Receptor Associated Kinase (IRAK-1) in IL-ip Signal
Transduction to NFk
B.
Emma-Louise Cooke
A dissertation submitted for the degree o f Doctor o f Philosophy to University College London at the University o f London.
June 2001
Department of Molecular Pathology, Windeyer Institute of Medical Sciences,
University College London, 46 Cleveland Street,
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest U643383
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
The experimental work described in this thesis was carried out in the Department of Molecular Pathology,
University College London and at GlaxoSmithKline Medicines Research Centre, Stevenage, Hertfordshire
between September 1997 and June 2001.
This thesis is the result of my own work and includes nothing that is the outcome of work done in
Kinase (IRAK-1) in IL -lp Signal Transduction to NF
kB.
Emma-Louise Cooke
Abstract
The IL-1 receptor-associated kinase (IRAKI) is essential for IL-1-stimulated NFicB-dependent gene
activation. To study the role of IRAK-1 in IL-ip signalling, we have generated a set of IRAK-1 variants
that express distinct domains of IRAK-1 either alone or in combination and have examined their effects on
an NFkB responsive reporter in HeLa cells. Unlike full-length lRAK-1, the deletion mutants were unable to activate N FkB in the absence of cytokine stimulation. However, an lRAK-1 variant lacking only the N-
terminal domain retained the ability of the full-length protein to potentiate both IL-ip and TNFa-induced
N FkB activation. In contrast, expression of the N-terminus or the C-terminus of lRAK-1, or a fusion protein incorporating both domains inhibited both IL-ip and TNFa-induced effects. Expression of an
lRAK-1 variant lacking only the C-terminal domain preferentially inhibited IL-ip versus TNFa-induced NF
kB activation. These data suggest that the C-terminal domain may link lRAK-1 to downstream signalling components common to both the lL-1 and TNF pathways. We have demonstrated that endogenous lRAK-1
becomes phosphorylated upon IL-ip treatment and can be detected along with NEMO and IKKP in high
molecular weight complexes of 600-800kDa. Moreover, lRAK-1 could be detected in NEMO
immunoprecipitates from IL-ip stimulated cells, suggesting that NEMO recmits lRAK-1 into the IkB
kinase complex. Additionally, we have identified proteins that potentially interact with the C-terminus of
lRAK-1 by utilising the yeast 2-hybrid system. A novel protein, which we called IRIP (IRAK-1-interacting
protein) was identified. Bioinformatic analysis suggests that IRIP is a member of a family of ecto-
nucleotide pyrophosphatase/phosphodiesterases and is likely to be the human homologue of a yeast protein
called GP17, which is involved in the addition of side chains to GPl anchors. It remains to be established
whether IRIP has phosphodiesterase/pyrophosphatase activity and also what role this protein may play in
I would like to take this opportunity to say a massive “thankyou” to all the people who have helped me
throughout my PhD. My warmest gratitude goes to my supervisor Keith Ray for his unrelenting support, for
all his helpful ideas, constructive criticism and encouragement during my project. I couldn’t have done it
without you! Thanks also to Iain for his technical know-how, advice, abuse and for keeping me sane (just
about!). I would also like to thank Pat Woo and Chulin Xia at University College London for their support.
At GlaxoSmithKline, special thanks goes to Alan Lewis for all his patience and help with the bioinformatics
and of course, thanks to everyone in the Cell Biology Department who have helped me and made my three
years at Glaxo enjoyable. In particular, thanks to Debs for the Friday night beers and numerous cups of hot
ribena to calm my nerves! Thanks also goes to Catherine, who has had the joyous task of proof-reading my
thesis - 1 owe you a few beers!
To my dear friends Kirsty, Sharon, Pritash and Clair, who are always there for me in times of crisis and of
course, are always eager to celebrate the good times!! You’re the best. Not forgetting Andy, thanks for
being the eternal optimist and always managing to say the right thing - how do you do that?!
Most of all I would like to thank my family, especially my Mum, Steve, Mark, Grandad and Joan, for always
TITLE I
DECLARATION III
ABSTRACT IV
ACKOWLEDGEMENTS V
LIST OF FIGURES XIII
ABBREVIATIONS XVI
CHAPTER 1 INTRODUCTION 1
1.1 OVERVIEW OF INNATE AND ADAPTIVE IMMUNITY 1
1.2 INTERLEUKIN-1 (IL-1) 6
1.2.1 Biological Effects of IL-1 6
1.2.2 Molecular Nature of IL-1 8
1.3 THE IL-1 RECEPTOR/TOLL-LIKE RECEPTOR SUPERFAMILY 11
1.4 IL-1 RI/TOLLR-MEDIATED SIGNALLING PATHWAYS 15
1.4.1 Clues from the Drojop/îiZa Toll Pathway 16
1.4.2 Early IL-1 R Signal Transduction Events 19
1.4.2.1 Recruitment of IRAK-1 into the IL-1 RI Complex 19
1.4.2.2 The Role of the Kinase Activity of IRAK-1 21
1.5 TUMOUR NECROSIS FACTOR (TNF) 23
1.5.1 The Biological Effects of TNF 23
1.6 THE TNF RECEPTOR SUPERFAMILY 25
1.7 INTRACELLULAR SIGNALLING IN RESPONSE TO TNF 28
1.7.1 Death Induction by TNF 28
1.7.2 Signalling by TNFRl 28
1.7.3 Induction of NFkB and Antiapoptotic and Proinflammatory Signals by TNF 30
1.9 INDUCIBLE TRANSCRIPTION FACTORS AND THE BASAL TRANSCRIPTION 34
MACHINERY
1.10 THE NFkB FAMILY OF TRANSCRIPTION FACTORS 36
1.10.1 Control of Transcription Initiation by the IkB Family of Proteins 39
1.10.1.1 The IkB Proteins: Structure and Function 39
1.10.1.1 Specificity of the IkB Proteins for RoI/NFkB Family Members 40
1.10.1.2 Structure of the IkBœ/NFkB Complex 41
1.10.1.3 Activation and Degradation of the IkBs 43
1.11 THE IkB KINASE (IKK) COMPLEX - A KEY REGULATOR OF NFkB ACTIVATION 45
1.11.1 The IkB Kinases: Structure and Function 46
1.11.2 NEMO (IKKy/IKAPl) 47
1.11.3 Physiological Functions of the IKKs 49
1.11.4 Inactivation of the IkB Kinase (IKK) Complex 50
1.12 THE UBIQUITIN SYSTEM 53
1.12.1 The Role of the Ubiquitin-proteasome Pathway in the Degradation of IxBa, (3-catenin and 54
the Processing of Cubitus Interruptus (Ci)
1.12.1.1 The Hedgehog (Hh) and WntAVingless (Wg) Signalling Pathways 54
1.12.1.2 The NFkB Signalling Pathways 55
1.12.2 Degradation of Ubiquitin-Protein Conjugates 58
1.12.3 The Regulatory Role Played by Ubiquitin in IKK Activation 59
1.13 AIMS 60
CHAPTER 2 MATERIALS AND METHODS 61
2.1 BACTERIAL CELL CULTURE AND TRANSFORMATION 61
2.1.1 Bacterial Growth Medium 61
