0022-538X/94/$04.00+0
Copyright
C) 1994,AmericanSociety
forMicrobiologySplice
Site
Selection
in
Polyomavirus Late Pre-mRNA Processing
DAVID B. BATT,LISAM. RAPP, ANDGORDON G.CARMICHAEL*
Departmentof Microbiology, UniversityofConnecticutHealth Center, Farmington, Connecticut 06030 Received 23 September 1993/Accepted2 December 1993
Polyomavirus
latepre-mRNAs contain one5' splice site and two message body 3' splice sites,which arenot used atequal frequencies. As a result of alternative splicing, the total latemRNApopulationconsistsof about 5% mVP2(no message bodysplice chosen), about 15% mVP3 (promoter-proximal3' splice site chosen), and about 801%mVP1 (promoter-distal3' splice site chosen).Todetermine whetherit issplicesite strength that determines the ratio ofspliced products, constructs containing duplicated or rearranged 3' splice sites were created. In construct VP1,1, 160 bpsurrounding the VP33' splice site was substituted with the corresponding region of the VP13' splice site. This construct resulted in the duplication of the VP1 3' splicing signal. VP3,3 (two identical VP3 3' splice sites) and VP1,3 (VP1 and VP3 3' splice sites reversed) were similarly created. Each construct maintained wild-type spacingbetween the3' splice sites.Analysis
of RNAsfrom transfections showed that in each construct,the3' splice site closestto thepolyadenylation site was usedpreferentially. Analysisof anumber of additionalconstructs indicatedthat there are no strongcis-acting positiveornegative regulatorsofpolyomavirus
latesplicing; rather, splicing choices appear to be determined
largely
byrelativeposition of splice sites.Alternative
splicing of pre-mRNAs is commonly used to regulate cellular and viral gene expression and to generate diversity. Although many of the molecular details of constitu-tivesplicing
have been described(20),
lessis known about the regulation of alternative splice site selection at the molecular level.There are a number of ways in which splicing signals can influence alternative splice site choice (41). The strength of 5' splice sites has been shown to influence alternative splicing; 5' splice sites that are nearer to consensus bind
Ul
ribonucleo-protein more efficiently and are used more frequently than nonconsensus sites (9, 16, 19, 24, 28, 45). For example, in Drosophila melanogaster, the muscle myosin heavy-chain premessage is alternately spliced to generate some mRNAs containing exon 18 and somelacking
it.Animproved 5' splicesite
attheexon18-intron 18 border increased theefficiency
ofintron
18 removal in an invitro system containing exon 18, intron 18, and exon 19(24).
Insimian
virus 40 (SV40), alternativerecognition
of two 5'splice
sites generates large andsmallT-antigenmRNAs.The relativestrengthsof the twosplice
siteswereshowntoinfluence the relative levels oflarge T and small T messages(16, 45).
In asynthetic
systemcontaining
fragments of the humanP-globin
gene, the inclu-sion of anoptional
33-nucleotide(nt)
internal exon was influenced by the sequence of the 5' splice site, with exon inclusionincreasing sixfold when the sitewas consensus(9).
Thestrength of 3' splice sites can also influence alternative
splicing.
In the D.melanogaster
musclemyosin heavy-chain
system mentioned above, inclusion of exon 18 is also influ-enced
by
thestrength
of the 3'splice site,
with consensus sequencesleading
togreaterinclusion(24).
InRoussarcomavirus,
balancedsplicing
generating
bothunspliced
andspliced
mRNAsis achieved bytwo weak 3' splice sites, which allows much of thepre-mRNAtoremainunspliced, with the remain-derbeing
alternatively spliced(34).
Inthis system,however, 3'splice
sites andtheirsurroundings
mayalso influencesplicing
by
a different mechanism. When the 3' splice site for the*Correspondingauthor. Mailingaddress:Department of
Microbi-ology, University of Connecticut Health Center, Farmington, CT 06030. Phone:(203)679-2259. Fax: (203)679-1239.
promoter-proximalenvgeneisdeleted, the relativeamountof src mRNA
increases;
however, when the 3'splice
site of thepromoter-distal
srcgeneisdeleted, the relativeamountof env mRNAdoesnotincrease(3).
The
polypyrimidine
stretch and branchpoint sequencescan influence bothsplice
site strength and alternativesplicing.
In Roussarcomavirus,
a24-bp
insertcreating
a newbranchpoint
increased the amount ofenv mRNAand altered the
ratio
of spliced versusunspliced
message(15).
In SV40, multiple branchsitesareimportant
inregulating
theratio oflarge
T to small T mRNAs, and mutations in these sites can alter the ratio of mRNAsby
100-fold(14,
37).
Branchpoint recognition
alsoplaysarole in the alternative choice oftwoclosely spaced 3'
splice
sites inpolyomavirus early pre-mRNA processing
(18).
Intherat,-tropomyosin
system,thepolypyrimidine
tract andasequence upstreamof the 3'splice
siteareimportant
indetermining
the inclusion or exclusion of themuscle-specific
exon(exon 7)
(23).
In asystemusing splicing signals
from the human,-globin
gene,both in vitro and invivo,
theinclusion of a short internal exonwas influencedby
the sequence of the branchpoint
and thepolypyrimidine stretch,
with consensussignals
increasing
the inclusion ofa 33-ntexonby
10-foldor more(9). Finally,
in yeastcells,
theuseof alternative 3'splice
sitescanbe influenced
by
thelength
ofaU-richtractpreceding
the 3'
splice
site(38).
Several
cis-acting
elements other than thosedirectly
in-volved insplicing
play a role insplice
site choice.Negative
regulators
ofsplicing
(NRS)
arecis-acting
elements found in Rous sarcoma virus which affect the ratio ofspliced
versusunspliced
mRNA.These sequences work in thesense orienta-tiononly
whenplaced
near a5'splice
site(35).
Inadenovirus type2,
acryptic
5'splice
site inearly region
3is
suppressed by
a 120-nt segment located upstream of the
splice
site(8).
