Chapter 48
Overview: Lines of Communication
• Neurons are nerve cells that transfer information within the body
• Neurons use two types of signals to
communicate: electrical signals (long-distance) and chemical signals (short-distance)
Concept 48.1: Neuron organization and structure reflect function in information transfer
• Introduction to Information Processing
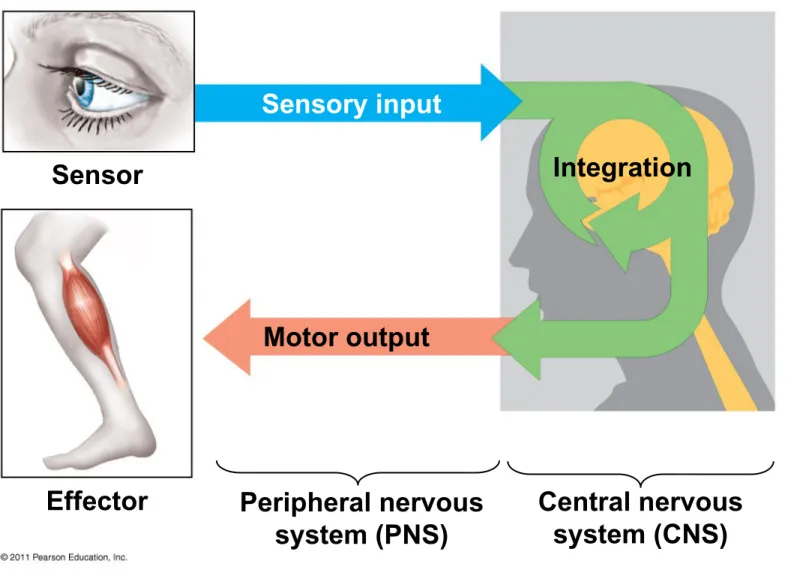
– Nervous systems process information in three stages: sensory input, integration, and motor output
• Sensors detect external stimuli and internal conditions and transmit information along
sensory neurons
• Sensory information is sent to the brain or ganglia, where interneurons integrate the information
• Motor output leaves the brain or ganglia via
motor neurons, which trigger muscle or gland activity
• Many animals have a complex nervous system that consists of
– A central nervous system (CNS) where
integration takes place; this includes the brain and a nerve cord
– A peripheral nervous system (PNS), which
carries information into and out of the CNS – The neurons of the PNS, when bundled
together, form nerves
Figure 48.3
Sensor
Effector
Sensory input
Motor output
Integration
Peripheral nervous system (PNS)
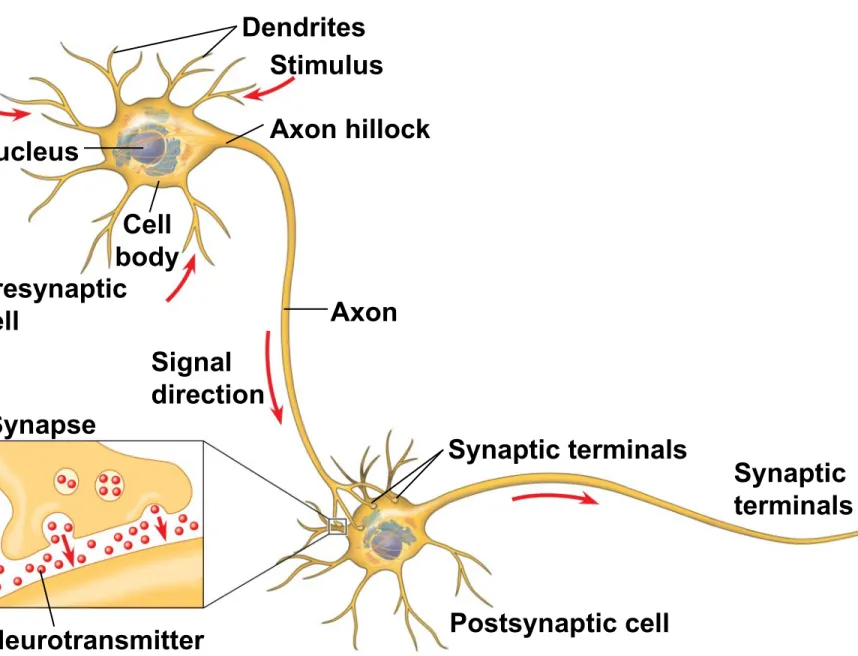
Neuron Structure and Function
• Most of a neuron’s organelles are in the cell body
• Most neurons have dendrites, highly branched extensions that receive signals from other
neurons
• The axon is typically a much longer extension that transmits signals to other cells at synapses • The cone-shaped base of an axon is called the
axon hillock
Figure 48.4
Nucleus
Dendrites Stimulus
Axon hillock
Cell body Presynaptic
cell
Signal direction
Axon
Synapse
Neurotransmitter
Synaptic terminals
Postsynaptic cell
• The synaptic terminal of one axon passes information across the synapse in the form of chemical messengers called neurotransmitters
• *A synapse is a junction between an axon and another cell
• Information is transmitted from a presynaptic
cell (a neuron) to a postsynaptic cell (a neuron, muscle, or gland cell)
• Most neurons are nourished or insulated by cells called glia
Concept 48.2: Ion pumps and ion channels establish the resting potential of a neuron
• Every cell has a voltage (difference in electrical charge) across its plasma membrane called a
membrane potential
• The resting potential is the membrane potential of a neuron not sending signals
• Changes in membrane potential act as signals, transmitting and processing information
Formation of the Resting Potential
• In a mammalian neuron at resting potential, the concentration of K+ is highest inside the cell, while
the concentration of Na+ is highest outside the cell
• *Sodium-potassium pumps use the energy of ATP to maintain these K+ and Na+ gradients across the
plasma membrane
• *Operation of sodium-potassium pump moves: – Sodium ions out of the cell
– Potassium ions into the cell
• The opening of ion channels in the plasma
membrane converts chemical potential to electrical potential
• A neuron at resting potential contains many open K+ channels and fewer open Na+ channels; K+
diffuses out of the cell
• The resulting buildup of negative charge within the neuron is the major source of membrane potential
Key
Na
K
Sodium-potassium pump
Potassium channel
Sodium channel
OUTSIDE OF CELL
INSIDE OF CELL
Concept 48.3: Action potentials are the signals conducted by axons
