. Current Opinion in Neurobiology, 31, pp. 1-6. doi:
10.1016/j.conb.2014.05.004
This is the accepted version of the paper.
This version of the publication may differ from the final published
version.
Permanent repository link:
http://openaccess.city.ac.uk/19427/
Link to published version:
http://dx.doi.org/10.1016/j.conb.2014.05.004
Copyright and reuse: City Research Online aims to make research
outputs of City, University of London available to a wider audience.
Copyright and Moral Rights remain with the author(s) and/or copyright
holders. URLs from City Research Online may be freely distributed and
linked to.
City Research Online: http://openaccess.city.ac.uk/ [email protected]
LFP
and
oscillations—what
do
they
tell
us?
Karl
J
Friston
1,
Andre´ M
Bastos
2,3,
Dimitris
Pinotsis
1and
Vladimir
Litvak
1Thisreviewsurveysrecenttrendsintheuseoflocalfield potentials—andtheirnon-invasivecounterparts—toaddress theprinciplesoffunctionalbrainarchitectures.Inparticular,we treatoscillationsasthe(observable)signatureof context-sensitivechangesinsynapticefficacythatunderliecoordinated dynamicsandmessage-passinginthebrain.Thisrichsourceof informationisnowbeingexploitedbyvariousprocedures—like dynamiccausalmodelling—totesthypothesesaboutneuronal circuitsinhealthanddisease.Furthermore,therolesplayedby neuromodulatorymechanismscanbeaddresseddirectly throughtheireffectsonoscillatoryphenomena.These neuromodulatoryorgaincontrolprocessesarecentraltomany theoriesofnormalbrainfunction(e.g.attention)andthe pathophysiologyofseveralneuropsychiatricconditions(e.g. Parkinson’sdisease).
Addresses
1TheWellcomeTrustCentreforNeuroimaging,UniversityCollege
London,QueenSquare,LondonWC1N3BG,UK
2
CenterforNeuroscienceandCenterforMindandBrain,Universityof California-Davis,Davis,CA95618,USA
3ErnstStru¨ngmannInstituteinCooperationwithMaxPlanckSociety,
Deutschordenstraße46,60528Frankfurt,Germany
Correspondingauthor:Friston,KarlJ([email protected])
CurrentOpinioninNeurobiology2015,31:1–6
ThisreviewcomesfromathemedissueonBrainrhythmsand dynamiccoordination
EditedbyGyo¨rgyBuzsa´kiandWalterFreeman
ForacompleteoverviewseetheIssueandtheEditorial
Availableonline30thJuly2014
http://dx.doi.org/10.1016/j.conb.2014.05.004
0959-4388/#2014TheAuthors.PublishedbyElsevierLtd.Thisisan openaccessarticleundertheCCBYlicense(http://creativecommons. org/licenses/by/3.0/).
Introduction
Our reviewcomprisesfour sections:the firstconsiders the centralroleofgain controlinhierarchicalmessage passingandpredictivecoding;withaspecialemphasis on precision, attention and sensory attenuation. The second treats oscillations and local field potentials as fingerprints that reveal asymmetries in forward and backward extrinsic connections incorticalhierarchies. Thisis aprescient areaofresearch,because ithas the potential to disclose the hierarchical connectome and the putative predictive coding it supports. The third section looksmorecloselyathorizontalconnectionsin visual cortex andhow local fieldpotentials have been
used to characterise context-sensitive changes in lat-eral interactions—in terms of effective connectivity and its underlying (GABAergic)synaptic gain control. Finally, we consideran example ofcouplingbetween cortical and subcortical systems that speaks to the use of oscillations in characterising pathophysiology. Specifically, we look at pathological beta oscillations
and their dynamic causal modelling in Parkinson’s
disease.
Oscillations,
precision
and
predictive
coding
Our treatment of oscillations rests on the premise thatdynamic coordination can be understood in terms of
predictivecoding[1–3].Predictivecodingsupposesthat thebrain isastatistical organ,generatingpredictionsor hypothesesaboutthestateoftheworld—predictionsthat aretestedagainstsensoryevidence.This(Bayesianbrain) perspective ispotentially importantbecausemany neu-ropsychiatricsyndromes(rangingfromautismto psycho-sis)canbecastintermsoffalseinferenceaboutstatesof the world (or the body) that may be due to aberrant neuromodulation or gain control at the synaptic level [4,5].