Removal of this segment resulted ina
competition
betweenthe natural and thecryptic site,
with the ratio determinedby
the relativestrengths
of the two sites(8).
Secondary
structurealso has beenreported
toplay
arole in alternativesplicing; however,
its roles in vitroandin vivo may be different(42, 43).
In aninvitro system,constructs contain-ing palindromic sequences which could sequestersplice
sites revealed thatsecondary
structure caninhibitsplicing,
presum-ably by
masking
thecis-acting signals
(43).
When these con-1797on November 9, 2019 by guest
http://jvi.asm.org/
1798 BATT ET AL.
late leader
1
i1o
_,oM
480p 46C0 4400 4209 400 3320v3609 3401 300S 2809mlivr- 4____
mVP3
-16%
mVPI
-80%
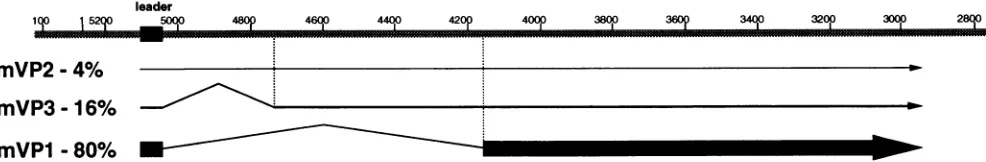
FIG. 1. Polyomavirus late pre-mRNA processing. Polyomaviruslatepre-mRNAmoleculescanbealternatively splicedtoform threeproducts, mVPI, mVP2,and mVP3. mVP2accountsforabout 5% of total latemessages,mVP3accountsfor about15%, andmVP1accountsforabout80%.
Thetoplinerepresentstheviral lateregionand shows thepositionof the late 5'noncodinglate leaderexonwhich isattachedtoeachlatemessage.
The thicknessof the linesreflects therelative proportions ofthe latemessages.
structswere analyzed in vivo, the secondarystructureseemed incapable of influencing splicing unless the stem structure
exceeded50 bp in length (43). Secondarystructureappearsto
play a role in the selection of splice sites in human growth
hormone (11), chicken 3-tropomyosin (10, 30), adenovirus EIA (5), and immunoglobulin heavy-chain (44) pre-mRNA splicing. In thesecases,the secondarystructure maysequester important cis-acting signals such asbranch points or3' splice
sites.
Splice site spacingcanalso affectsplicesite choice.InSV40, the ratio between large T and small T mRNAs results partly from the short small T intron, which isnotremoved efficiently (16). In a 3-globin gene derivative, the relative use of two
closely spaced competing 5' splice sites could be altered by changing the spacing between them. In this system,theuseof
theproximal splice site declined sharply when itwasseparated
bymore than 40bp from the distal site (7).
In addition to cis-acting signals, trans-acting factors
influ-ence alternative splice site choice. The effect of trans-acting
factors has been studied best in D. melanogaster, in which factors that bind near splice sites have been shown to either
suppress orenhance theiruse(33).Twocellularproteins which
can influence the choice oftwo 5' splice sites in SV40 early pre-mRNA processing have been described. At higher
concen-trations, the essential splicing factor ASF (alternative splicing factor)/SF2 (17, 26) allowed increased use of the promoter-distal smallT5'splice site relativetouseof theproximal large
Tsite (17), while another factor, DSF (distal splicing factor), promoted the useof the large T 5' splice siteoverthe small T
site (21).
Polyomavirus late pre-mRNAs are alternatively spliced to
formmessagesfor thestructural proteins VP1, VP2, and VP3.
Differences in the frequency of useoftwo alternative late 3' splice sites leads to ratios of about 5% mVP2 (no message
body splice chosen), 15% mVP3 (proximal 3' splice site chosen), and 80% mVPI (distal 3' splice site chosen) (Fig. 1). We have investigated the elements that influence the relative
useofeach 3' splice site. Toassesswhethersplice site strength determinesthe ratio ofspliced products,constructscontaining duplicatedorrearranged splice siteswerecreated. To examine
the possibility of positive or negative regulators, an insertion
and several deletion constructs were made. Finally, to deter-mine whether spacing is important, deletions which brought the two 3' splice sites closer to each other orwhich brought
both sites closer to the polyadenylation signalwere created.
Resultsindicate that there are no strongcis-acting regulators ofpolyomavirus late splicing; rather, splicing choicesappearto
be determined largely by the relative positions of splice sites within thetranscription unit.
MATERIALS AND METHODS
Restriction enzymes, T4 DNA ligase, DNA polymerase I
large fragment, T4 DNA polymerase, and T4polynucleotide kinase were from New England Biolabs and were used as
suggested by the manufacturer. RNase T2 was prepared as
describedbefore(31). [_x-32P]UTPwasfrom Amersham. Poly-omavirus strain 59RA was used and has been described
elsewhere(12, 13). The numberingsystemused here has been described elsewhere (4). All recombinant plasmids used here
were propagated in Escherichia coli JM83.
Cell culture and transfections. Mouse NIH 3T3 cellswere
obtained from the American Type Culture Collection. The techniques for their propagation and transfection have been described in detail elsewhere (4). Before transfection, all recombinant plasmids were cut with EcoRI to release intact viral genomes and dilute ligated with T4 DNA ligase as
described previously(1).