2.2 PLASMID MANIPULATION 63
2.2.1 Isolation of Plasmid DNA by the Alkaline Lysis Method 63
2.2.1.1 Small Scale Purification of Plasmid DNA (miniprep) 64
2.2.1.2 Large Scale Purification of Plasmid DNA (maxiprep) 64
2.2.2 Caesium Chloride (CsCl) Purification of Plasmid DNA 65
2.2.3 Quantitation of DNA 66
2.2.4 Restriction Enzyme Digestion of DNA 67
2.2.5 Agarose Gel Electrophoresis 67
2.2.6 Gel Purification of DNA Fragments 68
2.2.7 Phosphatase Treatment of DNA 69
2.2.8 Ligation of DNA 69
2.2.9 DNA Sequencing 69
2.2.10 Amplification of DNA by the Polymerase Chain Reaction (PCR) 71
2.2.10.1 PCR Primers 72
2.2.10.2 PCR Conditions 72
2.2.11 Quikchange Site-Directed Mutagenesis 73
2.2.12 Purification of PCR Products 73
2.3 ROUTINE YEAST CELL CULTURE 74
2.3.1 Yeast Growth Medium 74
2.3.2 Yeast Two-Hybrid Vectors 75
2.3.3 Yeast Strains 76
2.4 ESTABLISHMENT OF A Y190 REPORTER STRAIN EXPRESSING THE GAL4p 77
DNA-BINDING DOMAIN FUSED TO THE GENE OF INTEREST
2.4.1 Small Scale Preparation of Competent Cells 77
2.4.2 Small Scale Plasmid Transformation of Yeast 78
2.5.2 Library Transformation and Selection 80
2.5.3 Screening Using the X-Gal ^-Galactosidase Colony-Lift Filter Assay 81
2.5.4 Plasmid Isolation From Yeast 82
2.5.5 Plasmid Rescue Via Complementation of E.co/i Mutations 82
2.6 ROUTINE MAMMALIAN CELL CULTURE 83
2.6.1 General Maintenance of Cell Lines 83
2.6.2 Mammalian Expression Plasmids 84
2.7 TRANSIENT TRANSFECTION OF HELA CELLS BY CALCIUM PHOSPHATE 86
PRECIPITATION AND REPORTER GENE ASSAYS
2.8 PROTEIN EXPRESSION AND ANALYSIS 87
2.8.1 Preparation of Protein Samples 87
2.8.2 Protein Estimation 88
2.8.3 Phosphatase Treatment of Cell Extracts 88
2.8.4 Polyacrylamide Gel Electrophoresis (PAGE) 89
2.8.5 Staining of SDS-Polyacrylamide Gels with Coomassie Brilliant Blue 90
2.8.6 Drying of SDS-Polyacrylamide Gels 90
2.8.7 Western Blotting 91
2.8.8 Immunological Deteetion of Proteins 92
2.8.8.1 Antibodies Used for Western Blotting 92
2.8.9 Stripping of Nitrocellulose Membranes 93
2.8.10 Immunoprécipitation of Protein Complexes 93
2.9 SIZE EXCLUSION CHROMATOGRAPHY 94
ASSOCIATED KINASE (IRAK-1)
3.2 DEFINITION AND ANALYSIS OF THE STRUCTURAL DOMAINS WITHIN IRAK 97
3.2.1 Structural Properties of the Kinase Domain 97
3.2.2 The Death Domain 104
3.2.3 The C-Terminal Region 107
3.2.4 Relationship of IRAK-1 to Other Kinases 111
3.3 CLONING OF THE IRAK I VARIANTS 114
3.3.1 Mammalian Expression Vectors Used for Cloning 115
3.3.2 Generation of the [IRAK 1-K(212-532)], [IRAK1-C(533-712) and [IRAKI-KC(212-712)] 115
Constructs
3.3.3 Generation of the [IRAK1-FL(1-712)], [IRAK1-N(1-211)], [IRAKI-NK( 1-532)] and the 118
[IRAK 1 -NC( 1-211:533-712)] Constructs
3.4 TRANSFECTION STUDIES 119
3.4.1 Analysis of Data 119
3.4.2 General Transfection Approach 120
3.4.3 Time-Course of NFkB Activation Following Treatment of cells with IL-1|3 or TNFa 121
3.5 VALIDATION OF EXPRESSION 122
3.5.1 Validation of Expression Revealed a Problem with the pBKRSV Constructs 122
3.5.2 Modification of the Commercial pBKRSV Vector for Mammalian Cell Expression 125
3.5.3 Expression of the IRAK I Constructs Using the Modified pBKRSV Vector in 125
Mammalian Cells
3.6 NFkB REPORTER GENE ASSAYS 129
3.6.1 Full-length IRAK I Activates NFkB Upon Cytokine Stimulation 129
3.6.2 Full-length IRAK-1 and an NHz-Terminally Truncated Version of IRAK-1, 129
the N- and C-terminal Domains, [IRAK1-NC(1-211:533-712)], Inhibited NFkB Activation
Following Cytokine Stimulation
3.6.4 Expression of an IRAK-1 Variant Lacking the C-terminus, [IRAKl-NK(l-532)], 134
Preferentially Inhibited IL-ip-Driven NFkB Activation
3.7 DISCUSSION 134
CHAPTER 4 IL-1 p-DEPENDENT RECRUITMENT OF IRAK-1 INTO THE IkB KINASE 144
COMPLEX
4.1 INTRODUCTION 144
4.2 OBJECTIVES OF THE GEL FILTRATION AND IMMUNOPRECIPITATION STUDIES 145
4.3 SIZE EXCLUSION CHROMATOGRAPHY 146
4.3.1 Gel Filtration Column Chromatography 146
4.3.2 Analysis of Data 146
4.4 IL-1 p-STIMULATED MIGRATION OF ENDOGENOUS IRAK-1 INTO THE 147
IkB k in a s e (IKK) COMPLEX
4.4.1 Analysis of Endogenous IRAK-1-Containing Complexes in Early IL-ip Signal 147
Transduction Using Size Exclusion Chromatography
4.4.2 IL-ip-dependent Association of Endogenous IRAK-1 and NEMO in HeLa Cells 152
4.5 DETERMINATION OF THE ELUTION PROFILES OF THE [IRAK1-FL(1-712)], 154
[IRAKI-KC(212-712)], [IRAKI-0(533-712)] AND [IRAKl-NK(l-532)] VARIANTS
BEFORE AND FOLLOWING IL-lp STIMULATION USING SIZE EXCLUSION
CHROMATOGRAPHY
4.5.1 IL-ip-dependent Migration of the [IRAK 1 -KC(212-712)] and [IRAKI-0(533-712)] 154
Variants Into the IKK Complex
4.5.2 IL-ip-dependent Migration of the [IRAKl-NK(l-532)] Variant Into the IKK Complex 155
4.6 DISCUSSION 161
CHAPTER 5 IDENTIFYING PROTEIN-PROTEIN INTERACTIONS USING THE 168
YEAST TWO-HYBRID (Y2H) SYSTEM
5.1 THE BASIC PRINCIPLE OF TWO-HYBRID SCREENING 168
5.1.1 Origins of the Technique 168
5.1.2 Searching for Interacting Proteins Using the Two-Hybrid System 168
5.1.3 Two-Hybrid Vectors and Yeast Strains 170
2.1.4 Limitations of the Two-Hybrid System 171
5.2 OBJECTIVES OF THE YEAST TWO-HYBRID STUDIES 171
5.3 ESTABLISHING A REPORTER STRAIN EXPRESSING THE GAL4p DNA-BINDING 173
DOMAIN FUSED TO THE C-TERMINAL REGION OF IRAK-1
5.3.1 Construction of a Hybrid Gene Encoding the GAL4p DNA Binding Domain Fused to IRAK I 173
5.3.2 Verification of Expression Products 175
5.4 TWO-HYBRID SCREEN OF A HELA CELL cDNA LIBRARY 177
5.4.1 Evaluation of Controls 177
5.4.2 Screening of the HeLa Cell cDNA Library 179
5.4.2.1 Verification of Two-Hybrid Responses 179
5.4.2.2 Positives Identified By the Two-Hybrid Screen 183
5.5 IDENTIFICATION OF IRAKl-INTERACTING PROTEIN (IRIP) 185
5.5.1 Sequence and Structure of IRIP and its Related Proteins 177
5.5.2 Relationship of IRIP to the E-NPP family of Ecto-nucleotide Pyrophosphatase/ 194
Phosphodiesterases
5.6 IDENTIFICATION OF THE BETA 2 TUBULIN GENE 199
FACTOR NF-IL6
5.8 OTHER IRAK-CT-INTERACTING PROTEINS 208
5.9 DISCUSSION 210
CHAPTER 6 FINAL DISCUSSION
REFERENCES
217
231
List of Figures
FIGURE 1-1 Human Blood Monocyte Producing IL-1 a, IL-1|3, IL-18 and IL-lRa
FIGURE 1-2 The Currently Known Members of the IL-IR/TLR Superfamily
FIGURE 1-3 Conserved Signalling Pathways Involved in the Activation of Dorsal and Dif
in Drosophila and of N FkB in Mammals
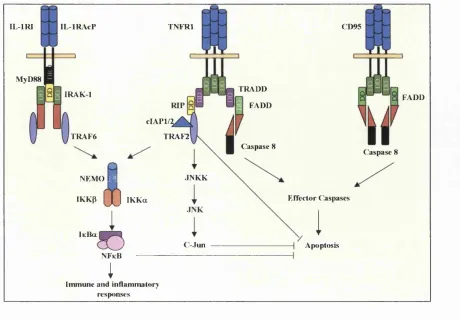
FIGURE 1-4 IL-ip, TNFa and CD95L Mediated Signalling Pathways Leading to Activation
of the Transcription Factors NFkB, AP I or Apoptosis FIGURE 1-5 Structure of the TRAF Family of Proteins