• Changes in membrane potential occur because neurons contain gated ion channels that open or close in response to stimuli
• When gated K+ channels open, K+ diffuses out, making the inside of the cell more negative
• This is hyperpolarization, an increase in magnitude of the membrane potential
© 2011 Pearson Education, Inc.
• Opening other types of ion channels triggers a
depolarization, a reduction in the magnitude of the membrane potential
• For example, depolarization occurs if gated Na+ channels open and Na+ diffuses into the cell
• Graded potentials are changes in polarization where the magnitude of the change varies with the strength of the stimulus
• These are not the nerve signals that travel along axons, but they do have an effect on the
generation of nerve signals
© 2011 Pearson Education, Inc.
• *If a depolarization shifts the membrane potential sufficiently, it results in a massive change in
membrane voltage called an action potential
• Action potentials have a constant magnitude, are all-or-none, and transmit signals over long
distances
• *Action potentials are self-propagating
• They arise because some ion channels are
voltage-gated, opening or closing when the membrane potential passes a certain level
Action potential M em b ra n e p o te n ti a l ( m V ) Falling phase Rising phase
Threshold (55)
Depolarization Undershoot Resting potential Time (msec) 50 0 50
100 70
0 1 2 3 4 5 6
Generation of Action Potentials: A Closer Look
• An action potential can be considered as a series of stages
• At resting potential
1. Most voltage-gated sodium (Na+) channels are
closed; most of the voltage-gated potassium (K+) channels are also closed
OUTSIDE OF CELL
INSIDE OF CELL Inactivation loop
Sodium
channel Potassiumchannel
• When an action potential is generated
2. Voltage-gated Na+ channels open first and Na+
flows into the cell
3. During the rising phase, the threshold is
crossed, and the membrane potential increases 4. During the falling phase, voltage-gated Na+
channels become inactivated; voltage-gated K+
channels open, and K+ flows out of the cell
OUTSIDE OF CELL
INSIDE OF CELL Inactivation loop
Sodium
channel Potassiumchannel
OUTSIDE OF CELL
INSIDE OF CELL Inactivation loop
Sodium
channel Potassiumchannel
Action potential Threshold Resting potential Time M em b ra n e p o te n ti al (m V ) 50 100 50 0 Na K Key 2 1 3 1 2 3 Resting state Depolarization
Rising phase of the action potential
OUTSIDE OF CELL
INSIDE OF CELL Inactivation loop
Sodium
channel Potassiumchannel
Action potential Threshold Resting potential Time M em b ra n e p o te n ti al (m V ) 50 100 50 0 Na K Key 2 1 3 4 1 2 3 4 Resting state Depolarization
Rising phase of the action potential
Falling phase of the action potential
5. During the undershoot, membrane
permeability to K+ is at first higher than at rest,
then voltage-gated K+ channels close and
resting potential is restored
OUTSIDE OF CELL
INSIDE OF CELL Inactivation loop
Sodium
channel Potassiumchannel
Action potential Threshold Resting potential Time M em b ra n e p o te n ti al (m V ) 50 100 50 0 Na K Key 2 1 3 4 5 1 2 3 4 5 1
Resting state Undershoot Depolarization
Rising phase of the action potential
Falling phase of the action potential
• *During the refractory period after an action potential, a second action potential cannot be initiated
• The refractory period is a result of a temporary inactivation of the Na+ channels
© 2011 Pearson Education, Inc.