Constructs used. The startingconstructfor allwork, pPy*, consisted of the wild-type polyomavirus strain 59RAgenome
insertedatits unique EcoRI site intoamodifiedpUC18vector
that lacked all sitesin the multisite cloning regionexceptthe EcoRI site. Site-directed mutagenesiswasthenusedtocreate
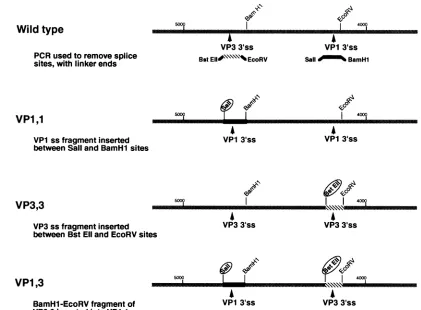
a Sall site 80 bp upstream of the VP3 3' splice site (clone pPy*C3) andto createaBstEII site80bpupstreamof theVP1 3' splice site (clone pPy*C5). PCR was used to amplify
sequencesspanning the VP1 3' splice site (80 bpupstreamand
80bp downstream of the 3' splice site) while also generating SallandBamHIrestriction sitesattheends(Fig. 2). PCRwas
similarly usedtoamplifysequencesspanning the VP3 3' splice site (80 bp upstream and 80 bp downstream of the 3' splice site)whilealsogenerating BstEII and EcoRV restriction sites
at the ends (Fig. 2). Clone VP1,1 was created by digesting
pPy*C3 and the VP1 3' splice site PCR product with Sall and BamHI and ligating the digested PCR product with the large fragment of pPy*C3 (Fig. 2). Clone VP3,3 was created by
digesting pPy*C5 and the VP3 3'splice site PCR product with BstEII and EcoRV andligating the digested PCR product with the large fragment of pPy*C5 (Fig. 2). VP1,3was created by removing the BamHI-BsaBIfragment from VP1,1 and replac-ing it with the samefragment fromVP3,3.
Constructp5'ss/Sacwasmadeby insertingacopyof the late leader 5' splice site into theSacl sitelocated atposition 4367 inthe late region ofpolyoma strain 59RA. The fragment used
wasdescribed elsewhere(32) and contained 6 bpupstreamand 15 bp downstream of the late leader 5' splice site. Construct pC5BBdel is a deletion of the BamHI-to-BstEII fragment of
clone pPy*C5 (Fig. 2). Construct pRVHdel was created by deletionofanEcoRV-HincII fragment, leavingan81-bp VP1
exon. Construct pHdel carries a deletion of the HinclI
frag-*rnlDO
- AOL--I
I---mmmp.
0-J.VlIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.63.556.88.169.2]Wild
type
PCR usedtoremovesplice sites,withlinker ends
VP1 ,1
VP1ssfragmentinserted betweenSallandBamHl sites
VP3,3
VP3 ssfragmentinserted between Bst Elland EcoRVsite
VP33'ss VPI 3'ss
BatEll90X
'%
EcoRV Sall _ BamHl5005>~~~~~ ~
~~~~~~~~~~~~~~~~~~~---VP13'ss VPI3'ss
500 4009
VP33'ss
s
5N
5000
VP1
,3
BamHl-EcoRV
fragment of VP1 3'ss VP33'ss
[image:3.612.94.515.79.389.2]VP3,3inserted into VP1,1
FIG. 2. Splice site duplication and rearrangement constructs. Constructs
VPl,l,
VP3,3, andVPI,3 were made as described in Materials and Methods. Briefly, PCR was used to amplify theVP1 and VP3 splice sites (80base pairs upstream and 80 base pairs downstream of the splice junctions). PCR primersweredesigned such that the amplified fragments contained convenient restriction enzyme cutting sites at their edges. Site-directedoligonucleotide mutagenesiswasthen usedtointroduce restriction sitesneareachsplice site(circled). Theintroduced restriction site alongwithanaturally occurringoneallowed thesplice sitestobe removed andreplaced with PCR-generated fragments. This allowed the creation ofVP1,1 (twoVPI 3'splice sites[3'ss])andVP3,3 (two VP3 3'splice sites).InbothVP1,1andVP3,3, the spacing between the 3' splice sites was closetothespacingfound in the wild type. ConstructVP1,3wascreatedbyexchangingthe BamHI-BsaBIfragment ofVPl,lfor the BamnHI-BsaBI fragmentofVP3,3.mentofthe lateregion. ConstructpPY-200Acarries a deletion ofapproximately200bpbetween the 5'splice siteofthe leader exon and the VP 3 3' splice site and was created by oligonu-cleotidemutagenesis.
Harvestingofcytoplasmic RNA.Cellswere harvested 44 to 48 hafter initiationof transfection. Eachplatewasrinsedwith Ix PBS- (phosphate-buffered saline
[PBS]
withoutMg2
andCa2+),
and RNA was harvested by the protocol described elsewhere (25). Briefly, cells were scraped into PBS and pelleted bycentrifugation at 1,000 x gfor 3 min. The pellet wasresuspendedinPBSandcentrifugedagain at1,000 xgfor 3 min.The cellpelletwasresuspended in 3 mlof NonidetP-40lysis
bufferandleftonice for10 min.Nucleiwere removed by centrifugationat1,000
xgfor 3 min. To the supernatant (the cytoplasmic RNA fraction) was added 1.42 g of guanidine thiocyanate. RNA wasseparatedfrom DNAbycentrifugation througha5.7 M cesium chloridecushionfor 20 h at110,000
x g(6).The pelletedRNAs were drained well and resuspendedin 300
pl
ofRNase-free water.Riboprobe preparation and RNase T2 protection assays. All riboprobeswere made fromclones by usinginvitro transcrip-tion withT3 or T7 RNApolymeraseand
[32P]UTP,
creatingan internal labeled probe. For constructsVP1,1, VP1,3,
and VP3,3, thecorresponding HindIIItoEcoRI fragmentsspanning the latesplice sites weresubcloned into pBS+. The probe forp5'ss/Sacwascreatedbycloning the EcoNI fragment into the SmaIsite ofpBS+. Theprobeclone forpRVHdelwasmadeby cloning thePstI-ApaI
fragment
surroundingthe altered region into the same sites in pBlueScript SK. Internally labeled riboprobeswereannealedto25 to 50 ,ug ofcytoplasmicRNAat 57°C for 15 to 20 h. Theresulting hybridsweredigestedwith 15[LI
of RNaseTF/T,
(31)at37°C for2h. Afterphenolextraction and ethanolprecipitation, the samplesweredissolved in dena-turing dye, and protected bandswere resolved on 6% polyac-rylamide-urea sequencing gels. Band intensitieswere quanti-fied with a Betascope Blot Analyzer, and afteradjusting
for uridine content,relative percentageswereassigned.