FIGURE 1-6 Model for the Initiation of Transcription by RNA Pol II
FIGURE 1-7 The Most Common Vertebrate IkB Proteins
FIGURE 1-8 Schematic Representation of the lKB/p65/p50 Crystal Structure
FIGURE 1-9 Alignment of the N-terminal Regulatory Regions of the IkB ’sand Cactus
FIGURE 1-10 Components of the IkB Kinase Complex
FIGURE 1-11 Model for Regulation of IkB Kinase Activity by Phosphorylation
FIGURE 1-12 Schematic Representation of the Ubiquitination of Phosphorylated IkB a by the SCFp '^*^^^ Complex
FIGURE 1-13 The 26S Proteasome
10
13
19
24
32
35
38
42
44
48
52
57
FIGURE 2-3 Mammalian Expression Vectors 85
FIGURE 2-4 Prestained Molecular Weight Markers 90
FIGURE 2-5 Transfer of Proteins to a Nitrocellulose Membrane 91
FIGURE 3-1 Predicted Domain Structure of IRAK-1 (Accession Number L76191) 98
FIGURE 3-2 Amino Acid Sequence of IRAK-1 (Accession Number L76191) 99
FIGURE 3-3 The Six-Helix Bundle Fold of Pelle-DD and Tube-DD and sequence Alignments 105
of Death Domains
FIGURE 3-4 Sequence Alignment of IRAK-1 Family Members 108
FIGURE 3-5 Sequence Alignment of the C-terminal Regions of IRAK Family Members 110
FIGURE 3-6 IRAK-1 Variants 114
FIGURE 3-7 The pBKRSV Vector 116
FIGURE 3-8 NFkB Activation in Mock HeLa Cells Transfected with the Reporter Genes Alone 120
FIGURE 3-9 Time Course of NFkB Reporter Gene Activation in response to HeLa Cell 123
Stimulation by IL-1 P and TNFa
FIGURE 3-10 Expression of the IRAK-1 Variants 124
FIGURE 3-11 Expression and Phosphorylation of the IRAK I Variants in the Presence and 127
Absence of Cytokine Stimulation
FIGURE 3-12 Effects of Protein Phosphatase-2A (PP-2A) on Mobility of IRAK-1 Immunoreactive 128
Bands
FIGURE 3-13 Effects of Full-length Wild-Type IRAK-1 and the IRAK I Variants on NFkB 130
Reporter Gene Activity in the Absence of Cytokine Stimulation
FIGURE 3-14 The Effects of Full-Length IRAK I and the IRAK I Variants on Cytokine Stimulated 131
NFkB Reporter Gene Activity
FIGURE 3-15 The Effects of the IRAK I Variants on Cytokine Stimulated NFkB Reporter Gene 133
FIGURE 4-3 Gel Filtration Analysis of the IkB Kinase Complex in HeLa Cells 151
FIGURE 4-4 Endogenous IRAK I and NEMO Coimmunoprecipitate Upon IL-ip Stimulation 153
FIGURE 4-5 Gel Filtration Analysis of [IRAK1-C(533-712)] in HeLa Cells 156
FIGURE 4-6 Gel Filtration Analysis of [IRAKI-KC(212-712)] in HeLa Cells 157
FIGURE 4-7 Gel Filtration Analysis of [IRAKl-NK(l-532>] in HeLa Cells 158
FIGURE 4-8 Gel Filtration Analysis of [IRAK 1 -FL( 1-712)] in HeLa Cells 160
FIGURE 5-1 A Schematic Representation of the Two-Hybrid System 169
FIGURE 5-2 Construction of a Hybrid Protein Encoding the Gal4p DNA-Binding Domain 174
Fused to IRAK I
FIGURE 5-3 Western Blot Showing Increasing Expression of the pBDGAL4Cam-IRAKlCT 176
Hybrid Protein in Y190 Cells
FIGURE 5-4 Controls for Two-Hybrid Screen 178
FIGURE 5-5 Restriction Digestion Analysis of the Positive cDNA Library Clones Identified in 181
the Primary Two-Hybrid Screen
FIGURE 5-6 Filter Lift Assay from the Secondary Screen 182
FIGURE 5-7 Deduced Protein Sequence of IRIP 186
FIGURE 5-8 Sequence Alignment of Members of the IRIP Family 187
FIGURE 5-9 A Phylogenetic Plot of the IRIP Family Members 190
FIGURE 5-10 Schematic Representation of the Predicted Structure of the IRIP-Related Integral 192
Transmembrane Proteins
FIGURE 5-11 Multiple Hydropathy Plot of the IRIP-Related Proteins 193
FIGURE 5-12 Sequence Alignment of Members of the IRIP Family with the E-NPP Family 196
FIGURE 5-13 The Structure of the E-NPP (Ecto-phosphodiesterase/Pyrophosphatase) Family and 198
Related Proteins
FIGURE 5-17 Identification of the Human tRNA Synthetase-Like Protein mRNA and the Human 209
Ribosomal S5 mRNA
FIGURE 6-1 Proposed Model for the Function of IRAK-1 in IL-iP-Induced Signal Transduction 219
To NFkB
Abbreviations
Abs Absorbance
AD Activation Domain
Amp Ampicillin
AP-1 Activating Protein 1
APC Antigen Presenting Cell
3-AT 3-Amino Triazole
ATP Adenosine Triphosphate
BD Binding Domain
BSA Bovine Serum Albumin
BCA Bincinchoninic Acid
CARD Caspase Recruitment Domain
Caspase Cysteine-dependent aspartate specific protease
cDNA Copy DNA
Ci Cubitus interruptus
CIAP Calf Intestinal Alkaline Phosphatase
cIAP Cellular Inhibitor of Apoptosis
CMV Cytomegalovirus
CNS Central Nervous System
Cos-2 Costal-2
CsCl Caesium Chloride
DD Death Domain
DEA Diethanolamine
DED Death Effector Domain
DISC Death-Inducing Signalling Complex
DNA Deoxyribonucleic Acid
DTT Dithiothreitol
ECL Enhanced Chemiluminescence
E. coli Escherichia coli
EDTA Ethylene Diamine Tetra-Acetic (Ethanoic) Acid
EGTA Ethylene Glycol-his (P-aminoethylether) N, N, N’, N’-Tetraacetic Acid
E-NPP Ecto-Nucleotide Pyrophosphatase/Phosphodiesterase
EtBr Ethidium Bromide
EV Elution Volume
FADD Fas-Associated Death Domain Protein
FCS Foetal Calf Serum
Fu Fused
p-Gal P-Galactosidase
GCG Genetics Computer Group
GCKR Germinal Centre Kinase-Related Protein
GRK G Protein Coupled Receptor Kinase
GRR Glycine Rich Region
GSK3P Glycogen Synthase Kinase 3(3
GTF General Transcription Factor
GTP Guanosine Triphosphate
HEK Human Embryonic Kidney
HEPES N-2-Hydroxyethy Ipiperazine-N ' -2-ethanesulphonic Acid
Hg Hedgehog
HLH Helix-Loop-Helix
HPLC High Performance Liquid Chromatography
HRP Horse Radish Peroxidase
ICE Interleukin-ip Converting Enzyme
IFN Interferon
Ig Immunoglobulin
IKK I Kappa B Kinase
IL-1 Interleukin-1
IL-18 Interleukin-18
IL-IR Interleukin-1 Receptor
IL-IRI Type I Interleukin-1 Receptor
IL-lRAcP Interleukin-1 Receptor Accesory Protein
IMAGE Integrated Molecular Analysis of Genomes and their Expression
IPTG Isopropyl P-D Thiogalactopyranoside
IR Intermediate Region
IRAK Interleukin-1 Receptor Associated Kinase
IRIP IRAKI-Interacting Protein
JNK c-Jun N-terminal Kinase
KCl Potassium Chloride
KD Kinase Domain
KDa Kilo Daltons
LAF Lymphocyte Activating Factor
LB Luria-Bertani Medium
LiAc Lithium Acetate
LPS Lipopolysaccharide
LRR Leucine Rich Repeat
LZ Leucine Zipper
MAPK Mitogen Activated Protein Kinase
MCE Mononuclear Cell Factor
MCS Multiple Cloning Site
MgClz Magnesium Chloride
M g S 0 4 Magnesium Sulphate
MHC Major Histocompatibility Complex
mRNA Message RNA
NaAc Sodium Acetate
NaCl Sodium Chloride
N a 2 H P 0 4 Sodium Hydrogen Phosphate
NaOH Sodium Hydroxide
NEMO NFkB Essential Modulator
NFkB Nuclear Factor Kappa B
NIK NFkB Inducing Kinase
NK Natural Killer
NLS Nuclear Localisation Signal