The circumstantial evidence for predictive coding is substantial—both in terms of the anatomy of extrinsic (between-areas) and intrinsic (within-area) connections and thephysiologyof synapticinteractions[3].Inthese schemes,top-down predictionsareusedto form predic-tion errorsateachlevelof corticaland subcortical hier-archies. The prediction errorsare then returnedto the levelabovetoupdatepredictionsinaBayesiansense.In brief,thepredictionerrorsreportthe‘newsworthy’ infor-mationfrom alowerhierarchical levelthatwasnot pre-dictedbythehigherlevel.Acrucialaspectofthismessage passing is the selection of ascending information by adjusting the‘volume’ or gainof prediction errors that compete for influenceover higherlevels of processing. Functionally, thisgaincorresponds totheexpected pre-cision(inversevarianceor signal-to-noise ratio)that sets theconfidenceaffordedtopredictionerrors. Psychologi-cally this hasbeen proposedas thebasis of attentional gain [6]. Physiologically, precision corresponds to the postsynapticgainor sensitivityof cellsreporting predic-tion errors(currently thoughtto belarge principalcells that send extrinsic efferents of a forward type, such as superficial pyramidal cells in cortex). Thisis important becausethesynapticgainorefficacyofcoupledneuronal
(oscillatory) behaviour. See Figure 1.Because, synchro-nousactivitydeterminessynapticgain[7],oscillationshave amechanisticimpactonneuronalprocessing—ratherthan beingepiphenomenal—whichcompletesthecircular caus-alitybetweensynchronyandsynapticefficacy.
Castinghierarchicalneuronalprocessingintermsof pre-dictive coding has proven useful in providing formal modelsofbehaviourandstructure–functionrelationships in thebrain. Itisnowarguably thedominantparadigm in cognitive neuroscience. Under predictive coding, the central role of precision—mediated by classical
neuromodulatory and synchronous gain control—fits
comfortablywithcomputationalandphysiological formu-lationsof neuronal processing. Crucially, electrophysio-logical studies of oscillations provide a rich source of empirical datafor estimatingsynaptic efficacy. In what follows,weconsiderrecentempirical approachesto un-derstanding the functional architectures of predictive coding using local field potentials and dynamic causal models(DCM) oftheirspectralbehaviour[8].
Physiologically, synaptic gain rests on a competition betweenexcitatoryandinhibitoryprocesses.Thismeans
2 Brainrhythmsanddynamiccoordination
Figure1
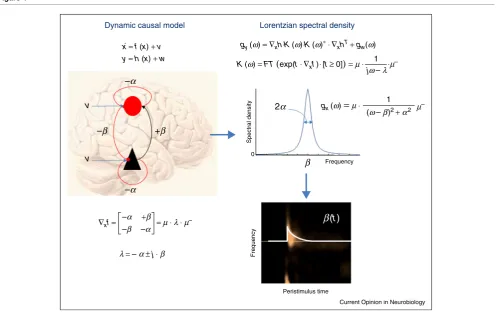
x = f (x) + v y = h (x) + w
Dynamic causal model Lorentzian spectral density
1
gy (ω) = ∇xh⋅K (ω)⋅K (ω)∗ ⋅ ∇xhT + gw(ω)
K (ω) = FT (exp(t ⋅ ∇xf)⋅ [t ≥ 0]) = μ ⋅
jω– λ⋅μ –
1
(ω–β)2 + α2
gx (ω)= μ ⋅ μ–
v v
–β
β
β(t)
2α
+β
–α –α
Frequency 0
Frequency
Spectral density
Peristimulus time = μ ⋅ λ ⋅ μ–
λ = – α ± j ⋅ β +β –α
–α ∇xf =
–β
Current Opinion in Neurobiology
[image:3.612.56.554.91.403.2]that a detailed characterisation of gain control should differentiate betweenexcitatoryandinhibitory postsyn-apticcurrents. However,LFPoscillationsare generated byboth—creatingadifficultinverseproblem.DCMtries to resolve this problem with the Bayesian inversion of physiologically plausible forward models of coupled inhibitory andexcitatorypopulations.
