Microbiology
Highly localized tracks
of
human
immunodeficiency
virus type 1
Nef
in
the nucleus of
cells of a
human
CD4+
T-cell line
K. G. MURTI*tt, P. S. BROWN*, LEERATNER§, AND J. VICTOR GARCIA*
*DepamentofVirologyandMolecularBiology, St. Jude Children'sResearch Hospital, 332North Lauderdale, P.O. Box 318, Memphis, TN 38101-0318;
tDepartmentofPathology, University ofTennessee, TheHealth ScienceCenter,800Madison Avenue, Memphis, TN 38163; and§Divisionof
Hematology-Oncology, WashingtonUniversitySchool ofMedicine,660SouthEuclid Avenue,St.Louis, MO63110
CommunicatedbyDavid M. Prescott, September7, 1993 (receivedforreviewJuly6, 1993)
ABSTRACT A humanT-cell line constitutively expressing thenef gene from the humanimmunodeficiencyvirus
type
1 SF2 isolate wasused to examie the distribution of the Nef protein in the nucleus. High-resolution immunogoldlabel-ing/electronmicroscopic studies with polydonal anti-Nef an-tibodies onnef+ and nef- cells revealed that asmallfraction of Nef is in thenucleusanditislocalizedinspecificcurvilinear tracks that extend between the nuclear envelope and the nucleoplasm. An examination of the sequence of the SF2 nef gene revealed aputative nuclear targeting sequence that was previously found in several other eukaryotic nucleoplasmic proteins. The nuclear localizationofNef suggests a potential nuclear functionfor this protein. The presence of Nef in distinct nuclear tracks suggests that Nef is transported along aspecific
pathway that extends from the nuclear envelope into the nucleoplasm. A previous study [Meier, U. T. & Blobel, G. (1992)Cell 70, 127-138] has shown that the nucleolar protein ofrat livercells (Noppl40)
shutties
from thenucleolustothe nuclear envelope on distinct tracks. The present study has suggested that the transport of anucleoplasmic protein may also occur ondistinctnuclearpathways.The human immunodeficiency virus (HIV) is a complex
retrovirus that contains several genes that regulate virus
replicationand geneexpression(1-3).Theroleofoneofthese genes,nef,hasbeenthesubjectof a great deal of controversy
(4). Nef is encoded by a single open reading frame that
overlapswith the 3'longterminal repeatofHIVandsimian immunodeficiencyvirus (SIV) (4-6). Nefisa27- to32-kDa proteinthatismyristoylatedatits amino terminus(7-9). In some instances, a25-kDaproduct thatis notmyristoylated
has beenreported; however, thisform of Nef isnotalways
detectable(10).nefmRNAisdetectedalongwith thatcoding for tat and rev early after infection (11). The presence of
antibodies reactive with Nef inpatientsinfectedwithHIV or in macaquesinfectedwithSIV is evidence ofnef expression
in vivo(12-15).AlthoughNef is notrequired forreplication
in vitro, it is present in HIV-1, HIV-2, and SIV (4). The conservation of this gene in all three viruses canbe consid-ered an indication that nefmight have an effect on virus
replicationordiseaseprogression. Indeed, experiments con-ductedusingSIV(mac239isolate) indicatethatnef plays an
important role
in
the
development of disease in vivo (16).
A knowledge of the subcellular distribution of Nef may
provide clues to its function. Previous light microscopic
studies using immunofluorescence or immunoperoxidase techniques have found Nefprimarily in the cytoplasm (7, 17-19)associated with theplasmamembrane,
Golgi
complex,andendoplasmicreticulum.A recentbiochemical
study
has demonstrated that a substantial fraction of Nef of HIV-1 associates with the cytoskeletal fraction ofTlymphocytes
Thepublicationcostsofthisarticleweredefrayedin partbypagecharge
payment. This articlemustthereforebeherebymarked "advertisement"
in accordance with 18 U.S.C.§1734solelytoindicate thisfact.
and that theassociation is enhanced bymyristoylation ofNef (20). Thedistribution of Nef inthe nucleus remains contro-versial. Some recent studies using immunocytochemical techniques at the light microscopic level have described the
possible association ofNefwiththenuclearenvelope andthe nucleoplasm (18, 19, 21). In thisstudy we have attempted to resolve the question ofthe nuclear localization of Nef in human T cells using the high-resolution method of
immu-nogold/electronmicroscopy. The studies done with ahighly specificpolyclonal antiserum to Nef have revealed that a fraction of Nef islocalizedindistinct tracks in the nucleus.