nm nanometer
NP-40 Nonidet P-40
PCR Polymerase Chain Reaction
PEG4000 Polyethylene Glycol 4000
PKA Protein Kinase A
PKA-Ca Type-a cAMP-dependent Protein Kinase Catalytic Subunit
PLAD Pre-ligand Binding Assembly Domain
PMSF Phenylmethylsulfonyl Fluoride
PMN Polymorphonuclear Leukocyte
p-NPP p-Nitrophenol Phosphate
PP Protein Phosphatase
PPi Pyrophosphatase
PRR Pattern Recognition Receptor
PVDF Poly vinylidene Difluoride
RAIDD RIP-Associated ICHl/CED3-Homologous Protein with a Death Domain
RHD Rel Homology Domain
RIP Receptor Interacting Protein
RNA Ribonucleic Acid
RNase Ribonulease
rpm Revolutions per minute
RSV Rous Sarcoma Virus
SAP Shrimp Alkaline Phosphatase
sAP Secreted Alkaline Phosphatase
SCF Skp 1 -Cullin-F-box
SD Synthetic Dropout
SDM Site-Directed Mutagenesis
SDS Sodium Dodecyl Sulphate
SEM Standard Error of the Mean
SIGGIR Single Ig IL-1 Receptor Relative
SODD Silencer of Death Domains
Su Suppressor of Fused
TACE TNFa Converting Enzyme
TAK-1 TGFp-Activated Kinase-1
TCA Trichloroacetic Acid
TD Transactivation Domain
TE Tris EDTA buffer
TNF Tumour Necrosis Factor
TNFRl Type 1 TNF Receptor
TRADD TNF-Receptor Associated Death Domain Protein
p-TRCP p-Transducin-Repeat-Containing Protein
TRIP TRAF-Interacting Protein
UAS Upstream Activating Sequences
UV Ultraviolet Radiation
18W 18-Wheeler
Wg Wingless
X-Gal 5-bromo-4-chloro-3-indolyl-P-D-galactopyranoside
The standard genetic code:
UUU Phe
UUC Phe
UUA Leu
UUG Leu
CUU Leu
c u e Leu
CUA Leu
CUG Leu
AUU He
AUG He
AUA He
AUG Met
GUU Val
GUC Val
GUA Val
GUG Val
AUG is part of the initiation signal, as well as being the codon for internal methionine
Ser UAU Tyr UGU Gys
Ser UAC Tyr UGG Gys
Ser UAA STOP UGA STOP
Ser UAG STOP UGG Trp
Pro CAU His GGU Arg
Pro CAC His GGG Arg
Pro CAA Gin GGA Arg
Pro GAG Gin GGG Arg
Thr AAU Asn AGU Ser
Thr AAG Asn AGG Ser
Thr AAA Lys AGA Arg
Thr AAG Lys AGG Arg
Ala GAU Asp GGU Gly
Ala GAG Asp GGG Gly
Ala GAA Glu GGA Gly
B Asx Asparagine or Aspartic acid
C Cys Cysteine
D Asp Aspartic acid
E Glu Glutamic acid
F Phe Phenylalanine
G Gly Glycine
H His Histidine
I He Isoleucine
L Leu Leucine
K Lys Lysine
M Met Methionine
N Asn Asparagine
P Pro Proline
Q Gin Glutamine
R Arg Arginine
S Ser Serine
T Thr Threonine
V Val Valine
W Trp Tryptophan
Y Tyr Tyrosine
Chapter 1
Introduction
Eukaryotic organisms have developed elaborate mechanisms to ensure that gene expression is tightly
regulated and specifically activated in response to particular developmental and extracellular stimuli. Gene
expression can be regulated in several ways, for example, post-transcripticmally through RNA splicing or
stability (Hodges and Bernstein, 1994; Ross, 1996), at the level of protein translation (Kaufinan et al,
1994) and also the stability of the mature protein can be controlled by processes such as the ubiquitin-
conjugation system (Jennissen, 1995). Sequence-specific DNA-binding proteins known as transcription
(trans-acting) factors modulate transcription initiation in Eukaryotes. These factors can repress or activate
transcription through inta-actions with transcription initiation sequences located within the promoter
regions of genes, or through interactions with regulatory (cis-regulatory) elements known as enhancers.
Many extracellular mediators such as growth factors and cytokines activate or repress the action of
transcription fectors, leading to changes in gene expression. Interleukin-1 (IL-1) is a key mediator in the
body’s response to microbial invasion, injury, or antigenic challenge. It activates a number of signalling
pathways, one of which culminates in the activation of the transcription factor NFkB. Dysregulation of
NFkB has been associated with cellular transformation and NFkB controls genes of significant biomedical
importance, such as those encoding proinflammatory cytokines, chemokines, interferons, major
histocompatibility complex (MHC) proteins, growth factors, cell adhesion molecules and viruses (Baeuerle
and Baltimore, 1996). Elucidatim of the IL-1 signal transduction pathway leading to NFkB activation
should therefore provide valuable insight into potential mechanisms to alleviate inflammation.
1.1
Overview of Innate and Adaptive Immunity
The immune system is an organisation of cells and molecules with specialised roles in defending against
infection. There are two fundamentally different types of responses to invading microbes, termed innate
and adaptive immunity (Delves and Roitt, 2000). Innate immune responses involve effector cells
(neutrophils, monocytes, macrophages) that can phagocytose and kill microbes, as well as cells tiiat release
inflammatory mediators (basophils, eosinophils, mast cells), which kill cells infected with intracellular
pathogens. The molecular components of innate responses include complement, acute-phase proteins, and
Adaptive immune responses involve the selection and proliferation of antigen-specific B and T cells, which
occurs when the surface receptors of these cells bind to antigen. Specialised cells, called antigen-presenting
cells (APCs), display the antigen to lymphocytes and collaborate with them in the response to the antigen
(Delves and Roitt, 2000). B cells secrete immunoglobulins (Igs), the antigen-specific antibodies
responsible for eliminating extracellular micro-organisms. T cells assist B cells to make antibodies and can
also eradicate intracellular pathogens by activating macrophages and by killing virally infected cells
(Delves and Roitt, 2000). The main distinction between the innate and adaptive arms of the immune system
lies in the mechanisms and receptors used for immune recognition. In the adaptive immune system,
diversity in the genes encoding T-cell receptor and B-cell receptor structures is generated somatically,
during the development of B and T cells (Medzhitov and Janeway, 2000). Each lymphocyte displays a
structurally unique receptor, so the repertoire of antigen receptors in the lymphocyte population is very
large and extremely diverse. This increases the probability that an individual lymphocyte will encounter an
antigen that binds to its receptor thereby triggering activation and proliferation of the cell (Medzhitov and
Janeway, 2000). However, since the binding sites of these antigen receptors arise as a result of random
genetic mechanisms, the receptor repertoire contains binding sites that can react not only with infectious
micro-organisms, but also with self-antigens.