RESULTS
Theprocessingofpolyomaviruslatepre-mRNAsis shown in Fig. 1. The mRNA for VP1 is formed when the 57-base noncoding late leader exon isjoined to the 3'
splice
site at position4158. The mRNA for VP3 is formed when the leader isjoined tothe 3' splicesiteat position4733. The mRNA for VP2isformedwhen the leaderexon isnotspliced
toeither 3' splice site. To measure theproportion
of mRNAs forVP1,
VP2, andVP3, we constructedradioactive,
internally
labeled riboprobes which span the two messagebody
3'splice
sites. RNaseprotectionassayswereperformed
oncytoplasmic
RNAVP33'ss
400 N
46,
e
Aco
A
--- ----A
on November 9, 2019 by guest
http://jvi.asm.org/
1800 BATT ET AL.
A
501 489
404 353
242
190
B
DNA
-.o- nomessagebody splice
.m- upstream body splicesiteused
-downstream bodysplicesite used
2 3 4
upstream3'as downstream3' a
5' 5' 5'T
- undigested cRNA probe
nomessagebodysplice
upstreambodysplik siteused
downstreambody spike site used
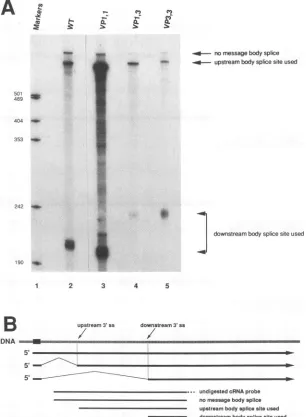
FIG. 3. Relativesplice site use in thewild type and inconstructs VP1,1, VP3,3, and VP1,3. Viral constructs were liberated fromplasmid backbones,recircularized,and transfected into NIH 3T3 cellsasdescribed in Materials and Methods. (A)CytoplasmicRNA washarvestedat48 h andsubjectedtoRNaseprotectionassaysusingriboprobesdiagrammedinpanelB. Arrowsindicate thepositions expected for protectedbands that representalternativesplicing products. WT,wild type.Markersare32P-labeledMspI fragmentsofpUC18;sizesareindicated in nucleotides.
harvested from cellstransfected with wild-type polyomavirus genomes.Results from RNase
protection
assaysindicated that the relativewild-type proportions
ofmVP1, mVP3, andmVP2 inpolyomavirus
late mRNA were80, 16,and4%, respectively(Fig.
3A, lane 2; Table1).
Todetermine whether the relative strength ofthevarious 3'
splice
sites dictatesthe ratio ofspliced productsin polyomavi-rus latepre-mRNA, constructs containing duplicated or rear-ranged splice sites were created (Fig. 2). First, site-directed mutagenesiswasused tointroduce unique restriction endonu-cleasesites upstream of each of the 3' splice sites. Use of these newsites inconjunction with naturally occurring unique down-streamrestriction sites allowed the release of each splicesite.Second,
PCRwasusedtoamplify fragments spanning
eachof the wild-type 3' splice sites. The oligonucleotides used toamplify
thesplice sites were designed to bind 80 bp on eitherside
of thesplice
sites and to create convenient restrictionendonuclease sites on the ends of the
amplified fragments.
Finally,
theconstructscontainingnewunique
restriction sites were digested with the appropriate enzymesto liberate frag-ments containing either the VP1 or the VP3 splice site, and these fragmentswere replaced with thePCR-generated
ones. Thisallowed the spacing between the 3'splice
sites in these constructs to be similar to that of the wild type. Construct VP1,1 contains two VP1 3'splice
sites, constructVP3,3
containstwoVP3 3'
splice
sites, andconstructVP1,3 contains bothsplice
sites but reversed from their wild-typepositions.
Constructs VP1,1 and VP3,3 allowed us to investigate the relative use oftwoidentical 3'splicesites, and construct VP1,3, when compared with the wild type, allowed us to investigate the role of
splice
sitestrength.
Results of RNase protection assays are shown in Fig. 3A, and
quantitation
is showninTable1.Values forrelative molar amountsof the variousprotected species have beenadjusted
toJ. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.157.464.82.499.2]TABLE 1. Relative splicesitechoice induplication and
rearrangement constructs
Bandintensity(%yc)'
Splicesiteused Wild
type
VPl?l
VPI,3 VP3,3None (unspliced) 4 1 3 2
Proximal 3' 16 36 44 15
Distal 3' 80 63 53 83
"The intensities of the bands (shown in Fig. 3A) were quantitated with a
Betagen Betascope blot analyzer and thenadjusted foruridine content (bands representinternally labeled fragments) to arrive at the values presented.
take into account the number of uridines present in the internally labeled protected fragments. For construct VPl,1 (Fig. 3A,lane 3), the proximal sitewasused36% of the time and the distal site was used 63% of the time, with the remaining 1% unspliced.Forconstruct VP3,3(Fig. 3A, lane 5), theproximal sitewasused 15% ofthe time and the distal site was used 83% of the time, with the remaining 2% unspliced. For construct VP1,3 (Fig. 3A, lane 4), the proximal site was used 44%ofthe time and the distal sitewas used53% ofthe time, with the remaining 3% unspliced. In repeated RNase protectionassays,percentages veryclosetothe datapresented wereobserved (datanotshown).RNaseprotectionassayswere also carried outwith shorterprobes (400 bp)which spanned onlythe distal splice site. Resultsobtainedwith these probes were consistent with those obtained with the longer probes (datanotshown). The conclusion from theseexperimentswas that splice site strength alone could not account for message ratios observed. This raised the possibility that splice site choice in this system is influenced bycis-acting sequences in the late transcriptional unit which lie outside the regions manipulated above.