MATERIALS ANDMETHODS
Cells and Antibodies. HPBALL human CD4+ T cells ex-pressing nef(HPBALL/LnefSN-S1) ortransduced with a control vector (HPBALL/LN) have been described (22). Cells were grown in RPMImedium containing25 mMHepes and supplementedwith 10% fetal bovine serum, penicillin, and streptomycin. A rabbit anti-Nef polyclonal antiserum (23) whosespecificitywasestablishedbyWestern blot
anal-ysis (see Fig. 1) was used in immunogold labeling studies. Monoclonal anti-vimentin (no. 814318) and anti-actin (no. 1378996) antibodies were obtained from Boehringer Mann-heim. Monoclonal anti-tubulin (no. MABO65) antibodies were purchased from Chemicon. The specificity of three
antibodies was established by immunofluorescence and/or
Western blot analysis.
Western BlotAnalysis. Todeterminethe specificity of the rabbit anti-Nef polyclonalantiserum,HPBALL/LnefSNor LNcellswerelysedin1%Nonidet P-40bufferandproteins
wereseparated bySDS/PAGEon a12.5%gel.Proteins were
transferredto anitrocellulosefllter, probedwith theanti-Nef antiserum (1:500 dilution), and developed with goat anti-rabbitalkaline phosphatase secondary antibody essentially
asdescribed (24).
Immunofluorescence and Immunogold Labeling.
Immuno-fluorescencewasperformedasdescribed(25). Briefly,cells were spun (Cytospin) onto glass slides, fixed with 3.7% paraformaldehyde, and permeabilized with acetone. Cells
werethenincubatedwith theprimaryantibodies [diluted 10-to20-foldwithphosphate-buffered saline (PBS)]at37°Cfor 1 hr. Afterthoroughrinsing, cells wereincubated asabove
withfluorescein-conjugatedgoatanti-rabbitantibodies(ICN; diluted 10-fold with PBS). The slides were viewed and
photographed inaZeiss IM35microscope.
Immunogoldlabelingwas doneas described(26,27)with
slightmodifications. Cellswerefixed in2.0% paraformalde-hyde/0.05% glutaraldehyde in 0.01 M sodium
cacodylate
buffer,dehydratedin
15%, 30%,
and50%ethanol for 15 mineach, and stained with2%uranylacetatein50%ethanol for 30 min. They were further
dehydrated
in 70% and 100%Abbreviations: HIV, humanimmunodeficiency virus; SIV, simian
immunodeficiencyvirus.
ITo whomreprintrequests should beaddressed.
Proc. Natl. Acad. Sci. USA 90(1993)
ethanol for 15 min each and immersed in 1:1 ethanol/LR White resin (Polyscience) for 1 hr. Finally, theywere
em-bedded in LRWhite resin for24hrat50°C. Sectionswerecut
withadiamondknifeon aSorvallMT6000 ultramicrotome
andpicked up onnickelgrids. Forantibody labeling, grids bearingsections were floated ondropsofprimaryantibodies
(anti-Neforanti-cytoskeleton) diluted10-to20-fold in
Tris-buffered saline (TBS, 500 mM
NaCl/25
mM Tris, pH 7.6) containing 0.1%fishgelatin and0.5% bovineserumalbumin.Incubation was carried out at 10°C for 16 hr. Grids were
rinsed with TBS and floated on gold-conjugated second
antibodies (Amersham) diluted 20-fold with TBScontaining fishgelatin and bovine serumalbumin. Incubation withthe
gold-conjugated antibodywas carried out at21°C for 3 hr.
After thorough rinsing, grids were stained with Reynold's lead citrate before electron microscopic examination. All
sampleswereexamined inaPhilips EM301 electron
micro-scopeoperatedat80kV.