Innate immune recognition is mediated by germ-line-encoded receptors that have broad specificity, because
they can recognise ligands which share a common molecular pattern. These receptors are therefore known
as pattem-recognition receptors (PRRs). The best-known examples of pathogen-associated molecular
patterns are highly conserved molecular structures, such as bacterial lipopolysaccharide (LPS),
peptidoglycan, lipoteichoic acids, mannans, bacterial DNA, double-stranded RNA and glucans (Medzhitov
and Janeway, 2000). Typically, PRRs are expressed on cells that are the first to encounter pathogens during
infection, such as neutrophils, macrophages and dendritic cells. PRRs are also found on B cells - the
professional antigen-presenting cells. Once pathogens are recognised by the PRRs, the innate immune cells
can perform their effector functions rapidly, as selection and proliferation is not necessary. This explains
the rapid kinetics of iimate immune responses, which are essential to limit the expansion of rapidly dividing
microorganisms (Medzhitov and Jane way, 1997a). As pathogen-associated molecular patterns are
produced only by microbial pathogens, the innate immune system can discriminate between “foreign” and
The recognition of “non-self’ may provide important signals to direct subsequent adaptive immune
responses.
Functionally, PRRs can be divided into three classes of molecules that are involved with secretion,
endocytosis or signalling. The best-characterised secreted receptor is the mannan-binding lectin, which
binds to carbohydrates on Gram positive and Gram negative bacteria, yeast and some viruses and parasites
(Medzhitov and Janeway, 2000). This initiates the lectin pathway of complement activation (Epstein et al,
1996; Fraser et a l, 1998). Endocytic PRRs occur on the surface of phagocytes and mediate the uptake and delivery of the pathogen into lysosomes, where it is destroyed and processed. The resulting peptide can be
presented by MHC molecules on the surface of the macrophage (Medzhitov and Janeway, 2000). An
example of such a PRR receptor is the macrophage mannose receptor (Fraser et a l, 1998). The signalling receptors, once they have recognised pathogen-associated molecular patterns, activate signal transduction
pathways that induce the expression of a variety of immune and inflammatory response genes.
Recently identified Toll family members appear to have a major role in the induction of immune and
inflammatory responses (Ghosh et al, 1998). The prototype Toll-receptor was discovered in Drosophila,
where signalling via the Toll- and related 18 wheeler (18W) receptors, mediate the production of anti-fungal
and anti-bacterial peptides respectively (Lemaitre et al, 1996; Williams et a l, 1997). In this respect, the
Drosophila receptor appeared to function as a PRR. Interestingly, evidence that the promoter regions of the
genes encoding these peptides contain NFkB binding sites and that the cytoplasmic region of the
Drosophila Toll protein was shown to exhibit sequence similarity to the cytoplasmic domain of the mammalian type I interleukin-1 receptor (IL-IRI) (Gay and Keith, 1991) further supported a role for Toll in
innate immunity. Both receptors induce signalling pathways that lead to the activation of transcription
factors of the NFkB family (Ghosh et a l, 1998). Subsequently, a human homologue of the Drosophila Toll receptor was cloned in 1997, called Toll-like receptor 4 (TLR4) (Medzhitov et a l, 1997) and to date 10 human TLRs have been reported or annotated in databases. TLR4 overexpression was shown to induce the
activation of NFkB, leading to the expression of a variety of cytokines and costimulatory molecules that are
TLR2, on the other hand, is required for responses to Gram positive bacteria and fimgi, responding to
products such as peptidoglycan and lipoteichoic acid (Brightbill et a l, 1999; Underhill et a l, 1999). These findings suggest that TLRs may fimction as PRR receptors of the innate immune system and play a role in
controlling adaptive immune responses.
As mentioned, the adaptive immune system responds to a pathogen only after it has been recognised by the
innate immune system, preventing the induction of immune responses to self-antigens. For example, T
cells use their antigen receptors to recognise a ligand in the form of a peptide bound to an MHC class II
molecule on the surface of an AFC (Medzhitov and Janeway, 2000). However, these peptides can be either
self peptides or peptides derived fi’om a microbial pathogen. The recognition of the peptide-MHC ligand by
the antigm receptor alone is not sufficient to activate T cells. Signals generated via PRRs on activated
innate immune cells also promote the expression of co-stimulatory molecules, such as CD80 and CD86 on
the surface of the APC (Medzhitov and Janeway, 2000). It is only when the APC expresses both antigen
and CD80 or CD86 molecules that the T cell can be activated. As this is controlled by pathogen
recognition, this is an important primary mechanism to ensure that T-cell responses are targeted only
towards APC’s recognising “non-self’ structures and may also serve to modulate the ensuing adaptive
response (Fearon and Locksley, 1996). These properties represent a previously unrecognised role for cells
of the innate system in primary recognition, education and integration of appropriate immune responses.
Innate responses fi'equoitly involve cytokines, such as IL-1, TNF and IL-6. Cytokines are soluble, low
molecular weight proteins that act as messengers both within the immune system and between the immune
system and oÜict systems of the body, forming an integrated network that is highly involved in the
regulation of immune responses (Delves and Roitt, 2000). Cytokine efiects are pleiotropic and influaiced
by dose as well as presence or absence of othCT cytokines. They are produced primarily by activated innate
effector cells, such as monocytes/macrophages, natural killer (NK) cells, polymorphonuclear leukocytes
(PMN) at sites of tissue infection or damage and can rapidly initiate a local acute inflammatory reaction
causing the recruitment and activation of additicmal inflammatory cells (Delves and Roitt, 2000). Under
normal conditions, these cells contribute to an acute self-limiting inflammatory response, which effectively
eliminates the infection and stimulates tissue repair to bring about resolution of injury and restoration of
Activated APCs that have reached local lymphatic structures can also produce cytokines, which regulate T
and B- cell responses to initiate longer-term adaptive immunity.
Certain defense mechanisms appear to be prominent during viral infections. Foremost amongst them is the
early production of IFNa and IFNP (Biron, 1998). IFNs, among other effects, strongly upregulate the
expression of the MHC class I molecules, thus increasing the efiSciency of presentation of viral peptides to
cytotoxic T cells (Sen and Lengyel, 1992). IL-1, TNFa and chemokines, direct the migration of antigai-
specific lymphocytes, along with other effector cells, to the site of infection either by inducing the
expression of adhesion molecules on endothelial cells (IL-1 and TNFa) or by stimulating chemotaxis
(chemokines) (Murphy, 1994; Shimizu et al, 1992). Finally, IL-6 induces the terminal differentiation of B lymphocytes into Ig-producing plasma cells (Medzhitov and Janeway, 1997b; Van Snick, 1990).
Under pathological conditions persistent activation of inflammatory cells by an irritant or foreign organism
may cause chronic overproduction of inflammatory cytokines and excessive activation of effector
mechanisms which lead to tissue damage. In these circumstances a h^eakdown or short-circuiting of nœmal
innate mechanisms of recognition may also lead to inappropriate presentation of self-antigens leading to
activation of auto-reactive T-cells and auto-immune disease. Monocytes and macrophages are important
effector cells which, when activated, can produce large amounts of potent proinflammatory cytokines such
as lL-1 and TNF. Recently it has been recognised that in addition to their important roles in host
protection, over-production of these cytokines can be contributory fectors in chronic inflammatory diseases
such as rheumatoid arthritis (RA), inflammatory bowel disease (IBD) and asthma (Dinarello, 1996).
Therapeutic strategies based on antagonists of these cytokines have shown clinical benefits in several
diseases, particularly RA and IBD confirming the importance and relevance of these cytokines in
1.2
Interleukin-1 (IL-1)
1.2.1 Biological Effects O f IL-1
Interleukin-1 is the name given to two genetically distinct polypeptides (IL-1 a and IL-ip) that signal
through the same cell surface receptor and share many biological properties. They were originally known
by a number of different names (endogenous pyrogen, mononuclear cell factor (MCF), lymphocyte
activating factor (LAF), catabolin etc) corresponding to their diverse biological activities (Stylianou and
Saklatvala, 1998). Much attention has focused on IL-1 because of its critical role in immune and
inflammatory responses. It is one of the key mediators in the body’s response to microbial invasion,
inflammation, immunological reactions and injury. In the first few hours after infection or injury, the
biological effects of IL-1 are manifested in nearly every tissue and OTgan (O'Neill, 2000). On activation by
IL-1, cells display an inflamed phenotype, caused by the expression of over 90 genes, including those
coding for other cytokines, cytokine receptors, acute-phase reactants, growth factors, tissue remodelling
enzymes, extracellular matrix components and adhesion molecules (O'Neill, 1995). Most of these genes are
NFicB-regulated, so this transcription factor has become the focus of extensive research due to its virtual
ubiquity as a regulator of inflammatory and immune gene expression. Much of the work presented in this
thesis has focused on elucidating the mechanisms by which IL-1 signals via the IL-IRI to activate NFkB.