Conduction of Action Potentials
• At the site where the action potential is
generated, usually the axon hillock, an electrical current depolarizes the neighboring region of the axon membrane
• Action potentials travel in only one direction: toward the synaptic terminals
• Inactivated Na+ channels behind the zone of
depolarization prevent the action potential from traveling backwards
Evolutionary Adaptation of Axon Structure
• The speed of an action potential increases with the axon’s diameter
• In vertebrates, axons are insulated by a myelin sheath, which causes an action potential’s
speed to increase
• Myelin sheaths are made by glia—
oligodendrocytes in the CNS and Schwann cells in the PNS
Axon Myelin sheath
Schwann cell
Nodes of Ranvier
Node of Ranvier
Layers of myelin Axon
Schwann cell
Nucleus of Schwann cell
0.1 m
• Action potentials are formed only at nodes of Ranvier, gaps in the myelin sheath where
voltage-gated Na+ channels are found
• Action potentials in myelinated axons jump between the nodes of Ranvier in a process called saltatory conduction
Cell body
Schwann cell
Depolarized region (node of Ranvier)
Myelin sheath Axon
Concept 48.4: Neurons communicate with other cells at synapses
• At electrical synapses, the electrical current flows from one neuron to another
• At chemical synapses, a chemical
neurotransmitter carries information across the gap junction
• Most synapses are chemical synapses
• The presynaptic neuron synthesizes and packages the neurotransmitter in synaptic vesicles located in the synaptic terminal
• The action potential causes the release of the neurotransmitter
• The neurotransmitter diffuses across the
synaptic cleft and is received by the postsynaptic cell
*
Stages in transmission at a Chemical
Synapse
1. An Action Potential depolarizes the
membrane of the axon terminal.
2. Calcium ions rush into neuron’s cytoplasm.
3. The synaptic vesicles release
neurotransmitter into the synaptic cleft.