RESULTS
NefExpression in HPB-ALLCells.We haveestablisheda
series ofcell populationsthatconstitutivelyexpressthenef
geneofHIV-1(SF2isolate;refs. 6 and 22). The human T-cell
line
HPBALL/LnefSN-S1
was chosen to determine theintracellular distributionof Nef because it is a human CD4+
T-cellline susceptibleto HIVinfection. Asshown inFig. 1,
HPBALL/LnefSN-S1 cells express a 27- to 29-kDaprotein
that reacts specifically with a rabbit anti-Nef antiserum. These cellswereoriginally isolated by fluorescence-activated
cellsortingonthebasis of their low CD4cell surface levels (22). Alow levelof surface CD4 expression correlates with Nefexpression andserves as anindicator of thepresenceof
afunctionalnefgene (22, 24).
Lcalizationof Nef by Immunofluorescence. Asapreludeto
the immunogold/electron microscopy studies to map the
subcellular localization ofNef, we performed
immunofluo-rescence studies with a polyclonal anti-Nef antiserum on
nef-
(HPBALL/LN)
andnef+
(HPBALL/LnefSN-Sl)
cells. As showninFig. 2, thecellsaregenerallyround andcontainedalargekidney-shaped nucleusthatfilledmostof the volume ofthecell. Thecytoplasmwaspolar andwas most
abundantnear anindentation ofthenucleus, typicallyseenin T cells. In nef+ cells, the anti-Nef antibody revealed a
z
C,)
4)
z z
-J -1
dom
FIG. 1. Specificityofanti-Nef rabbit polyclonalantiserum. Extracts from HP-BALLcellstransduced with LnefSN or
acontrol vector,LN,wereseparatedby
32.5
SDS/PAGE
on12.5%gels,and Nefwasdetectedby
Western
blotanalysiswitha -.--Nef rabbit anti-Nefspecific
antiserumusing=27.5
alkalinephosphatase-labeled
goat anti-rabbit antibodies (heavychain specific)asdescribed in the text. Thepositionof Nef onthe gel is indicated. Positions of prestained molecular mass standards are indicatedin kDa.
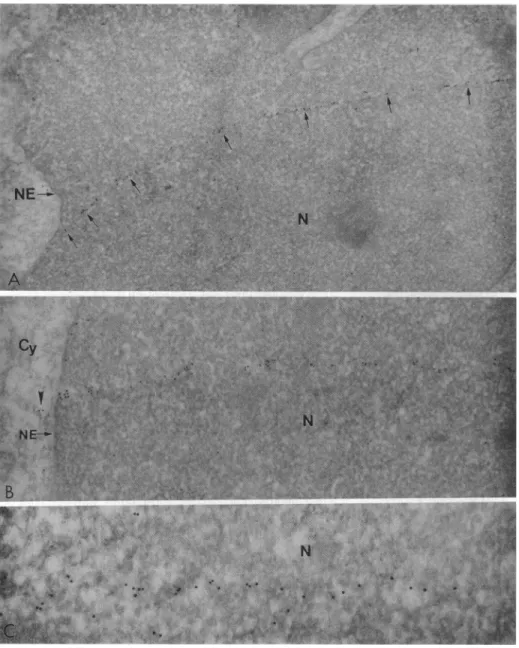
homogeneous labeling of the cytoplasm (Fig. 2A). Ingeneral, thedistribution of Nef seemspolar butthisevidently isdue tothepolar distribution of the cytoplasm; the thin ring of the cytoplasm surrounding the nucleus, when visible, also showedlabeling. Thenuclei, ingeneral, showed littleorno
fluorescence (Fig. 2A). However, focusing of the nucleiat different planes revealed narrow bands of fluorescence in somenuclei (Fig. 2B). These bands were seenin8ofthe 89 nuclei examined and each nucleus showed only one band. The bands are very faint and required long exposures to
photograph them. No fluorescence was observed in nefr cellsincubated withtheanti-Nefserum(Fig. 2C) orinnef+ cellsincubated withnormalrabbitserum(Fig.2D),although severalhundred of thesecells wereexamined.