IL-1 is primarily produced by activated monocytes and macrophages, although almost all cells, including
fibroblasts, endothelial cells, kératinocytes, osteoblasts and neutrophils, can produce IL-1 to some extent
(Bresnihan and Cunnane, 1998; Rosenwasser, 1998). While the constitutive expression of IL-1 is very low
in health, it is highly inducible in response to a variety of stimuli, including endotoxins, micro-organisms,
various cytokines, fragments of complement, immune complexes and products of activated T-lymphocytes
(Dinarello, 1992). It is able to act on most tissues and can mediate many physiological, metabolic,
inflammation, immunological and haematopoietic responses. Table 1-1 gives examples of the plethora of
Immunological Properties
T cell activation: synergy with IL-6 for IL-2 synthesis
Increased lymphokine synthesis (IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-10 and IL-12)
Increased IL-2RP
B cell activation via induction of IL-6: increased antibody production
Natural killer activity: synergy with IL-2 and IFN
Inhibition of tolerance to protein antigens
Pro-inflammatory Properties
Fever, sleep, anorexia, neuropeptide
Amino acid turnover, hyperlipidemia
Hypotension, myocardial suppression, shock, death
Cytotoxicity (apoptosis of insulin-producing islet P cells)
Endothelial cell activation
Increased adhesion molecule expression (ICAM-1, VCAM-1 and ELAM-1)
Neutrophilia
Neutrophil priming
Neutrophil tissue infiltration via induction of IL-8
Eosinophil degradation
Cyclooxygenase and lipooxygenase gene expression
Synthesis of collagenases
Cartilage breakdown
Osteoblast activation
Induction of osteoclastogenesis and bone resorption
T ab le 1-1: T h e B io lo g ica l E ffects o f IL-1. Figure adapted from (D inarello, 1992) and (Dinarello, 1996).
Abbreviations: lC A M -1, intracellular adhesion m o lec u le-1; V C A M -1, vascular cell adhesion m o lec u le-1; ELAM -1,
1.2.2 Molecular Nature o f IL -1
As mentioned above, the biological activity of IL-1 is mediated by the two agonist forms IL-1 a and IL-ip
(Auron et a l, 1984; March et a l, 1985), which show only 25% primary amino acid sequence identity, yet bind to the same cell surface receptor, the IL-IRI and initiate similar biological responses (Dower et a l,
1985; Dower et a l, 1986; McMahan et a l, 1991; Sims et a l, 1988), These cytokines are synthesised as
31kDa precursors (pro-IL-la and pro-IL-ip) without hydrophobic leader sequences and are proteolytically
cleaved into mature ITkDa proteins (Cameron et a l, 1986; Kostura et a l, 1989). Unlike IL-ip, IL-l a is
fully active as a precursor protein and remains intracellularly (Mosley et a l, 1987). IL -la is not commonly found in the circulation or in body fluids, except during severe disease, in which case it is released from
dying cells. In the absence of cell death, pro-IL-la is cleaved by calcium-dependent membrane-associated
cysteine proteases called calpains, allowing mature IL-1 a to be secreted from cells (Kobayashi et a l, 1990), (Miller et a l, 1994; Watanabe and Kobayashi, 1994). The cysteine protease responsible for the processing
of pro-lL-ip from its inactive precursor to the bioactive form is called the interleukin-ip converting
enzyme (ICE) / caspase-1 (Black et a l, 1988; Black et a l, 1989a; Cerretti et a l, 1992; Kostura et a l, 1989; Thomberry et a l, 1992). Caspase-1 cleaves pro-IL-ip at Asp 116-Ala 117, creating the 17.5kDa mature biologically active cytokine, wiiich is exported from the cell via a non-classical secretory pathway (Auron
and Webb, 1987; Black et a l, 1989b; Hazuda et a l, 1990; Howard et a l, 1991; Thomberry et a l, 1992).
Although caspase-1 is unable to cleave pro-IL-la, it can cleave an inactive precursor of interferon-y-
inducing fector / IL-18, the third member of the IL-1 family of cytokines (Ghayur et a l, 1997; Gu et a l,
1997; Howard et a l, 1991; Okamura et a l, 1995; Ushio et a l, 1996). Caspase-1 is therefore involved in the regulation of IL-18 production and the subsequent activation of T cells. Human caspase-1 is
synthesised in cells of the monocytic lineage as an inactive 45kDa precursor, wiiich is processed
proteolytically to generate enzymatically active 10- and 20kDa proteins (Miller et a l, 1993; Thomberry et a l, 1992).
The fourth member of the IL-1 femily is the naturally occurring IL-1 receptor antagonist (IL-lRa). IL-lRa
is also synthesised as a precursor protein, pro-lL-lRa, that is translated in the Endoplasmic Reticulum (ER)
In contrast to IL-1 a and IL-1 (3, pro-IL-lRa evolved with a 25 amino-acid leader sequence and once
cleaved, is readily transported out of cells and termed secreted IL-lRa (sIL-lRa) (Dinarello, 1994). An
intracellular form of the IL-lRa, (icIL-lRa) which is retained inside cells due to the absence of a leader
peptide has also been identified (Haskill et a l, 1991). icIL-lRa is synthesised by alternate mRNA splice/insertion of the IL-lRa transcript, replacing the first exon coding for the signal peptide (Dinarello,
1996; Haskill et a l, 1991). Studies using human blood monocytes have shown that the gene for sIL-lRa is initially expressed following cell stimulation and can be visualised in the Golgi during the first 4-6 hours
(Andersson et a l, 1992). However, after 24 hours, icIL-lRa is the primary transcript detected in the cytoplasm of these cells (Andersson et a l, 1992). Both sIL-lRa and icIL-lRa are fimctionally indistinguishable. sIL-lra blocks cell surface IL-1 receptors, while at least in kératinocytes and epithelial
cells, the constitutively produced icIL-lRa is thought to inhibit intracellular binding of IL-1 a to nuclear
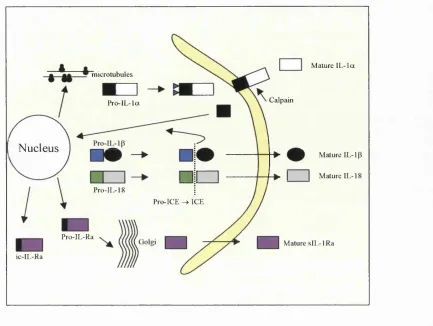
receptors (Dinarello, 1994; Haskill et al, 1991). See figure 1-1 for a diagrammatic representation of the production of active IL-1 a, IL-ip, IL-lRa and IL-18.
Analysis of the human genome sequence and other DNA databases has led to the discovery of six gene
sequences predicted to encode homologues of IL-1, called IL-1F5-IL-1F10 (Barton et al, 2000; Busfield et al, 2000; Debets et al, 2001; Kumar et al, 2000; Lin et al, 2001; Mulero et al, 1999; Pan et al, 2001; Smith et a l, 2000,). AU members of the IL-1 family, with the exception of IL-18, which maps to chromosome llq , are clustered in the same region of human chromosome 2q, consistent with the notion of
evolution by gene duplication (Dinarello, 1996; Kumar et a l, 2000; Smith et a l, 2000). The discovery of these novel members of the IL-1 family is interesting due to the identification of orphan receptors belonging
to the IL-IR superfamily (O’Neill and Dinarello, 2000). Possible roles for some of the novel IL-1
homologues in immunity and inflammation are beginning to emerge. For example, IL-1F5 and IL-1F9 are
expressed strongly in kératinocytes and the expression of IL-1F9 is induced in skin during contact
hypersensitivity and in psoriasis patients (Dunn et al, 2001). IL-1F7 and IL-IF 10 have been shown to bind to the IL-18R and the soluble IL-IRI respectively, although specific ftmctions for these proteins, as well as
microtubules
Calpam P ro-lL -la
Nucleus —
Pro-IL-18
Pro-ICE ^ ICE
Pro-IL-Ra
ic - I I,- R a
Mature IL -Ia
Mature IL-1 p I I Mature IL-18
Mature sIL-lR a
Figure 1-1: H um an Blood M onocyte P rod ucin g I L - I a , IL -ip , IL -18 and IL rlR a. Figure adapted from (Dinarello,
1996; Dinarello, 1998). P ro-IL -la is translated in association with microtubules and remains cytoplasm ic, where it is
myristoylated. This allow s translocation o f p ro-IL -la to the cell membrane where it is cleaved by cysteine proteases
known as Calpains (Dinarello, 1998). The mature 17kDa IL -la is released from cells, whereas p ro-IL -la and the pro
piece (amino acids l - 1 15 o f IL -Ia ) can bind to nuclear D N A (Dinarello, 1998; W essendorf et al., 1993). The IL ip
-converting enzym e (ICE) is translated in the Endoplasmic Reticulum as an inactive precursor (pro-ICE) and requires
tw o internal cleavage steps to produce an active ICE heterodimer (M iller et ai, 1993; Thomberry et a i, 1992).