Hierarchical
message
passing
and
the
spectral
connectome
Thissectionfocusesonrecenttrendsinthe characteris-ation of functional integration in cortical hierarchies. Much current work focuses onspectral asymmetries in the(functionalandeffective)connectivitybetween des-cending(top-down)andascending(bottom-up)extrinsic (between area) projections. Bastos et al. have shown canonicalpatternsofdirectedinteractionsbetween differ-entvisualareas, withtheta and gammaoscillations pre-dominating in thebottom-up (orfeedforward) direction andbetaoscillationssignalinthetop-down(orfeedback) direction.Inaddition,thismetricoffunctionalasymmetry betweeninter-arealoscillatoryinteractionspredictedthe underlyinganatomical asymmetriesintermsof laminar-specifictop-downandbottom-upconnections(seeVezoli, Bastos,Fries,thisissueformoredetails).Similarspectral asymmetries in forward and backward message passing have alsobeen found inthe auditorysystem [9].The emergingpictureisthatoscillatorycouplingprescribesa functional cortical hierarchy that closely matches the anatomicalhierarchy[10].Thefunctionofthishierarchy is anopenquestion,althoughpredictivecoding models offeranintriguing explanation—intermsofhierarchical Bayesianinference.
Complementingthis work,Richteret al.(seethisissue) describebetaoscillationsfromextrastriatecortextoV1in themonkeythatpredictthestrengthofevokedpotentials in V1,andmaythereforebeacandidatemechanismfor gain control. Furthermore, the top-down beta predicts stimulus-response mappings that the animal has been trainedto perform.Thus,beta signalsmayprovide top-down influences that contextualize lower-level proces-sing. These findings fit comfortably with predictive coding models[3],whichpredictthattop-down cortico-cortical connectionsconvey prediction signals atslower time scales(e.g. beta) compared to bottom-up connec-tions that convey predictionerror signals at faster time scales(e.g. gamma).Recentwork supportsthis hypoth-esis,linkingfastandslowfrequenciestopredictionerror and predictions,respectively:
Bauer and colleagues [11] have recently demonstrated that thecumulative probability of astimulus change(a
proxy for stimulus predictability) was tracked by
attention-dependent alpha-band oscillations, while the inverse of cumulative probability (a proxyfor surprise)
was tracked by attention-dependent gamma-band
oscillations. This suggests that neuronal signalling of predictions is mediated by alpha and prediction errors by gamma. These are exactly the sort of spectral dis-sociations one would expect if lower frequencies were involvedinrelayingpredictionsandfasterfrequenciesin relayingpredictionerror.
Theseexperimentalfindingsarenowbeingincorporated intomodelsofcanonicalmicrocircuitry[3,12]to under-standatamechanisticlevelhowoscillationscontributeto top-downandbottom-upprocessing.Akeychallengefor futureworkwillbetounderstandnotonlythefunctional segregation between top-down and bottom-up signalling but also the functionalintegration ofthese streams,which maybesubservedbylaminarspecificprocessing within thecorticalmicrocircuit[3,13].
Gain
control
and
lateral
interactions
in
cortex
The precedingsectionsfocusedonthedynamic coordi-nationamong corticalareasas indexedbytheirspectral coupling.Here,wefocusoncorticalgaincontrol(implicit in the optimisation of precision) within the intrinsic connections of the canonical cortical microcircuit. In particular, we look atrecent advances in characterising excitatory-inhibitorybalance—asmediatedbyhorizontal connections withinvisualcortex—using dynamiccausal modellingand neuralfields.Dynamic causal modelling (DCM) is a biophysically
informedBayesianframeworkforcomparinghypotheses or networkmodelsof (neurophysiological)timeseries.It isan establishedprocedurein theanalysisof functional magnetic resonance timeseries [14,15] and isnow used increasinglyforthecharacterisationof electrophysiologi-calmeasurements.Thereisanextensiveliteratureonthe validation of DCM rangingfrom facevalidationstudies [16] to construct validation in terms of multimodal
measurements [17], pharmacological manipulations
[8,18] and psychophysical constructs [19]; for example, predictive coding. Predictive validity has been estab-lished in studies of pathophysiology [20]. Generally, dynamiccausalmodellingusespointsources(cf., equiv-alent current dipoles); however, recent developments nowallow theuseofneuralfieldsintheforwardmodel.