NuclearLocalizaton of Nef byImmunogold Electron Micros-copy. Thetechnique used for immunogold labelingwas the
post-embeddingmethod(27). Fixed anddehydrated cellsare
embedded inawater-soluble embeddingresin (LRWhite)and
sectioned, andsectionsareincubated with primary and
sec-ondary (gold-conjugated) antibodies. We have maintained threesetsof controlstoensurethespecificity of the antibodies used. First, we performed the immunogold labeling with
anti-Nefantibodiesusing sections ofHPBALL/LN control cells(nef-). As shown in Fig. 3A, the nonspecific binding of theanti-Nefantibodies inthesecells wasnegligible. Second,
as a control for nonspecific binding of primary and
gold-conjugated secondary antibodies,weperformed immunogold labeling onnef+
(HPBALL/LnefSN-S1)
cells using normalFIG. 2. Immunofluorescence analy-sis of Nef+(HPBALL/LnefSN-S1)and Nef- (HPBALL/LN)cells with antise-rumagainstNef.(A) Nef+ cells labeled with the anti-Nef antiserum show label in thecytoplasm. (B)TheNefW cells labeled
asin Awerefocusedonthe interior of the nucleus. Noteafluorescentbandacross
the nucleus.(C) Nef- cells labeledwith the anti-Nef antiserumwerenotlabeled. (D)Nef+ cellslabeled with normal rabbit
serum werealsonotlabeled.(x800.)
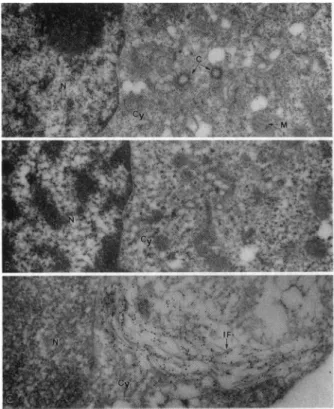
FIG. 3. Electron micrographs of controls for the immunogold labeling technique. (A)Nef- cells wereincubated with polyclonal
anti-Nefantiserum followedby gold-conjugated anti-rabbit antibody.
Labelingofcytoplasmic and nuclear componentsisnegligible. (B)
Nef+cellswereincubated withnormal rabbitantiserum followed by
goatanti-rabbitantibody conjugatedwithgold particles.No
nonspe-cifilc labelingof eithercytoplasmornucleus is evident.(C) Nef+ cells
were incubated with a monoclonal anti-intermediate filament
(vi-mentin) antibodyfollowedby gold-conjugated (anti-mouse) second
antibodies. The label is seen overintermediate filaments (IF). M,
mitochondrion; C, centriole; N, nucleus; Cy, cytoplasm. (A, x8100; B, x8550; C, x19,800.)
rabbit antiserumfollowedby gold-conjugated anti-rabbit
an-tibody.The resultsillustrated inFig.3B show that neitherthe rabbit serum proteins nor the secondary antibodies bind nonspecifically tonef+ cells. Finally, tocheck for the speci-ficity of labeling of subcellular structures by immunogold labeling method used here, nef+ cellswere labeled with an
anti-vimentin (intermediate filament) specific antibody. The intermediate filaments are readily identifiable cytoplasmic structuresthat provide convenient markers to testthe
reso-lution and specificity of immunogold labeling. The results illustrated inFig. 3C show the exclusivedistribution of label
overthe 10-nmintermediate filaments. Littleor nolabelingin the nucleus was observed in any of these controls. These studies suggest that the immunogold technique used here provides specific labelingof subcellular structures.
When T cells expressing Nef (HPBALL/LnefSN-S1)
were examined by the immunogold labeling method, the results were as follows. In afew sections of the nuclei the label due to Nefwasdetected inhighlylocalized tracks that extended between the nuclearenvelopeand thenucleoplasm. Fig. 4 illustrates these tracks in sections of three different nuclei(A-C).Thelongestof the tracks measured about7Am (Fig. 4A)and the tracksappearedtocommence/terminateat
the cytoplasmic side of the nuclearenvelope (Fig. 4B).The
tracks were seen in only4 nuclei among 100 examined and
serial sections revealed only one track per nucleus. It is
possiblethat these tracks mayoccurwithgreater frequency than that observed but that they are not detectable due to
technicalreasons.The tracksoccurin thin bands andoccupy
afraction of the total nuclear volume and, therefore, their detection would depend on their perfect alignment to the plane of sectioning. In addition to the tracks, a small amount of label is alsofound in thenucleoplasm but the nucleoliare totally free from the label. These tracks may correspond to the fluorescence bands observed in a few nuclei by the immunofluorescence method (Fig. 2B). A thorough exami-nation of sections of hundreds of nuclei of nef- cells incu-bated with anti-nef serum and nef+ cells incubated with normalserumhasfailed to reveal any nuclear labeling. In fact there is only one other instance in published literature concerning a nuclear protein that forms tracks in the nucleus. Astudy by Meier and Blobel (28) has shown that a nucleolar phosphoprotein (Noppl40) of rat liver cells shuttles on tracks that extend between the nucleolus and nuclear pore com-plexes. The Nef tracks are different from Noppl40 tracks in thattheytraversethenucleoplasm with no relationship to the nucleolus.