4. Neurotransmitter binds with receptors
Presynaptic
cell Postsynaptic cell
Axon
Presynaptic membrane
Synaptic vesicle containing
neurotransmitter Postsynapticmembrane Synaptic
cleft
Voltage-gated
Ca2 channel Ligand-gatedion channels
Ca2
Generation of Postsynaptic Potentials
• Direct synaptic transmission involves binding of neurotransmitters to ligand-gated ion channels
in the postsynaptic cell
• Neurotransmitter binding causes ion channels to open, generating a postsynaptic potential
• Postsynaptic potentials fall into two categories
– Excitatory postsynaptic potentials (EPSPs)
are depolarizations that bring the membrane potential toward threshold
– Inhibitory postsynaptic potentials (IPSPs) are
hyperpolarizations that move the membrane potential farther from threshold
Neurotransmitters
• There are more than 100 neurotransmitters, belonging to five groups: acetylcholine, biogenic amines, amino acids,
neuropeptides, and gases
• A single neurotransmitter may have more than a dozen different receptors
• *Cause a response at chemically-gated/ligand-gated ion channels found at synapse
Acetylcholine
• Acetylcholine is a common neurotransmitter in vertebrates and invertebrates
• It is involved in muscle stimulation, memory formation, and learning
• Vertebrates have two major classes of
acetylcholine receptor, one that is ligand gated and one that is metabotropic
Sensory and Motor Mechanisms
Chapter 50
Chapter 50
Overview: Sensing and Acting
• Sensory processes convey information about an animal’s environment to its brain, and muscles and skeletons carry out movements as instructed by
the brain
Concept 50.1: Sensory receptors transduce stimulus
energy and transmit signals to the central nervous system
• All stimuli represent forms of energy
• Sensation involves converting energy into a change in the membrane potential of sensory receptors
• When a stimulus’s input to the nervous system is processed a motor response may be generated
• This may involve a simple reflex or more elaborate processing
Sensory Pathways
• Sensory pathways have four basic functions in common
– Sensory reception – Tranduction
– Transmission – Integration
Sensory Reception and Transduction
• Sensations and perceptions begin with sensory reception, detection of stimuli by sensory
receptors
• Sensory receptors interact directly with stimuli, both inside and outside the body
• Sensory transduction is the conversion of
stimulus energy into a change in the membrane potential of a sensory receptor
• This change in membrane potential is called a
receptor potential
• Receptor potentials are graded potentials; their magnitude varies with the strength of the stimulus
Transmission
• After energy has been transduced into a receptor potential, some sensory cells generate the
transmission of action potentials to the CNS
• Some sensory receptors are specialized neurons while others are specialized cells that regulate
neurons
• Sensory neurons produce action potentials and their axons extend into the CNS
• The response of a sensory receptor varies with intensity of stimuli
• If the receptor is a neuron, a larger receptor
potential results in more frequent action potentials • If the receptor is not a neuron, a larger receptor
potential causes more neurotransmitters to be released
Figure 50.4
(a) Single sensory receptor activated Gentle pressure
Sensory receptor
More pressure
Low frequency of
action potentials per receptor
High frequency of
action potentials per receptor
(b) Multiple receptors activated
Sensory receptor Gentle pressure
More pressure
Fewer receptors activated
Types of Sensory Receptors
• Based on energy transduced, sensory receptors fall into five categories
– Mechanoreceptors – Chemoreceptors
– Electromagnetic receptors – Thermoreceptors
– Pain receptors
Mechanoreceptors
• Mechanoreceptors sense physical deformation caused by stimuli such as pressure, stretch,
motion, and sound
• The knee-jerk response is triggered by the
vertebrate stretch receptor, a mechanoreceptor that detects muscle movement
• The mammalian sense of touch relies on
mechanoreceptors that are dendrites of sensory neurons
Concept 50.2: The mechanoreceptors responsible for hearing and equilibrium detect moving fluid or settling
particles
• Hearing and perception of body equilibrium are related in most animals
• For both senses, settling particles or moving fluid is detected by mechanoreceptors
Hearing and Equilibrium in Mammals
• In most terrestrial vertebrates, sensory organs for hearing and equilibrium are closely associated in the ear
Outer ear Middle ear Inner ear Skull bone Malleus Incus Stapes Semicircular canals Auditory nerve to brain Cochlea Eustachian tube Round window Oval window Tympanic membrane Auditory canal Pinna 1 m