Pro-IL-ip and pro-IL-I8 are cleaved by ICE and the active mature IL -Pro-IL-ip and IL-18 m olecules are released into the
extracellular compartment (Gu et a i, 1997; Thomberry et a i, 1992). The pro-pieces o f IL -ip and IL-18 are found both
intra- and extracellularly, but the precursor m olecules are predominantly found in the cytosol and are inactive
(Dinarello, 1998). IL -lR a is synthesised as a precursor, pro- IL-IRa that is translated in the Endoplasmic Reticulum
and transported to the Golgi (Eisenberg et ai, 1990). pro-IL-lRa evolved with a 25 amino-acid leader sequence and
once cleaved, is readily transported out o f cells and termed secreted IL-IRa (sIL-IR a) (Dinarello, 1994). An
intracellular form o f the IL-IRa. (icIL-IR a) also exists, that is retained inside cells due to the absence o f a leader
1.3
The IL-1 Receptor/TolI-like Receptor Superfamily.
Two different IL-1 receptors, types I and II (IL-IRI and IL-IRII) have been cloned and characterised (Sims
and Dower, 1994). These receptors are single transmembrane proteins that possess 3 conserved
extracellular Ig-like domains of 319 amino acids responsible for ligand binding (McMahan et a l, 1991; Sims et a l, 1988). The IL-IRI has a long cytoplasmic region of 217 ammo acids which allows it to transduce cellular signalling events, while IL-IRII, containing less than 30 cytoplasmic amino acids, acts as
a decoy receptor, competing with IL-1 (3 for binding to the IL-IRI (Colotta et a l, 1993; Stylianou et a l,
1992). In 1995, it was shown by Greenfeder et al, that an additional subunit of the IL-IRI was necessary for IL-1 signal transduction, called the IL-1 receptor accessory protein (IL-lRAcP) (Greenfeder et a l,
1995). This accessory protein is capable of cross-linking to IL-1 and increases the afiBnity of the IL-IRI for
the ligand by about 5-fold (Greenfeder et a l, 1995; Korherr et al., 1997; Wesche et a l, 1997a). Furthermore, studies using gene knockout mice have revealed that both the IL-IRI and the accessory
protein are essential for IL-1-dependent NFkB activation (Labow et a l, 1997; Wesche et a l, 1997a). Moreover, structural studies have identified differences between agonist/antagonist-bound conformations of
the IL-lRl, which may relate to interactions of the IL-lRAcP with the receptor complex (Auron, 1998;
Schreuder et a l, 1997; Vigers et a l, 1997). These studies have shown that the ligands IL -la, IL-lp and the
IL-IRa are all made up of 12 (3-strands, which form an open P-barrel, that is capped at one end by a P-sheet.
IL-ip associates with the extracellular domain of the IL-IRI via two distinct interactions: the open end of
the barrel interacts with domain III of the receptor, while domains I and II act co-operatively to bind the
back side of the IL-ip through distinct residues. This results in IL-1 P being buried deep into the receptor,
with a total buried ligand surfece of 3.089Â. In contrast, the IL-IRa binds with high affinity to domains I
and II of the IL-IRI, but shows minimal association with domain HI. Only 1.744Â of the surface of IL-IRa
is buried in the receptor. It can be proposed therefore, that the binding site missing iBrom the IL-IRa, is in
feet, the site that binds the IL-lRAcP, although no coxtal structure has yet emerged. In addition to the IL-
These proteins are classified according to the presence of extracellular Ig-like motife and an
intracytoplasmic TIR (Toll/IL-1 receptor) homology domain. Among these are the IL-18 receptor (1L-18R)
(Hoshino et al., 1999; Torigoe et a i, 1997) and it’s accessory protein, AcPL (Bom et al., 1998), IL-lRrp2 (Lovenberg et al., 1996), T1/ST2 (Mitcham et al., 1996) and the recently discovered IL-IRAPL (IL-1 receptor accessory protein-like) (Carrie et al., 1999) (see Figure 1-2, subgroup 1), The IL-18R and it’s accessory protein form a complex to initiate signal transduction and appear to signal in a similar fashion to
IL-1 (Bom et a l, 1998; Kojima et a l, 1998; Matsumoto et a l, 1997). Like lL-1, IL-18 triggers phosphorylation of IRAK-1 and the recruitment of IRAK-1 to the IL-18 receptor complex (Kojima et a l,
1998; Robinson et a l, 1997). However, IL-18 and IL-1 act on different cell types and lead to divergent
cellular responses. IL-18 has been implicated primarily in inducing IFNy production fi’om NK and T-helper
(Thl) cells (Adachi et a l, 1998; Takeda et a l, 1998), whereas IL-1 is a potent inducer of IL-6 fi"om fibroblasts and macrophages during inflammation (Akira and Kishimoto, 2000; Bankers-Fulbright et a l,
1996; Kanakaraj et a l, 1998). The T1/ST2 receptor is expressed on macrophages and Th2 cells, but its precise role remains unclear (Mitcham et a l, 1996). IL-lRrp2 is more closely related to the IL-IRI than any other proteins, including the IL-IRII and the IL-lRAcP, yet it is does not bind, at least by itself, to IL-
la , IL-ip or the IL-lRa (Lovenberg et a l, 1996). IL-IRAPL is a novel receptor found on the X- chromosome, which is associated with mental retardation. Its discovery has highlighted the importance of
IL-1 signalling pathways in cognitive fimction and in the normal physiology of the CNS. The ligands for
these three novel receptors remain to be identified.