Inthecytoplasm(datanotshown) the label due to Nef was mostabundant near the indentation of the nucleus as was the case withcellsprocessed by immunofluorescence (see Fig.
2A);thisregioncontained mostof the cellorganelles,
includ-ing the microtubuleorganizing center, Golgi complex, and vesicles.
Dothe Nuclear TracksRepresent CytoskeletalFilaments? It has beenhypothesizedthat thetransportofRNA(29, 30) and
proteins (28)occurs onspecific tracksin the nucleus and that thesetracks maybecomposed of cytoskeletal filaments (e.g.,
microfilaments, microtubules,orintermediate filaments). To determine if any of these filaments form tracks within the nucleus ofTcells, wehaveconductedimmunogold labeling studies withanti-tubulin, anti-vimentin, and actin anti-bodies. The nuclei remained largely unlabeled with either anti-vimentin (Fig. 3C) oranti-tubulin antibodies (data not
shown).However, anti-actinantibodies showed some
label-ingof the nuclei(Fig.5). Although notasclear as Nef tracks, the label duetoactin showedapreferentialalignment in the nucleus. Additionally, short tracks of actin were seen
ex-tendingbetween thenuclearenvelope and the nucleoplasm
(Fig. 5).These resultsareconsistent withprevious biochem-ical evidence for the presence of actin in the nucleus (for
references, seeref. 28)anditsproposedrole in theshuttling ofproteinsbetween thenucleus and cytoplasm(28).
DISCUSSION
Thenefgeneis present inHIVand SIV and severalfunctions have beenascribedtoit. Thesefunctions include a negative effectonHIVreplicationin vitro(31-34),down-regulationof CD4 from the cell surface (8, 22, 24), and binding and
hydrolysis of GTP (35). Conflictingreports havealso been
published questioningthe role of Nef in each of the above functions(9, 36). Theissue of thebiologicalfunction of Nef is furthercomplicated bythefact thatisoformsof Nef exist thatdiffer either in the primaryamino acid sequence orin theirposttranslational modification (37)and these isoforms may havedifferent functions.
In an attempt to gain insights into the function of Nef,
earlier studies havefocusedonthe intracellularlocalization of Nefusing immunocytochemicalstudiesatthelight
micro-scopiclevel(7, 17-19, 21)andobtained varied results. Initial studies suggested an exclusive cytoplasmic distribution of Nef (7, 17), whereas three recent studies indicated the presence of Nef in the nucleus as well (18, 19, 21). In the present study our objective was to obtain a more precise
intracellular localization ofNef
using immunogold
labelingtechniquesat the electron microscopic level.
Thestudies described here have
important
implications
for thefunction of Nef in HIV-1 lifecycle
and for thegeneral
question
of theimport
ofproteins
into the nucleus.They
havey
Proc. Natl. Acad. Sci. USA 90(1993)
11898 Microbiology: Murtietal.
shownthatafraction of Nef ispresentin thenucleusand that it occurs in localized tracks within the nucleoplasm. The precise nuclear role of nuclear Nef remains unknown but
someprevious studies have suggested aregulatory rolefor
Nef in theHIVgeneexpression (31-34). Thelocalizationof
Nefindistinct tracksinthenucleoplasmissignificantinview of transportofproteinsintothenucleus. The firstexample of
a nuclear protein that occurs in tracks is the nucleolar
FIG. 5. Electronmicrographshowingimmunogold labelingofthe
nucleus ofNef+ cellswithamonoclonal anti-actin antibody.Inthe
nucleus the actin label shows preferentialalignment along tracks.
(Inset) Shorttrack of actin labelnearthe nuclearenvelope(NE). (x27,000;Inset, x46,800.)