The preceding results could be explained if there were an NRS which acted to suppress splicing to the proximal splice site, orif there were a positive cis regulatorofsplicing acting
A
on the distal splice site. Alternatively, 3' splice site choice could be determined by relative position. To determine whether there is a negative or positive regulator element in polyomavirus, several constructs were made. Negative cis regulation of theproximal 3' splice sitewastested byinserting a small DNA fragment containing 21 bp spanning the late leader 5' splicesite (as well as plasmid linker sequences) at a Saclsite 430 ntdownstream of the VP3 3' splice site (Fig.4A). If the VP3 3' splice site is negatively regulated, then this new exon should be skipped frequently, producing a 579-nt pro-tected band; however, if the splice site is not negatively regulated, the exon should not beskipped, producing a 43 1-nt protected band. An RNase protectionassay (Fig.4B) showed prominent protected bands of 431 and 254 nt but no band of 579 nt. The absence of a 579-nt protected band, along with carefulquantitation of thebands observed (almost equimolar levelsof the431- and254-nt species), revealed that theVP3 3' splice site is almost neverskippedinthis mutant.Further,this construct, as well as all others studiedhere, produced wild-type levels of late messages (data not shown). Results obtained from this experiment were consistent with the absence of an NRS for the proximal (VP3) 3' splice site. In a similar experiment reported previously (32), we showed that if the same5'splicesite was inserted within theVP1bodyexon, then the VP1 3' splice site was not skipped during pre-mRNA processing. Although this experiment indicatedthat the VP3 3' splicesite is notnegativelyregulated,itremainedpossiblethat theinsertion of the 5' splicesite in that constructstrengthened the VP3 3' splice site and/or overrode an NRS. Therefore, deletionsonboth sides of the VP3 3' splicesite were created. Construct pPY-200A (Fig. 5A) creates a 200-bp deletion upstream of the VP3 3' splice site, and construct pC5BBdel deletes 500bpdownstream of the VP3 3'splicesite(Fig. 5A). Construct pC5BBdel is discussed in greater detail in the following paragraph. Neither of these constructs significantly altered thesplicingratios (Fig.5B and C), arguing againstan NRS for the VP3 3' splicesite.
B
late VP3 VP1 leader 3 aa Sea 3ass
5'ss
0 CO,
Go
. 0.
Q
Q0
VP3splice only allsplice sites used VP1splice only
-skipleader and
VP3splicesites
mredictedbonds
riboprobe-1096 nt
946nt
41 nt 254n VP3spliceonly
useallsplice sites 254nt VP1spliceonly
579nt 2U nt skip VP3 3splice site only
<- probe-1096nt <- 948nt
-o.- 431nt
.-- 254nt
* S
2
FIG. 4. Analysis ofsplicing inconstructp5'ss/Sac.(A) The late region isdiagrammed, alongwith thepositionof the 5'splicesite(5'ss)inserted
inconstructp5'ss/Sac.An EcoNIfragmentspanningtheVP1 and VP33'splice sites(3'ss)wasusedtogeneratea1,096-ntspecificriboprobe for
RNase protectionassays. Sizes of bandsexpected for varioussplicingevents areshown.(B)Theriboprobeillustrated inpanelAwasused inan
RNaseprotectionassayofcytoplasmicRNAharvested 48 h aftertransfection ofNIH3T3cells withp5'ss/Sac.The absence ofa579-ntfragment indicates that the VP3 3' splicesite isnotskipped in thisconstruct.Almost all messagesappeartohave used allavailable splicesites.
RgssiblesDlkicng
events
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.60.298.100.170.2]1802 BATT ET AL.
263
le Lr
5000
lateleader I
mVP33' as rr pPY-200 pCSBBd.l
4000 3000
A~~~~
nVPI3'5ss
pRVHd.l
at~~~~~~~"
I
aI
a.
VP2
-VP3 *.VP2+VP3
_
203nl .0;..
1 2 3 4 5
D
306n
203nr
190t-l
cc
VP1 -PI
[image:6.612.125.498.75.418.2]ri-1 2 3 4 5
FIG. 5. Analysis of deletionconstructs.(A)Schematicdiagramof thepolyomaviruslateregion (topline)andsequencesdeleted in constructs
pPY-200A, pC5BBdel, pRVHdel,andpHdel.The VP1 and VP3 3'splicesites(3'ss)areindicated,andtheBstEIIsitepresentinconstructPy*C5
isshown inboldface. (B) RNaseprotection assaysofcytoplasmicRNAs fromcellstransfectedwith the wild typeandpPY-200A. Assayswere
performedasdescribed in Materials andMethods, usingthe sameprobeasinFig. 3A, lane 2. Markersare the same as inFig. 3.(C) RNase
protectionassaysofcytoplasmicRNAs from cells transfected with thewildtype(WT),pC5BBdel,andpHdel. Assayswereperformedasdescribed
in Materialsand Methods. Markersarethesame asinFig.3.(D)RNaseprotectionassaysofcytoplasmicRNAs from cells transfected with the
wildtype (WT), pHdel,andpRVHdel. Markersarethe same asin Fig.3.
To determine whether the distal (VP1) 3' splice site is positively cis regulated,several additionalconstructsweremade (Fig. 5A).Figure 5C shows theresults ofanRNaseprotection
assay using two of these constructs and a VP1 3' splice
site-specific probe. Analysis of cytoplasmic RNA from wild-type-transfected cells (Fig. 5C, lane 1) revealed 76% VP1 splicing, with 24% mVP2 plus mVP3. Construct pC5BBdel lackssequencesfromBamHItoBstEIIinconstructpPy*C5.In thisconstruct,theVP1 andVP3 3'splicesitesareseparatedby 136 nt. Analysis of cytoplasmic RNAs from cells transfected with thisconstruct revealed67%VP1splicing,with 33% mVP2 plus mVP3 (Fig. 5C, lane 3). Construct pHdel lacks a504-bp
HincII fragment from the late region (Fig. 5A). Analysis of cytoplasmic RNAs harvested from cells transfected with this
construct(Fig.5C,lane4)revealedthesameratios(76%mVP1 and24% mVP2plus mVP3)asfoundfor thewildtype(lane 1). Figure 5D shows the analysis of splicing ratios in constructs
pRVHdelandpHdel. Construct pRVHdel lacks 1,147 bp of the VP1 bodyexon(Fig. 5A). For thisconstruct,aprotected band
of 81 ntrepresents splicing to the VP1 3' splice site, while a
protected band of 196nt represents unspliced VP2-plus-VP3-spliced molecules. Figure 5D, lane 4, shows that construct
pRVHdel uses the two 3' splice sites almost equally (52%
mVP1and 48% mVP2plus mVP3).Incontrast,pHdelshowed
asplicingratio similar tothatobserved for the wild type(Fig. SD,lanes 2 and3). Theconclusion from these experiments is
thatonlymodesteffectsonVP13'splicesite choiceresultfrom
deletionsinthe polyomavirus lateregion,with theonly signif-icantchangeinrelativesplicinglevelsbeingobserved whenthe
terminal VP1bodyexon wastruncatedtoonly82nt. That these
dataarguefor the absenceofapotentpositive regulatorofVPl
splicingwillbe addressed further below. DISCUSSION
Alternative3'splicesiteselectionduringRNAprocessingof
polyomavirus late pre-mRNA molecules generates multiple mRNAs.The promoter-distal VP1 3' splicesite is chosenthe
majority of the time. Here we have examined how this
pre-ferential choice is made. Is the VP1 3'splicesitestrongerthan
the VP3 3' splice site, thus allowing it to compete more
effectively forthe splicing machinery, or are there cis-acting
sequencesinthe lateregionthatinfluence 3'splicesite choice?