Dynamiccausalmodellingofneuralfieldsandcortical gaincontrol
Neural fields treat neuronal signalling as a continuous process on the cortical sheet using partial differential
equations [21,22]. By combining neural fields with
of receptive fields in visual cortex (V1) had a smaller stimulussummationfield,whilein[27]theyshowedthat thebalance of excitatory-inhibitory influencescouldbe modulatedbystimuluscontext.
Neuronalresponses invisualareasaresensitiveto both stimuluscontrastandtop-down factors[28].This con-text-sensitivity is thought to underlie visual attention [29]. It is also known that gamma band oscillations (30–100Hz)inV1aresensitivetocontrast,stimulussize andattention [30].Furthermore, attentionincreasesthe
peak frequency of gamma oscillations [31]. This
suggests an intimate link among gamma oscillations, stimuluscontrast,horizontalconnectionsandcorticalgain control.
TheserelationshipshavebeenexaminedusingBayesian modelcomparisonofdynamiccausalmodelsthatembody competinghypotheses abouthow visualcontrasteffects lateralinteractions[32].Usinginvasive
electrophysiologi-cal responses from awake-behaving monkeys several
mechanisms were compared [31]: candidate DCMs
allowed for contrast-dependentchangesin thestrength of recurrent local connections[6], thestrength of hori-zontalconnections[33]orthespatialextentofhorizontal connections[27]. The ability of each model to explain inducedresponseswasevaluatedintermsoftheir Baye-sianmodelevidence;whichprovidesaprincipledwayto
evaluate competing hypotheses. Bayesian model
com-parisonsuggested that increasing contrastincreases the sensitivityor gain of superficialpyramidal cells to hori-zontal inputs from spiny stellate populations. This is consistent with precision or gain control in predictive coding—assumingthatincreasingcontrastincreases sig-nal-to-noise. Furthermore, they provide a mechanistic explanationforwhythereceptivefieldsofV1unitsshrink withincreasingcontrast.
Gain
control
and
pathophysiological
oscillations
Weclosewithanimportantexampleofdynamic coordi-nation in pathophysiology. Namely, the emergence of pathologicalbetaoscillationsinParkinson’sdisease(PD) andtheircharacterisationwithintracorticaland non-inva-sivemethods to examinethe underlyingdirected func-tional connectivity (Granger causality) and effective connectivity(dynamiccausal modelling).
PD is associated with degeneration of dopaminergic
neurons in the substantianigra. However, the
mechan-isms mediating Parkinsonian symptoms are not well
understood. One robust finding—in both patients and animalmodels—isincreasedoscillationsinthelowerbeta band (around 18–20Hz) in the basal ganglia (BG), particularlythesubthalamicnucleus(STN).Their ampli-tudecorrelateswithslownessandrigiditybutnottremor [34].Betaoscillationsdecreasewithmovementandtheir
baselinelevelisgreatlyreducedbydopaminergic medi-cation and DeepBrain Stimulation (DBS)[35].Recent studiespointtoacausalroleofbetainmovementslowing; transcranial alternating current stimulation in the beta
band slows voluntary movement in healthy subjects
[36,37]and adaptiveDBStriggeredbyhighbeta power appearsto be superior to constantDBS in ameliorating Parkinsoniansymptoms[38].
Thecentralroleof abnormalbetahasmotivatedafocus on its generative mechanisms. The reciprocally
con-nectedglutamatergicSTN andGABAergic Globus
Pal-lidus(GP)arenaturalcandidatesforgeneratingbeta[39].