FIG. 4. Electron
micrograph
showing immunogoldlabeling
of thenuclei of three different Nef+ cell.Noete
tracks ofgold par-~~. ~ ~ 5~E.tidles
extendingfrom the nuclearpt, -Tf
envelope
tothenucleoplasmin As andB.(A)Arrows markthetrack
~~~ ~~~~
~Of
Nef label.(B)The tracks ofNef ~~~~~~~appear
to originate in the cyto-plasm(arrowhead).(C)Partof the Nef track at high magnification.NE, nuclear
envelope;
Cy,cyto-plasm.
(A,
x23,800;
B,
x30,800;
C,
x56,700.)phosphoprotein
(Noppl40)
of rat livercells
(28).Noppl40
binds
nuclear
localization signal peptides and shuttles be-tween the nucleolus and cytoplasm. Immunogold labeling studies have shown that this protein occurs in tracks thatextendfrom the nuclearenvelopetothe
fibrillar
component ofthenucleolus. These results led theauthors (28)tosuggest thatNoppl40
shuttles on distinct tracks that may becom-posed of actin with the motive force for transport being
generated
by the putative nuclear myosinmotors.Although details ofthe mechanism ofnucleartransportremain toberesolved, it appears that most, if not all,
macromolecular
trafficincludingthatof mRNAmolecules (29, 30)proceeds
ondistinct nuclearpathways.Toourknowledge, demonstra-tionofanucleoplasmicprotein
ondistinct trackshasnotbeenreportedpreviously. TheNeftracks inthe nucleus remark-ably resemblethe
Noppl40
tracksand, likethe latter, mayrepresentnucleartransportpathways. The fact that thereis
onlyoneNeftrack pernucleus suggeststhat Nef traversesto thenucleus alongasingle
specific
pathwayinto the nucleo-plasm. Ashas beensuggested
beforeforNoppl40
(28),thespecificity
mayreside in the nuclear porecomplextowhichthe protein binds and to which the track is
presumably
anchored.
Previous studieshave shownthat the
eukaryotic
nucleus containsanorganized
matrixofproteinaceous
filament thatprovidesascaffoldfor the
organization
andfunctionof thenuclear components (for references, see ref. 38). Recent studies have shownthat various biochemical reactions, in-cluding DNA replication, transcription, and RNA
process-ing,take place ondefined regions ofthe nuclear matrix and the matrix may also provide tracks for the movement of proteins and RNA transcripts (28-30) in and out of the nucleus. Theidentity of proteins that compose the nuclear matrix remains tobe resolved. Some studies have implicated lamins (proteins of the fibrous lamina beneath the inner nuclearmembrane) in forming the nuclear matrix (39); lamins are related toproteins of cytoplasmicintermediate filaments and can formcoiled-coil rods, which, inturn, can associate toform higherorder structures (40). Another protein that has been detected in thenucleus (forreferences, see ref. 28) and isbelieved to composethe matrixis actin. It hasalso been speculated thatactin may provide tracks in the nucleus along whichmacromolecules move in and out of the nucleus (28). A recent study has demonstrated theaffinityof HIV-1 Nef to the cytoskeletalcomponents in vitro and in vivo (20). Allof
these findings led us to conduct apreliminary immunogold labeling study to determine if any of themajor cytoskeletal
proteins occur in tracks within the nucleus. The results
suggested that, among the cytoskeletal proteins, actin ap-pears to formshort trackswithin the nucleus.
Thelocalizationof Nef in the nucleus also led us to search for a nuclearlocalization signalin the nefsequence. Since
Nef is found in thenucleoplasm and not in thenucleolus,we focused on localization sequences on nucleoplasmic pro-teins. Previousstudies with Xenopus oocyte nuclearproteins (nucleoplasminand N1) have shown thatthese proteins share a bipartite nucleartargeting motif(41). Themotif consistsof a 16-amino acid sequence with two basic residues at the amino-terminalend, 10 "spacer"residues,andaclusterof4 basic residues at the carboxyl-terminal end (see below). Aminoacids inboth basic domains arerequired for nuclear
targeting and the transportdefect ofamutationinonedomain
is amplified by a simultaneous mutation in the other. In addition to Xenopus proteins, anumber ofeukaryotic and viral nuclearproteins contain this motif(41). The SF2 clone of Nef used here also shares thismotif(24).