Asimple splice site strength model would predict that when
twoidentical 3' splice sitesare present,such asfor constructs
VP1,1 andVP3,3, theywould be used equally; likewise,when
A
B
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
the sitesarereversedintheir
order,
then the ratioofsplice
siteuseshould alsobe reversed.Thiswas notobserved.
Instead,
in this system,the distal 3'splice
sitewasalways preferred
over theproximal
site. Thatsplice
sitestrength probably plays
a minor role can best seenby comparing
the relativesplicing
frequencies
ofconstructsVP1,3
and VP3,3(Fig.
3 andTable1).
VP1,3
usestheproximal
site40% ofthe timeand the distal site 60% ofthetime,
whileVP3,3
usestheproximal
site 15% ofthe time and the distalsite 80% of the time.Theability
of the VP1 3'splice
sitetobe usedmorefrequently
thantheVP3 3'splice
site at the sameposition (proximal position)
argues that the VP1 3'splice
site is somewhatstrongerthan the VP3 3'splice
site.However,
it should bepointed
out that in absolute terms,both 3'splice
sitesarelikely
tobeweak. This conclusion is basedon the observation thatdespite
thepres-ence oftwo 3'
splice sites,
about 5% of the mRNA remainsunspliced
(mVP2). Further,
in numerous attempts, we have been unabletoobservepolyomavirus
late pre-mRNAsplicing
in vitro
(32a).
Asa
splice
sitestrength
modelcouldnotexplain
the resultsobserved,
thequestion
remained as towhy
the distal site ispreferred.
The ratios observed could result from either cisdownregulation
ofsplicing
totheproximal
site(such
asby
an NRSelement)
or cisupregulation
of the distal site(by
apositive element). Analysis
of a number of additionalcon-structs revealed that a strong
positive
or strongnegative
cisregulator
of eithersplice
site isunlikely.
Insertion ofa5'splice
site downstream of theVP3 3'
splice
sitebut upstream ofthe VPI 3'splice
site(construct p5'ss/Sac; Fig. 4A)
revealed no apparentdownregulation
oftheVP3 3'splice
site. Infact,
this 3'splice
sitewas nowusedvirtually
100% of the time(Fig. 4B),
indicating
that it is an efficient 3'splice
site in the propercontext. ApotentNRS elementwouldmost
likely
have ledtofrequent
skipping
of the VP3 3'splice
site. Deletions both upstream and downstream of the VP3 3'splice
site(Fig. 5)
confirmed the absence of an NRS. A series of deletion
constructs was then used to argue
against
positive
cisregula-tion ofthe distal 3'
splice
site. As seenfromthe aggregate of resultspresented
inFig.
5,
deletion ofessentially
allsequences between the VP3 3'splice
siteand the latepoly(A)
sitedonot alter the fact that the distal site is chosenpreferentially.
Reducing
the distancebetween the VP3andVPI 3'splice
sites(construct pC5BBdel)
resultedina modestchange
in relativesplicing
ratios. ConstructpHdel (500-bp
deletion in the VPIexon)
gave ratios identical towild-type ratios,
while alarge
deletion which truncated the VP1
body
exon to 81 nt(con-struct
pRVHdel)
resulted innearly equal
use of the twoalternative 3'
splice
sites. For this latterconstruct,we donot know whether thesomewhatalteredsplicing
ratio is duetothe presenceofaweakpositive regulator (which
has been deleted inthisconstruct)
or tothe fact thatnowthe terminalVPI exon is too short(82 nt)
to beefficiently
processed
oraccurately
detected in our RNaseprotection
assays.Attempts
tostudy
constructswith
intermediate-length
VP1 exons,or exonscon-taining heterologous inserts,
failed due to the activation ofcryptic splice sites,
making analysis
ofsplicing
ratiosimpossible
to
interpret.
Consistent with the lack ofapositive
ornegative
regulator
ofpolyomavirus
latesplicing,
wehave alsoobserved that a 6-nt deletion of the VPI 3'splice
site leads to an increase in the amounts of mVP2 andmVP3,
and a 6-nt deletion of the VP3 3'splice
site leads to an increase in theamountsof mVP1 and mVP2
(data
notshown).
The data
presented
above,
whileallowing
us to rule outstrong
positive
ornegative
cisregulators,
do not allow us to rule out the existence of several weakregulators placed
in different areas and which workadditively
orsynergistically.
Inthe work describedhere, we also did not rule out thepossibility of splice site regulation by two small regions at the extreme ends of the late viral transcription unit. A 45-bp region immediately downstream of the late leader5' splice site was notremoved, nor was the 50 bp immediately preceding the late polyadenylation signal.