A recent simulation study showed that oscillations
emerge with realistic connectivity based on the latest empiricalfindings[40].Inlightofthesesimulations,one mightassumethatbetaoscillationsgeneratedinBGreach thecortexviathethalamusand‘jam’it.However,studies
using simultaneous MEG and STN-LFP recordings in
DBSpatientssuggest that thepathophysiology is more complicated; functional connectivity in the beta band, manifest as cortico-STN coherence, is prominent and involves ipsilateral motor areas of the cortex [41,42]. However,thefrequencyofthiscoherencedoesnotmatch thatofSTNbetaoscillations;ratheritisintheupperbeta band(25–30Hz).Moreover,directedfunctional connec-tivityanalysesshowthatthecortexdrivestheSTNinthis frequencyband[41].Additionalevidencefordissociation between the pathological beta and cortico-STN coher-enceisthefactthatthecoherenceisonlyweaklyaffected
by dopaminergic medication and movement [43]. This
cortico-STN coherence contrasts with the
dopamine-sensitive synchronisation in the lower beta band—evi-dent within and between basal ganglia nuclei, as was recently shown bylooking at different cellpopulations withinoneSTN[44,45]andbetweenbilateralSTN[46].
Dynamiccausalmodellingofbetaoscillationsand synapticgain
Dynamiccausalmodellinghasthepotentialtoreconcile thesefindings andrevealthearchitecturesthatunderlie pathologicaloscillations.TwoDCMstudiesofbeta oscil-lations—oneinaratmodelofPDandoneinpatients— have been published to date [47,48]. Both show an increase in cortical drive to theSTN, accompanied by changesin STN-GP coupling in the pathologicalstate. ThusDCMpointstochangesinsynapticgaincausedby abnormalneuromodulationasthekeymechanism under-lyingpathologicalincreasesinBGbetasynchrony.How thatsynchronyimpairsmovement isstillan open ques-tion—anditmaytranspirethatthemechanism involves BG outputs to other subcortical structures, rather than disruptionof motorcorticalprocessing.
From the perspective of predictive coding, the role of beta activity fits comfortably with the observations in
the visual system that top-down beta modulates the
excitability of evoked responses. In the motor system, betaoscillationsmayreflecttheprecisionorgainafforded byproprioceptivesignals—asisevidentbytheir attenu-ationduringmovement.Thisattenuationhasbeenlinked
to sensory attenuation during self-made acts [49],
suggestingafailureofsensoryattenuationinParkinson’s disease that rests on dopaminergic modulation of beta activity[5,50].
Conclusion
Thisreviewhasconsideredseveralperspectivesonhow LFPoscillationscanbeusedtoinformcomputationaland clinicalmodelsofneuronalcoupling.Wehavefocusedon DCMasawayofformalisinghypothesesaboutdirected (effective) connectivity. In closing, it should be noted that—unlike descriptive (functional) connectivity mea-suresof statisticaldependencies—effectiveconnectivity is only as goodas the modelthat definesit.Clearly,to fully harnessthemacroscopic dynamicsof electrophysi-ology, there is a long road ahead to validate current models in terms of microscopic and intracellular pro-cesses.
Conflict
of
interest
statement
Nothing declared.Acknowledgements
K.F.andD.P.arefundedbytheWellcomeTrust(GrantNo.088130/Z/09/Z). V.L.issupportedbytheNationalInstituteforHealthResearch UniversityCollegeLondonHospitalsBiomedicalResearchCentre.We wouldalsoliketothankourreviewerforguidanceinpresentingthese ideas.
References
and
recommended
reading
Papersofparticularinterest,publishedwithintheperiodofreview, havebeenhighlightedas:
ofspecialinterest
ofoutstandinginterest
1. RaoRP,BallardDH:Predictivecodinginthevisualcortex:a functionalinterpretationofsomeextra-classical receptive-fieldeffects.NatNeurosci1999,2:79-87.
2. FristonKJ,KiebelSJ:Predictivecodingunderthefree-energy principle.PhilosTransRSocB2009,364:1211-1221.
3. BastosAM,UsreyWM,AdamsRA,MangunGR,FriesP, FristonKJ:Canonicalmicrocircuitsforpredictivecoding. Neuron2012,76:695-711.
4. FletcherPC,FrithCD:Perceivingisbelieving:aBayesian approachtoexplainingthepositivesymptomsof schizophrenia.NatRevNeurosci2009,10:48-58.