Nucleoplasmin KRpaatkkagqaKKKK Thyroid al KRvakrklieqnReRRR
Nef-SF2 KRsmggwsaireRmRR
The presence of the nuclear targeting sequence may be required but notsufficient forthenuclear localization ofNef. Indeed, asubstantial fraction of Nef appears to be
cytoplas-mic associating with organelles and cytoskeletal elements,
suggesting that it may disrupthost cytoplasmicactivities. It is still possible that a fractionofNef hasanuclearfunction
that is yet to be identified.
We thank Mrs.GlenithNewberryfortypingthemanuscript.This work was funded in part by Grants CA-59195 from the National Cancer Institute and 1405 and 1559fromthe American Foundation for AIDS Research (J.V.G.), Grant FRA-373 from the American Cancer Society and Contract 17-90C-0125 from the U.S. Army (L.R.), Cancer CenterSupportGrant CA-21765 from the National Institutes of Health toSt. Jude Children's ResearchHospital,and the American Lebanese SyrianAssociated Charities.
1. 2. 3.
4. 5.
Culien,B. R. (1991)Annu.Rev. Microbiol. 45,219-250.
Pavlakis, G. N. &Felber,B. K. (1990)NewBiol.2,20-31.
Vaishnav, Y. N. & Wong-Staal,F.(1991)Annu. Rev.Biochem.60,
577-630.
Hovanessian,A. G.(1992)Res. Virol. 143,31-81.
Ratner, L.,Haseltine,W. A., Patarca, R., Livak,K. J., Starcich,
B., Josephs,S.F., Doran,E.A., Rafalski,J.A., Whitehorn,E. A.,
Baumeister, K., Ivanoff, L., Petteway, S.R., Jr., Pearson,M.I.,
Lautenberger, J. A., Papas,T.S.,Ghrayeb, J., Chang, N.,Gallo,
R.C. &Wong-Staal, F. (1985)Nature(London) 313, 277-284.
6. Sanchez-Pescador, R., Power,M.D.,Barr, P. J.,Steimer,K.S., Stempien,M.M.,Brown-Shimer,S.L.,Gee,W.W.,Renard, A., Randolph, A., Levy, J. A., Dina,D.&Luciw,P.(1985)Science
227, 484-492.
7. Franchini, G., Robert-Guroff, M., Ghrayeb, J., Chang, N. &
Wong-Staal,F.(1986) Virology155, 593-599.
8. Guy, B., Kieny,M.P.,Riviere, Y.,LePeuch,C.,Dott, K.,Girard, M.,Montagnier,L. &Lecocq, J.-P. (1987)Nature(London) 330, 266-269.
9. Hammes, S.,Dixon, E., Malim, M.,Cullen,B. &Greene,W.C. (1989)Proc.Natl.Acad. Sci. USA 86, 9549-9553.
10. Kaminchik, J., Bashan, N., Itach, A.,Sarver, N.,Gorecki,M. & Panet,A.(1991)J.Virol.65,583-588.
11. Robert-Guroff, M., Popovic, M., Gartner, S., Markham, P.,Gallo,
R.C. &Reitz,M. S.(1990)J.Virol. 64, 3391-3398.
12. Ameisen, J. C.,Guy, B., Chamaret, S.,Loche,M., Mouton,Y., Neyrinck,J.L.,Khalife, J., Leprevost,C.,Beaucaire, G., Boutil-Ion,C.,Gras-Masse,H.,Maniez,M.,Kieny,M.P.,Laustriat, D., Berthier, A., Mach, B.,Montagnier, L., Lecoq, J. P. &Capron,A.
(1989) AIDSRes.Hum. Retroviruses5, 279-291.
13. Gombert,F.O.,Blecha, W., Tahtinen, M.,Ranki, A.,Pfeifer,S., Troger,W., Braun, R.,Muller-Lantzsch,N.,Jung,G., Rubsamen-Waigman,H.&Krohn,K.(1990)Virology176,458-466. 14. Kirchhoff, F., Vos,G., Nick, S.,Stahl-Hennig,C.,Coulibaly, C.,
Frank, R.,Jentsch,K.D. &Hunsmann, G.(1991) Virology 183,
267-272.