The results discussed above show that the relative position-ing of splice sites in the transcript appears to be the most importantdeterminant ofsplice site choice in the polyomavirus late system. Even when two identical 3' splice sites exist in the same transcript, the distal site is preferred. This isconsistent with theinvivofindingsof Kuhne et al. (27),who found that constructs which contain two rabbit ,3-globin 3' splice sites always used the downstream site. However, it contradicts results of Lang and Spritz (29), who concluded that the upstream 3' splice site, in human
y-globin
constructs contain-ing duplicated 3' splice sites,wasalwayschosenin vivo.Finally, Reed and Maniatis (39) concluded from in vitro studies of human 3-globinconstructscontaining duplicated 3' splicesites thatthe upstream 3' splicesite waschosen ifthe second exon was205 nt inlength,and the downstream site was chosen if the secondexon wasshorter than 55 nt. However, these research-ersalso found that exonic sequences couldinfluence which site waschosen.One working model, which is consistent with the results reported here, is that the length of the late terminal exon influences theprocessingof the latepre-mRNA.Robberson et al. (40) postulated thatexonsrather thanintronsaretheinitial units of definition in pre-mRNA splicing. In their model, 3' splice sites are not normally used unless the downstream borderof theexon(eithera5' splice site for internalexons or apolyadenylation site for terminal exons) is also recognized. More recentevidencehasbeen presentedthat, in fact, eukary-otic exonsarescanned in a 5'-to-3' directionduring splicesite
selection,
and that exonskippingorsplice site skipping might result when exons are excessively long (36). Interestingly, in p5'ss/Sac,the exon createdbythe insertion ofthe 5'splice site is 431 nt long. While this is somewhat longer than the approximately300-nt length limit reported for most (but notall)
internalexons(36), it is nevertheless efficiently includedin maturemRNA. Inpolyomavirus late transcripts, theVP1 body exon is greater than 1 kb in length, already longer than the great majority of vertebrate 3' exons (22). This, coupled with the fact that both the VP1 3' splice site and the viral late polyadenylation signal may beweak, wouldlead to inefficient splicing of mVPI molecules, which is observed. However, otherresults shown heresuggestthatamodelrelying solelyon thespacing
of 3' splice site and the poly(A) signal cannot simply account for the results observed. Relative position, rather than spacing, appears to be the major determinant of splice site choice in the polyomavirussystem.Although relative splice site position is of primary impor-tance to
splice
sitechoiceinthis system,splicing ratiosarealso subtly influenced byavariety ofotherfactors,such asrelativesplice
site strengths and the spacing between splice sites. We arecurrently investigating
the role of each oftheseparameters in polyomavirus late pre-mRNA processing.ACKNOWLEDGMENTS
This work was supported by grant CA45382 from the National CancerInstitute.
We thankother members of thelaboratoryforhelpfulcomments on
the manuscript and N. Barrett for technical assistance in the early phaseof this work.
on November 9, 2019 by guest
http://jvi.asm.org/
1804 BATT ET AL.
REFERENCES
1. Adami, G. R., and G. G. Carmichael. 1986. Polyomavirus late
leader region serves and essential spacer function necessary for
viability and late gene expression. J. Virol. 58:417-425.
2. Ausubel, F. M., R. Brent, R. E. Kingston, D. D.Moore, J. A. Smith, J. G. Seidman, and K. Struhl (ed.). 1987. Current protocols in
molecular biology. Greene Publishing Associates and Wiley-Inter-science, New York.
3. Berberich, S. L., and C. M. Stoltzfus. 1991. Mutations in the
regions of the Rous sarcomavirus 3' splice sites: implications for
regulation of alternative splicing. J. Virol. 65:2640-2646.
4. Cahill, K. B.,and G. G.Carmichael.1989.Deletionanalysisof the polyomavirus latepromoter: evidence forbothpositive and neg-ative elements in the absence of early proteins. J. Virol.
63:3634-3642.
5. Chebli, K.,R. Gattoni, P. Schmitt, G.Hildwein, and J. Stevenin. 1989. The216-nucleotideintron ofthe Ela pre-mRNAcontainsa
hairpin structure that permits utilization of unusually distant branch acceptors. Mol. Cell. Biol.9:4852-4861.
6. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J. Rutter. 1979. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry
18:5294-5299.
7. Cunningham, S. A., A.J. Else, B. V. L. Potter, and I. C.Eperon. 1991. Influences of separation and adjacent sequences on the use of alternative 5' splice sites. J. Mol. Biol. 217:265-281.
8. Domenjoud, L., H.Gallinaro,L. Kister, S. Meyer, and M. Jacob. 1991. Identification of a specific exon sequence that is a major
determinant in the selection between a natural and acryptic 5' splice site. Mol. Cell. Biol. 11:4581-4590.
9. Dominski, Z., and R. Kole. 1992. Cooperation of pre-mRNA sequence elements in splice site selection. Mol. Cell. Biol. 12: 2108-2114.
10. D'Orval, B. C., Y. D. Carafa, P. Sirand-Pugnet, M. Gallego, E. Brody, and J. Marie. 1991. RNA secondary structurerepression of
a muscle-specific exon in HeLa cell nuclear extracts. Science 252:1823-1828.
11. Estes, P. A.,N. E. Cooke, and S. A. Liebhaber. 1992. Anative RNA secondary structure controls alternative splice-site selection and
generates two human growth hormone isoforms. J. Biol. Chem. 267:14902-14908.
12. Feunteun, J., L. Sompayrac, M. Fluck, and T. L. Benjamin. 1976. Localization of gene functions in polyomavirusDNA. Proc.Natl. Acad. Sci. USA 68:283-288.
13. Freund, R.,G. Mandel, G. G.Carmichael, J. P. Barncastle, C. J. Dawe, and T. L. Benjamin. 1987.Polyomavirustumorinduction in
mice: influences of viral coding and noncoding sequences on tumor profiles. J.Virol. 61:2232-2239.
14. Fu, X., H. Ge, and J. L. Manley. 1988. Therole ofthe
polypyri-midine stretch at the SV40 early pre-mRNA 3' splice site in
alternative splicing. EMBO J. 7:809-817.
15. Fu, X., R. A. Katz, A. M. Skalka, and T. Maniatis. 1991. The role of branchpoint and 3'-exon sequences in thecontrol of balanced
splicing of avianretrovirus RNA.Genes Dev. 5:211-220.