5. AdamsRA,StephanKE,BrownHR,FrithCD,FristonKJ:The computationalanatomyofpsychosis.FrontPsychiatry2013,
4:47.
6. FeldmanH,FristonKJ:Attention,uncertainty,andfree-energy. FrontHumNeurosci2010,4:215.
7. ChawlaD,LumerED,FristonKJ:Therelationshipbetween synchronizationamongneuronalpopulationsandtheirmean activitylevels.NeuralComput1999,11:1389-1411.
8. MoranRJ,SymmondsM,StephanKE,FristonKJ,DolanRJ:Anin vivoassayofsynapticfunctionmediatinghumancognition. CurrBiol2011,21:1320-1325.
9.
ArnalL,GiraudA:Corticaloscillationsandsensorypredictions. TrendsCognSci2012,16:390-398.
Areviewhighlightingrecentempiricalandtheoreticalworklinking neu-ronal oscillationsat gammaand beta bands toprediction error and predictions,respectively,withafocusontheauditorysystem. 10. FellemanD,VanEssenDC:Distributedhierarchicalprocessing
intheprimatecerebralcortex.CerebCortex1991,1:1-47.
11. BauerM,StennerM,FristonK,DolanR:Attentionalmodulation ofalpha-betaandgammaoscillationsreflectfunctionally distinctprocesses.JNeurosci2014.(inpress).
12.
LeeJ,WhittingtonM,KopellN:Top-downbetarhythmssupport selectiveattentionviainterlaminarinteraction:amodel.PLoS ComputBiol2013,9:e1003164.
Thiscomputational studymodelshow atop-downattention-directing signal atbetafrequencies canbeintegratedwithinacorticalcolumn modeled with laminar resolution to enhance processing and boost gammaoscillationsoftheattendedinput.
13.
MarkovN,Ercsey-RavaszM,VanEssenD,KnoblauchK, ToroczkaiZ,KennedyH:Corticalhigh-densitycounterstream architectures.Science2013,342:1238406.
Areviewsummarizingrecentworkontheanatomicalconnectivityofthe neocortexbasedonretrogradetracingstudiesThisbodyofworkaddsto theevidence thatlaminar-specificconnectionsdefinebottom-up and top-downcounterstreams,formingadense,highlyinterconnected cor-ticalhierarchy.
14. DaunizeauJ,DavidO,StephanKE:Dynamiccausalmodelling:a criticalreviewofthebiophysicalandstatisticalfoundations. Neuroimage2011,58:312-322.
15. FristonKJ,BastosA,LitvakV,StephanKE,FriesP,MoranRJ:
DCMforcomplex-valueddata:cross-spectra,coherenceand phase-delays.Neuroimage2012,59:439-455.
16. DavidO,KiebelS,HarrisonL,MattoutJ,KilnerJ,FristonK:
DynamiccausalmodelingofevokedresponsesinEEGand MEG.NeuroImage2006,30:1255-1272.
17. DavidO,GuillemainI,SailletS,ReytS,DeransartC,SegebarthC, DepaulisA:IdentifyingneuraldriverswithfunctionalMRI:an electrophysiologicalvalidation.PLoSBiol2008,6:2683-2697.
18. MoranRJ,JungF,KumagaiT,EndepolsH,GrafR,DolanRJ, FristonK,StephanKE,TittgemeyerM:Dynamiccausalmodels andphysiologicalinferenceavalidationstudyusingisoflurane anaesthesiainrodents.PLoSONE2011,6:e22790.
19. BrownH,FristonK:Dynamiccausalmodellingofprecisionand synapticgaininvisualperception–anEEGstudy.Neuroimage 2012,63:223-231.
20. BolyM,GarridoMI,GosseriesO,BrunoMA,BoverouxP, SchnakersC,MassiminiM,LitvakV,LaureysS,FristonK:
Preservedfeedforwardbutimpairedtop-downprocessesin thevegetativestate.Science2011,332:858-862.