Bundled hairs projecting from a hair cell
Cochlear duct Bone Auditory nerve Vestibular canal Tympanic canal Organ of Corti Tectorial membrane Basilar membrane
Hair cells Axons of sensory neurons
To auditory nerve
Hearing
• Vibrating objects create percussion waves in the air that cause the tympanic membrane to vibrate • The three bones of the middle ear transmit the
vibrations of moving air to the oval window on the cochlea
• These vibrations create pressure waves in the fluid in the cochlea that travel through the
vestibular canal
• *Pressure waves in the canal cause the basilar
membrane to vibrate, bending its hair cells called sterocilia
• This bending of hair cells depolarizes the
membranes of mechanoreceptors and sends
action potentials to the brain via the auditory nerve
– *Cause ion channels to open or close
“Hairs” of hair cell Neurotrans-mitter at synapse Sensory neuron Action potentials Time (sec)
(a) No bending of hairs
S ig n al M em b ra n e p o te n ti a l (m V ) 70 50 0 70
0 1 2 3 4 5 6 7
70 50
0
70
0 1 2 3 4 5 6 7
70 50
0
70
0 1 2 3 4 5 6 7
Time (sec) Time (sec)
Receptor potential M em b ra n e p o te n ti al ( m V ) M em b ra n e p o te n ti al ( m V ) S ig n al S ig n al More neuro- trans-mitter Less neuro- trans-mitter
(b) Bending of hairs in one direction (c) Bending of hairs in other direction
Concept 50.3: Visual receptors on diverse animals depend on light-absorbing pigments
• Animals use a diverse set of organs for vision, but the underlying mechanism for capturing light is the same, suggesting a common evolutionary origin
Evolution of Visual Perception
• Light detectors in the animal kingdom range from simple clusters of cells that detect direction and intensity of light to complex organs that form
images
• Light detectors all contain photoreceptors, cells that contain light-absorbing pigment molecules
The Vertebrate Visual System
• In vertebrates the eye detects color and light, but the brain assembles the information and perceives the image
Figure 50.17a Sclera Suspensory ligament Cornea Iris Pupil Aqueous humor Lens
Vitreous humor Optic disk
Sensory Transduction in the Eye
• Transduction of visual information to the nervous system begins when light induces the conversion of cis-retinal to trans-retinal
• Trans-retinal activates rhodopsin, which activates a G protein, eventually leading to hydrolysis of
cyclic GMP
• When cyclic GMP breaks down, Na channels
close
• This hyperpolarizes the cell
• The signal transduction pathway usually shuts off again as enzymes convert retinal back to the cis
form
Figure 50.18 Light Inactive rhodopsin Active rhodopsin Transducin Phosphodiesterase
Concept 50.5: The physical interaction of protein filaments is required for muscle function
• Muscle activity is a response to input from the nervous system
• The action of a muscle is always to contract; extension is passive
Vertebrate Skeletal Muscle
• Vertebrate skeletal muscle moves bones and the body and is characterized by a hierarchy of
smaller and smaller units
• A skeletal muscle consists of a bundle of long fibers, each a single cell, running parallel to the length of the muscle
• Each muscle fiber is itself a bundle of smaller
myofibrils arranged longitudinally
• The myofibrils are composed of two kinds of myofilaments
– Thin filaments consist of two strands of actin and
two strands of a regulatory protein
– Thick filaments are staggered arrays of myosin
molecules
• Skeletal muscle is also called striated muscle
because the regular arrangement of myofilaments creates a pattern of light and dark bands
• The functional unit of a muscle is called a
sarcomere and is bordered by Z lines • *Sarcomere combine to make myofibrils
Figure 50.26
Muscle
Bundle of muscle fibers
Nuclei Single muscle fiber (cell)
Plasma membrane Myofibril
Z lines
Sarcomere
TEM
0.5 m
The Sliding-Filament Model of Muscle Contraction
• According to the sliding-filament model,
filaments slide past each other longitudinally, producing more overlap between thin and thick filaments
Figure 50.27
Sarcomere 0.5 m
Z M Z
Relaxed muscle
Contracting muscle
Fully contracted muscle
• The sliding of filaments relies on interaction between actin and myosin
• The “head” of a myosin molecule binds to an actin filament, forming a cross-bridge and pulling the
thin filament toward the center of the sarcomere • Muscle contraction requires repeated cycles of
binding and release
• Glycolysis and aerobic respiration generate the ATP needed to sustain muscle contraction
Thick filament Thin
filaments
Thin filament Myosin head
(low-energy configuration) ATP Thick filament Myosin-binding sites Actin
Myosin head (high-energy configuration
ATP
ADP P i
ADP
Cross-bridge Myosin head
(low-energy configuration) Thin filament moves
toward center of sarcomere.