16. Fu, X., and J. L. Manley. 1987. Factors influencing alternative
splice site utilization in vivo. Mol. Cell. Biol. 7:738-748. 17. Ge, H., and J. L. Manley. 1990. Aprotein factor, ASF, controls
cell-specific alternative splicing ofSV40earlypre-mRNA in vitro.
Cell 62:25-34.
18. Ge, H., J. Noble, J. Colgan, and J. L. Manley. 1990. Polyoma virus small tumor antigen pre-mRNA splicing requires cooperation between two 3' splice sites. Proc. Natl. Acad. Sci. USA 87:3338-3342.
19. Grabowski, P. J., F. U. H. Nasim, H. C. Kuo, and R. Burch. 1991.
Combinatorial splicing of exon pairs by two-site binding of UI
small nuclear ribonucleoprotein particle. Mol. Cell. Biol.
11:5919-5928.
20. Green, M. R. 1989. Pre-mRNA processing and mRNA nuclear
export. Curr. Opin. Cell Biol. 1:519-525.
21. Harper, J. E., and J. L. Manley. 1991. A novel protein factor is required for use of distal alternative 5' splice sites in vitro. Mol.
Cell. Biol. 11:5945-5953.
22. Hawkins, J. D. 1988. A survey on intron and exonlengths.Nucleic Acids Res. 16:9893-9908.
23. Helfman, D. M., R. F.Roscigno, G. J. Mulligan, L. A.Finn,and
K. S.Weber. 1990. Identification of two distinct intron elements involved in alternative splicing of ,-tropomyosin pre-mRNA. GenesDev. 4:98-110.
24. Hodges, D., and S.I. Bernstein. 1992. Suboptimal 5' and 3'splice sites regulate alternative splicing of Drosophila melanogaster myosin heavychain transcripts in vitro. Mech. Dev. 37:127-140. 25. Hyde-DeRuyscher, R. P., and G. G. Carmichael. 1990.
Polyoma-virus late pre-mRNA processing: DNA-replication-associated changes in leader exon multiplicity suggest a role for leader-to-leadersplicing in the early-lateswitch. J.Virol. 64:5823-5832. 26. Krainer, A. R., G. C. Conway,andD.Kozak. 1990. The essential
pre-messenger RNAsplicing factor-SF2 influences 5' splice site selection byactivating proximalsites. Cell 62:35-42.
27. Kuihne, R., B. Wieringa, J. Reiser, and C. Weissmann. 1983.
Evidence against a scanning model for RNA splicing. EMBO J. 2:727-733.
28. Kuo,H. C., F. U. H. Nasim,and P.J.Grabowski.1991.Controlof
alternativesplicing by the differentialbindingofU1 small nuclear ribonucleoprotein particle. Science 251:1045-1050.
29. Lang, K. M., and R. A. Spritz. 1983. RNA splice site selection: evidence for a 5' to 3' scanning model. Science 220:1351-1355. 30. Libri, D., A. Piseri, and M. Y. Fiszman. 1991. Tissue-specific
splicing in vivo of the ,B-tropomyosin gene: dependence on an
RNAsecondary structure. Science 252:1842-1845.
31. Lichtler, A., N. L.Barrett, andG. G. Carmichael. 1992. Simple, inexpensive preparationof T1/T2 ribonuclease suitable forusein RNase protectionexperiments. BioTechniques 12:231-232. 32. Luo, Y., and G. G. Carmichael. 1991. Splice site skipping in
polyomavirus late pre-mRNAprocessing. J. Virol. 65:6637-6644. 32a.Luo, Y., and G. Carmichael.Unpublished data.
33. Maniatis, T. 1991. Mechanisms ofalternative pre-mRNAsplicing. Science 251:33-34.
34. McNally, M. T., and K. Beemon. 1992.Intronic sequences and 3' splice sites control Rous sarcoma virus RNA splicing. J. Virol. 66:6-11.
35. McNally, M.T., R. R. Gontarek, and K. Beemon. 1991. Charac-terization of Rous sarcoma virus intronic sequences that nega-tively regulate splicing. Virology185:99-108.
36. Niwa, M., C. C. MacDonald, and S. M. Berget. 1992. Are vertebrate exons scanned during splice-site selection. Nature (London)360:277-280.
37. Noble, J. C. S., C. Prives, and J. L. Manley. 1988. Alternative splicing of SV40 early pre-mRNA is determined by branch site selection. Genes Dev. 2:1460-1475.
38. Patterson, B.,andC. Guthrie. 1991.AU-richtractenhances usage ofanalternative 3' splice site in yeast. Cell 64:181-187. 39. Reed, R., and T. Maniatis. 1986. A rolefor exon sequences and
splice site proximity in splice-site selection. Cell 46:681-690. 40. Robberson, B. L., G. J. Cote, and S. M. Berget. 1990. Exon
definition may facilitatesplicesite selection in RNAs withmultiple
exons. Mol. Cell.Biol. 10:84-94.
41. Smith, C. W. J., J. G. Patton, and B. Nadal-Ginard. 1989. Alternative splicinginthecontrol of gene expression.Annu. Rev. Genet. 23:527-577.
42. Solnick, D. 1985. Alternative splicing causedby RNA secondary structure. Cell 43:667-676.
43. Solnick, D., and S. I. Lee. 1987. Amount of RNA secondary structurerequired to induce analternativesplice. Mol. Cell. Biol. 7:3194-3198.
44. Watakabe, A., K. Inoue, H. Sakamoto, and Y. Shimura. 1989.A
secondarystructure at the 3'splice siteaffectsthe in vitrosplicing reaction of mouse immunoglobulin-mu chain pre-messenger RNAs. Nucleic Acids Res. 17:8159-8169.
45. Zhuang, Y., H. Leung, and A. M. Weiner. 1987. The natural 5' splice site of simian virus 40large Tantigen can be improved by increasing the basecomplementaritytoUtRNA.Mol. Cell. Biol. 7:3018-3020.
J. VIROL.