15. Sabatier, J. M., Clerget-Raslain, B., Fontan,G., Fenouillet, E., Rochat, H., Granier, C., Gluckman, J.C., VanRietschoten, J., Montagnier,L.&Bahraoui,E.(1989)AIDS3, 215-220.
16. Kestler,H.W.,Ringler, D. J., Mori, K.,Panicali,D.L.,Shehgal,
P.K.,Daniel,M. D.&Desrosiers,R.C.(1991)Cell65, 651-662.
17. Hammes, S.R., Dixon, E.P., Malim, M.H., Cullen, B. R. & Greene,W.C.(1989)Proc.Natl.Acad. Sci. USA86,9549-9553.
18. Ovod, V., Lagerstedt, A., Ranki, A.,Gombert,F.O.,Spohn, R., Tihtinen,M.,Jung, G. &Krohn,K. J. E.(1991)AIDS6,25-34. 19. Kohleisen, B., Neumann, M., Herrman, R., Brack-Werner, R.,
Krohn,K.J.E.,Ovod, V., Ranki,A.&Erfle,V.(1992)AIDS6, 1427-1436.
20. Neiderman,T. M. J.,Hastings,W. R.& Ratner,L.(1993)Virology
197,420-425.
21. Kienzle, N.,Bachmann,M.,Muller,W.E.G. &Muller-Lantzsch,
N.(1992) Arch. Virol. 124,123-132.
22. Garcia,J. V.&Miller,A. D.(1991)Nature(London)350,508-511.
23. Niederman,T. M.J.,Garcia,J.V.,Hastings,W.R.,Luria,S. & Ratner,L.(1992)J.Virol.66,6213-6219.
24. Anderson, S., Shugars, D. C.,Swanstrom,R.&Garcia,J.V.,J.
Virol.67,4923-4931.
25. Murti,K.G., Kaur,K. &Goorha, R.(1992) Exp. CellRes.202,
36-44.
26. Murti, K.G., Brown,P.S.,Bean,W.J.,Jr., &Webster,R.G. (1992)Virology186,294-299.
27. Murti,K.G.,Davis,D. S. &Kitchingman,G.(1990)J.Gen.Virol.
71,2847-2857.
28. Meier,U. T.&Blobel, G. (1992) Cell 70, 127-138.
29. Lawrence,J.B.,Singer,R.H. &Marselle,L. M.(1989) Cell57,
493-502.
30. Huang, S. &Spector,D. L.(1991)Genes Dev.5, 2288-2302.
31. Niederman, T.,Thielan,B.&Ratner,L.(1989)Proc.Natl. Acad.
Sci. USA86,,1128-1132.
32. Ahmad,N.&Venkatesan, S.(1988)Science241, 1481-1485.
33. Cheng-Mayer, C., Iannello, P., Shaw, K.,Luciw, P. &Levy,J. (1989)Science246, 1629-1632.
34. Luciw,P. A.,Cheng-Mayer,C. &Levy, J.A.(1987)Proc.Natl.
Acad. Sci. USA84,1434-1438.
35. Nebreda,A.R.,Bryan,T.,Segade, F.,Wingfield, P., Venkatesan, S. &Santos,E. (1991) Virology183,,151-159.
36. Kim, S., Ikeuchi, K., Byrn, R.,Groopman, J. &Baltimore, D.
(1989)Proc.Natl. Acad.Sci. USA 86,9544-9548.
37. Ratner, L., Starcich, B., Josephs, S. F., Hahn, B.H., Reddy,
E. P.,Livak, K.J., Petteway, S.R.,Jr., Pearson,M.L.,
Hasel-tine,W.A., Arya, S. K. &Wong-Staal, F. (1985)Nucleic Acids
Res. 13,8219-8229.
38. Mirzayan, C.,Copeland,C.S. &Snyder,M.(1992)J.CellBiol.116,
1319-1332.
39. Gerace, L.,Blum,A.&Blobel,G.(1978)J.CellBiol.79,546-566. 40. Aebi, V., Cohn,J.,Buhle,L.&Gerace,L.(1986)Nature(London)
323, 560-564.
41. Robbins,J.,Dilworth,S.M.,Laskey,R.A.&Dingwali,C.(1991)