P i
3 4
2 1
5
P i ADP
The Role of Calcium and Regulatory Proteins
• The regulatory protein tropomyosin and the
troponin complex, a set of additional proteins, bind to actin strands on thin filaments when a muscle fiber is at rest
• This prevents actin and myosin from interacting
Figure 50.29
Tropomyosin
Actin Troponin complex
Ca2-binding sites
(a) Myosin-binding sites blocked
Ca2
Myosin-binding site
• For a muscle fiber to contract, myosin-binding sites must be uncovered
• *This occurs when calcium ions (Ca2+) bind to the troponin complex and expose the myosin-binding sites
• Contraction occurs when the concentration of Ca2+ is high; muscle fiber contraction stops when the concentration of Ca2+ is low
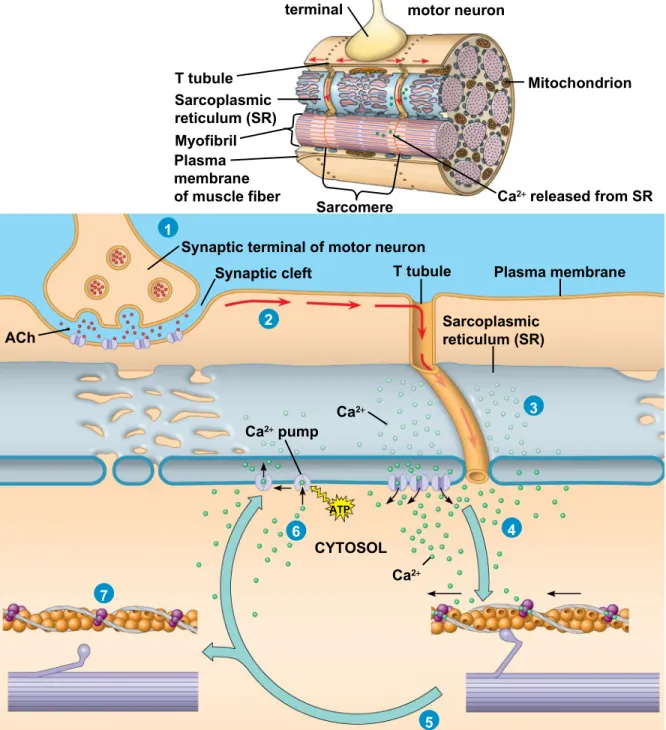
• The stimulus leading to contraction of a muscle fiber is an action potential in a motor neuron that makes a synapse with the muscle fiber
• The synaptic terminal of the motor neuron releases the neurotransmitter acetylcholine
• Acetylcholine depolarizes the muscle, causing it to produce an action potential
Figure 50.30 Synaptic terminal Axon of motor neuron T tubule Sarcoplasmic reticulum (SR) Myofibril Plasma membrane of muscle fiber
Sarcomere Ca2 released from SR Mitochondrion 2 1 3 4 5 6 7
Synaptic terminal of motor neuron Synaptic cleft T tubule
Sarcoplasmic reticulum (SR)
Plasma membrane ACh
Ca2 pump
Ca2
Ca2
CYTOSOL
Figure 50.30a
Synaptic terminal
Axon of
motor neuron
T tubule
Sarcoplasmic reticulum (SR) Myofibril
Plasma
membrane
of muscle fiber Sarcomere Ca2 released from SR
• Action potentials travel to the interior of the muscle
fiber along transverse (T) tubules
• The action potential along T tubules causes the
sarcoplasmic reticulum (SR) to release Ca2+
• *The Ca2+ binds to the troponin complex on the thin
filaments
• This binding exposes myosin-binding sites and allows the cross-bridge cycle to proceed
• Myosin heads perform a power stroke
2
Synaptic terminal of motor neuron Synaptic cleft T tubule
Sarcoplasmic reticulum (SR)
Plasma membrane
ACh
Ca2 pumpCa
2
Ca2
CYTOSOL
ATP
1
3
4
5 6
7
• When motor neuron input stops, the muscle cell relaxes
• Transport proteins in the SR pump Ca2+ out of the cytosol
• Regulatory proteins bound to thin filaments shift back to the myosin-binding sites
• Amyotrophic lateral sclerosis (ALS), formerly called Lou Gehrig’s disease, interferes with the
excitation of skeletal muscle fibers; this disease is usually fatal
• Myasthenia gravis is an autoimmune disease that attacks acetylcholine receptors on muscle fibers; treatments exist for this disease
Nervous Control of Muscle Tension