21. PinotsisD,MoranR,FristonK:Dynamiccausalmodelingwith neuralfields.Neuroimage2012,59:1261-1274.
22. DecoG,JirsaVK,RobinsonPA,BreakspearM,FristonK:The dynamicbrain:fromspikingneuronstoneuralmassesand corticalfields.PLoSComputBiol2008,4:e1000092.
23. AngelucciA,BressloffP:Contributionoffeedforward,lateral andfeedbackconnectionstotheclassicalreceptivefield centerandextra-classicalreceptivefieldsurroundofprimate V1.ProgBrainRes2006,154:93-120.
24. PinotsisD,SchwarzkopfD,LitvakV,ReesG,BarnesG,FristonK:
Dynamiccausalmodellingoflateralinteractionsinthevisual cortex.Neuroimage2013,66:563-576.
25. SceniakMP,HawkenMJ,ShapleyR:Visualspatial
characterizationofmacaqueV1neurons.JNeurophysiol2001,
85:1873-1887.
26. SceniakM,RingachD,HawkenM,ShapleyR:Contrast’seffect onspatialsummationbymacaqueV1neurons.NatNeurosci 1999,2:733-739.
28.
SaniI,SantandreaE,GolzarA,MorroneMC,ChelazziL:Selective tuningforcontrastinmacaqueareaV4.JNeurosci2013,
33:18583-18596.
This paper includes experimental results about selective contrast responsefunctionsbasedonspikingactivityfrommacacqueareaV4 inresponse tostaticstimulipresentedatvaryingcontrastlevels.The authors conjecture that contrast is not only a measure of stimulus intensity,butalsoabasicfeature,likecolorororientation.
29. DesimoneR:Visualattentionmediatedbybiasedcompetition inextrastriatevisualcortex.PhilosTransRSocLondonSerB 1998,353:1245-1255.
30. RayS,MaunsellJ:Differencesingammafrequenciesacross visualcortexrestricttheirpossibleuseincomputation.Neuron 2010,67:885-896.
31.
BosmanC,SchoffelenJ-M,BrunetN,OostenveldR,BastosA, WomelsdorfT,RubehnB,StieglitzT,DeWeerdP,FriesP:
Attentionalstimulusselectionthroughselective
synchronizationbetweenmonkeyvisualareas.Neuron2012,
75:875-888.
Using electrocorticographic and local field potential recordings, the authors showed that synchronization between rhythms in areas V1 andV4impliedaselectiveenhancementofrelevantsynapticinput,while themonkeyattendedtooneoftwosimultaneouslypresentedstimuli; a finding consistent with the ‘communication through coherence’ hypothesis.
32. PinotsisD,BrunetN,BastosA,BosmanC,LitvakV,FriesP, FristonK:Contrastgaincontrolandhorizontalinteractionsin V1:aDCMstudy.Neuroimage2014,92C:143-155.
33. TraubR,JefferysJ,WhittingtonM:Simulationofgamma rhythmsinnetworksofinterneuronsandpyramidalcells. JComputNeurosci1997,4:141-150.
34.
OswalA,BrownP,LitvakV:Synchronizedneuraloscillations andthepathophysiologyofParkinson’sdisease.CurrOpin Neurol2013:1-9.
Areviewsummarizingtheknownoscillatoryphenomenainthe cortico-BGcircuitinPDandtheirrelationtoclinicalimpairment.
35. EusebioA,ThevathasanW,DoyleGaynorL,PogosyanA,ByeE, FoltynieT,ZrinzoL,AshkanK,AzizT,BrownP:Deepbrain stimulationcansuppresspathologicalsynchronisationin parkinsonianpatients.JNeurolNeurosurgPsychiatry2011,
82:569-577.
36. PogosyanA,GaynorLD,EusebioA,BrownP:Boostingcortical activityatBeta-bandfrequenciesslowsmovementinhumans. CurrBiol2009,19:1637-1641.
37. JoundiRA,JenkinsonN,BrittainJ-S,AzizTZ,BrownP:Driving oscillatoryactivityinthehumancortexenhancesmotor performance.CurrBiol2012,22:403-407.