• Contraction of a whole muscle is graded, which means that the extent and strength of its
contraction can be voluntarily altered
• There are two basic mechanisms by which the nervous system produces graded contractions
– Varying the number of fibers that contract
– Varying the rate at which fibers are stimulated
• In vertebrates, each motor neuron may synapse with multiple muscle fibers, although each fiber is controlled by only one motor neuron
• A motor unit consists of a single motor neuron and all the muscle fibers it controls
Figure 50.31
Spinal cord Motor unit 1
Motor unit 2
Synaptic terminals
Nerve
Motor neuron cell body
Motor neuron axon
Muscle
Tendon
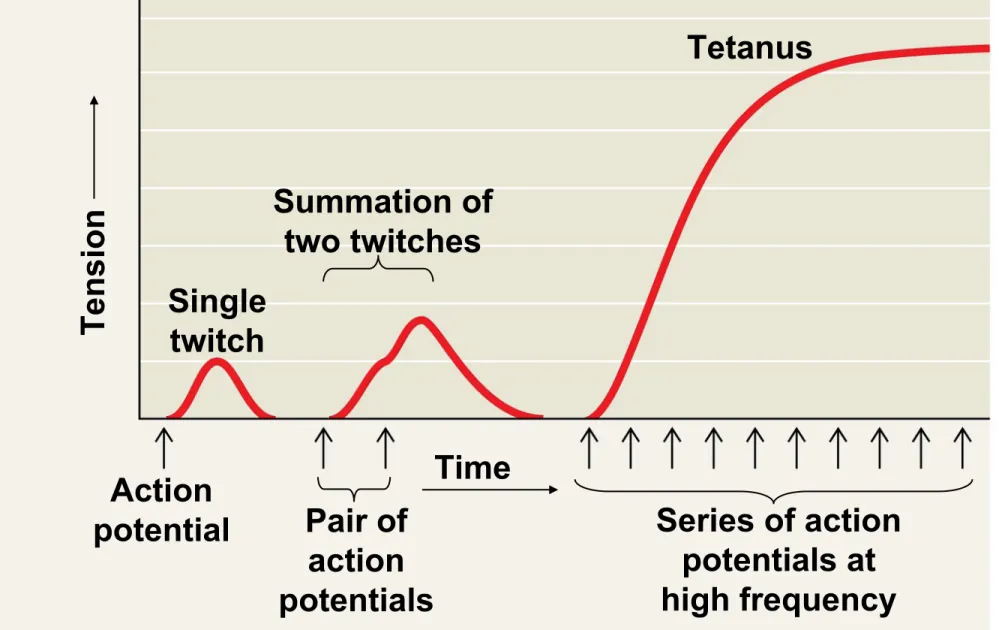
• Recruitment of multiple motor neurons results in stronger contractions
• A twitch results from a single action potential in a motor neuron
• More rapidly delivered action potentials produce a graded contraction by summation
Figure 50.32 Tetanus Summation of two twitches Single twitch Action
potential Pair of action potentials
• Tetanus is a state of smooth and sustained
contraction produced when motor neurons deliver a volley of action potentials
Types of Skeletal Muscle Fibers
• There are several distinct types of skeletal
muscles, each of which is adapted to a particular function
• They are classified by the source of ATP powering the muscle activity or by the speed of muscle
contraction
Oxidative and Glycolytic Fibers
• Oxidative fibers rely mostly on aerobic respiration to generate ATP
• These fibers have many mitochondria, a rich blood supply, and a large amount of myoglobin
• Myoglobin is a protein that binds oxygen more tightly than hemoglobin does
• Glycolytic fibers use glycolysis as their primary source of ATP
• Glycolytic fibers have less myoglobin than oxidative fibers and tire more easily
• In poultry and fish, light meat is composed of
glycolytic fibers, while dark meat is composed of oxidative fibers
Fast-Twitch and Slow-Twitch Fibers
• Slow-twitch fibers contract more slowly but sustain longer contractions
• All slow-twitch fibers are oxidative
• Fast-twitch fibers contract more rapidly but sustain shorter contractions
• Fast-twitch fibers can be either glycolytic or oxidative
– *Through conditioning fast glycolytic fibers can become fast oxidative fibers
Concept 50.6: Skeletal systems transform muscle contraction into locomotion
• Skeletal muscles are attached in antagonistic
pairs, the actions of which are coordinated by the nervous system
• The skeleton provides a rigid structure to which muscles attach
• *Skeletons function in support, protection, movement, and blood production