38.
LittleS,PogosyanA,NealS,ZavalaB,ZrinzoL,HarizM,FoltynieT, LimousinP,AshkanK,FitzGeraldJ,GreenAL,AzizTZ,BrownP:
AdaptivedeepbrainstimulationinadvancedParkinson disease.AnnNeurol2013,74:449-457.
Firstdemonstrationoftheuseofsubthalamicbetapowerasacontrol signalforclosedloopDBS.
39. HolgadoAJN,TerryJR,BogaczR:Conditionsforthegeneration ofbetaoscillationsinthesubthalamicnucleus-globuspallidus network.JNeurosci2010,30:12340-12352.
40. Nevado-HolgadoAJ,MalletN,MagillPJ,BogaczR:Effective connectivityofthesubthalamicnucleus–globuspallidus networkduringParkinsonianoscillations.JPhysiol2014,
592:1429-1455.
41. LitvakV,JhaA,EusebioA,OostenveldR,FoltynieT,LimousinP, ZrinzoL,HarizMI,FristonK,BrownP:Restingoscillatory cortico-subthalamicconnectivityinpatientswithParkinson’s disease.Brain2011,134:359-374.
42. HirschmannJ,O¨ zkurtTE,ButzM,HomburgerM,ElbenS, HartmannCJ,VesperJ,WojteckiL,SchnitzlerA:Distinct oscillatorySTN-corticalloopsrevealedbysimultaneousMEG andlocalfieldpotentialrecordingsinpatientswith
Parkinson’sdisease.Neuroimage2011,55:1159-1168.
43. LitvakV,EusebioA,JhaA,OostenveldR,BarnesG,FoltynieT, LimousinP,ZrinzoL,HarizMI,FristonK,BrownP: Movement-relatedchangesinlocalandlong-rangesynchronizationin Parkinson’sdiseaserevealedby
simultaneousmagnetoencephalographyandintracranial recordings.JNeurosci2012,32:10541-10553.
44. MoshelS,ShamirRR,RazA,NoriegaFR,deER,BergmanH, IsraelZ:Subthalamicnucleuslong-rangesynchronization—an independenthallmarkofhumanParkinson’sdisease.Front SystNeurosci2013,7:79.
45. HohlefeldFU,HuchzermeyerC,HueblJ,SchneiderG-H,NolteG, Bru¨ckeC,Scho¨neckerT,Ku¨hnAA,CurioG,NikulinV:Functional andeffectiveconnectivityinsubthalamiclocalfieldpotential recordingsofpatientswithParkinson’sdisease.Neuroscience 2013,250:320-332.
46. LittleS,TanH,AnzakA,PogosyanA,Ku¨hnA,BrownP:Bilateral functionalconnectivityofthebasalgangliainpatientswith Parkinson’sdiseaseanditsmodulationbydopaminergic treatment.PLOSONE2013,8:e82762.
47. MoranRJ,MalletN,LitvakV,DolanRJ,MagillPJ,FristonKJ, BrownP:Alterationsinbrainconnectivityunderlyingbeta oscillationsinParkinsonism.PLoSComputBiol2011,
7:e1002124.
48.
MarreirosAC,CagnanH,MoranRJ,FristonKJ,BrownP:Basal ganglia-corticalinteractionsinParkinsonianpatients. Neuroimage2012,66C:301-310.
DCMstudyofcortico-basalgangliacircuitbasedoncombined intracra-nial andEEG data inPD patients. Showsthe potential of DCM for identifyingmechanismsofpathologyanddesigningpossibletreatment strategies.
49. BrownH,AdamsRA,PareesI,EdwardsM,FristonK:Active inference,sensoryattenuationandillusions.CognProcess 2013.[Epubaheadofprint].
50. GeorgeJ,StrunkJ,Mak-McCullyR,HouserM,PoiznerH,AronA:
DopaminergictherapyinParkinson’sdiseasedecreases corticalbetabandcoherenceintherestingstateand increasescorticalbetabandpowerduringexecutivecontrol. NeuroimageClin2013,3:261-270.