The second group witiiin the IL-IR/Toll receptor super family, consists of type I membrane proteins that
contain multiple leucine rich regions (LRR) extracellularly (see Figure 1-2, subgroup 2). Toll is the
prototypic member of this group, but there are now eight other receptors identified in the fly, including
18W (Eldon et a l, 1994), MstProx (Mitcham et a l, 1996) and Tehao (Luo and Zheng, 2000) (Figure 1-2, subgroup 2). In 1991, Nick Gay and Fionna Keith made the striking observation based on amino acid
sequence comparisons, that the intracellular domain of the IL-lRl contained a 200 amino acid region
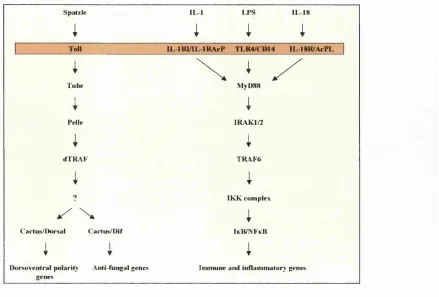
F ig u re 1-2: T h e C u rr en tly K n o w n M em b er s o f th e I L -I R /T L R S u p erfa m ily . A ll members contain a conserved
cy to so lic region termed the TIR dom ain (T oll-IL -IR dom ain). T he superfamily can be divided into tw o subgroups. The
members o f subgroup 1, the im m unoglobulin (Ig) subgroup, all contain extracellular Ig dom ains and include receptors
and accessory proteins for IL-1, IL-18, and the orphan receptor T I/S T 2. Subgroup 2, the leucine-rich repeat (LRR)
subgroup, includes the signalling receptors for LPS (TLR-4) and m olecules from G ram -positive bacteria such as
peptidoglycan and lipoproteins (TLR-2). Several plant members are also in this subgroup. MyDSB is ex clu siv ely
cy to so lic and is a signalling adapter for IL -IR I, IL-18R , TLR-2, and TLR-4, i f not the entire fam ily. Superfam ily
members m ay also occur in Caenorhabditis elegans and Streptomyces coelicor (not show n). (B ) Three w ell-conserved
regions in the TIR dom ain, as derived from an alignm ent o f 31 fam ily members (O ’N eill and Dinarello, 20 0 0 ) S in gle
letter abbreviations for amino acid residues are as follow s: A , Ala; C, Cys; D , Asp; F, Phe; G, Gly; I, He; K, Lys; L,
Leu; P, Pro; R , Arg; S, Ser; W , Trp; and Y , Tyr. The entire dom ain spans about 2 0 0 amino acids with varying numbers
o f am ino acids separating the b oxes, depending on the particular receptor. There are additional regions within the
dom ain that exhibit high similarity. Subconsensus sequences for IL -IR , TLR, and plant members have also been
calculated (R ock et a l, 1998), but the three boxes show n are the m ost highly conserved regions. A Pro to His mutation
in box 2 o f the g en e encoding TLR -4 renders TLR -4 unable to signal in C 3H /HeJ m ice (Poltorak et a l, 1998). Studies
usin g site-directed m utagenesis indicate that A rg and A sp in b ox 2, and Phe and Trp in b ox 3, are essential for IL -IR I
function (H eguy et a l, 1992). Arg'*^’ in IL-IR I, w hich lies outside the three conserved b oxes, is also required for
signallin g (H eguy et a l, 1992). Figure and legend taken from review b y Luke O ’N eill with perm ission (O 'N eill and
T h e IL -IR /T L R superfam ily
(A) Superfamily members
Sub-group 1
Sub-group 2 Name IL-1RI IL-IRII B15R IL-IRAcP IL-18R IL-ISRAcP T1/ST2 IL-1Rrp2 JL-1RAPL SIGGIR MyD88 "dToll cIToll trToll 18W MstProx Tehao TLR1 TLR2 TLR3 TLR4 TLR5 TLR6 N L6 RPP1 RPP5
I TIR
Domains
LRR
(B) The TIR domain
o o ca
O O O C Z ]
O t U H C Z l
D-K-YDAF-SYS
Box 1
G Y K L C I-R D -P G
Box 2 Present in Mammals Mammals Vaccinia Mammals Mammals Mammals Mammals Mammals Mammals Mammals Mammals Insects Insects Insects Insects Insects Insects Mammals Mammals Mammals Mammals Mammals Mammals Plants Plants Plants Plants FWKK
As mentioned earlier, this conserved moti^ which has been termed the TIR (Toll/IL-1 Receptor) domain,
has subsequently been used to define members of the IL-1 receptor / Toll receptor super family (O'Neill and
Dinarello, 2000). It appears that Drosophila is capable of discriminating between classes of invading microorganisms. Toll receptors play a central role in innate immunity in the adult fly, controlling
expression of antifimgal gmes through NFkB sites in their promoters and also in the establishment of
dorsoventral polarity during embryogenesis (Hashimoto et a l, 1988; Lemaitre et a l, 1996; Levashina et a l,
1998). Fly embryos lacking the Toll-related receptor 18W, show increased lethality to bacterial challenge
and hence have confirmed that 18W also plays a role in eliciting the innate immune response in Drosophila
(Eldon et a l, 1994; Williams et a l, 1997). 18W signalling is responsible for mounting an immune defence against bacterial gene products such as attacin, through the activation of Dif, whereas Toll appears to
function in antifimgal responses by activation of Dorsal (Lemaitre et a l, 1996; Williams et a l, 1997). The roles of MstProx and Tehao however remain to be established. Four plant proteins involved in disease
resistance have also been identified. These are the tobacco N gene, the flax L6 gene and RPPl and RPP5
found in Arabidopsis (Lawrence et a l, 1995; Meyers et a l, 1999; Parker et a l, 1997; Whitham et a l,
1994). Like the Toll receptors, these receptors contain a TIR domain preceded by nucleotide binding and
LRR segments. However, in contrast to the Toll receptors, these proteins are intracellular. There ligands
are wholly unknown.
As in insects, a key feature of innate immunity in mammals is the ability to limit infectious challenge
rapidly (Hoffinann et a l, 1999). Over the last few years, a family of mammalian Toll-like receptors (TLRs) has been identified (Rock et a l, 1998). These receptors are classified as Toll-like receptors because they possess an extracellular leucine-rich repeat (LRR) region (Medzhitov et a l, 1997; Rock et a l, 1998). To date, at least nine human cDNAs for Toll receptors have been reported or sequences annotated in public
databases (Chaudhary et a l, 1998; Mitcham et a l, 1996; Rock et a l, 1998; Takeuchi et a l, 1999). Not all of these TLR’s have been characterised functionally, but it is believed that at least TLR4 functions in
As can be seen in Figure 1-2, there are two members of the IL-IR/Toll R superfamily that do not contain
either IgG-like domains or LRRs. One is the cytosolic adapter molecule MyD88, which contains an N-
terminal death domain (DD), followed by the TIR domain and the other is a recently identified protein
termed SIGGIR (single Ig IL-1 receptor relative) (Thomassen et a l, 1999). Gene knockout studies have shown that MyD88 is essential not only for IL-lRI-mediated signal transduction, but also for endotoxin
signalling via the TLR4 receptor and in signalling mediated by the IL-18R (Adachi et a l, 1998; Kawai et a l, 1999). The ligand for the SIGGIR receptor and its function remain to be determined.
1.4
IL-lRI/TollR-mediated Signalling Pathways
It has been estimated that on many cell types, there are fewer than one hundred IL-1 receptors, of which less
than 5% need to be occupied to elicit a biological response. Consequently, IL-lR-mediated signal
transduction must be highly efficient and receptor signals greatly amplified. Rapid amplification of signals
is achieved through the activation of kinase cascades. IL-1 activates four signalling cascades, three of
which involve the mitogen activated protein kinases (MARK) p42/p44, c-Jun N-terminal kinase (INK) and
p38 and the other is a well characterised pathway which culminates in the activation of the transcription
factor NFkB (O'Neill, 1996). Since the discovery of the IL-IRI, many studies have focused on
understanding the mechanisms involved in receptor signal transduction leading to activation of these kinase
cascades in an attempt to explain the efficient coupling of the IL-IRI to downstream signalling events. Due
to the central importance of NFkB in the regulation of many inflammatory genes, there is considerable
interest in elucidating the mechanisms involved in activation of this transcription factor.
The intracellular portion of the IL-IRI receptor contains a domain that is homologous to the Drosophila
Toll receptor, which has been termed the TIR domain (Gay and Keith, 1991; Schneider et a l, 1991). Heguy et al, (1992) showed by site-directed mutagenesis, that residues within this region that are essential for IL-lR-mediated signal transduction, are conserved in the Toll protein, suggesting that both receptors
may share a common mechanism of action (Heguy et al, 1992). These studies also demonstrated that non conserved amino acid sequences could be mutated without affecting IL-1 signalling function (Heguy et al,
1992). The intracellular domains of the IL-IR/Toll receptors, although large enough, do not contain motifs
predicted to have intrinsic enzymatic activity, which might couple the receptor complexes to
It therefore seemed likely that novel, but similar signal transduction processes were involved in both IL-IR
and Toll-mediated signalling. This has turned out to be the case. Molecular characterisation of genes
involved in the Drosophila Toll signal transduction pathway revealed that striking parallels existed between
the cytokine-induced activatiœi of NFkB in mammals and the activation of the morphogen Dorsal in the fly
(see below). Taken together these observations suggested that the Toll signal transduction cassette pathway
had been conserved throughout evolution and that a homologous system was involved in the IL-IR-
mediated pathway in mammalian cells. Subsequently, this paradigm has been further strengthened by the
discovery of mammalian Toll-like receptors and additional signalling components. The Drosophila system has continued to provide a general insight into the regulation of Rel proteins in mammals.
1.4.1 Clues from the
Drosophila
Toll Pathway
During early embryogenesis in the fruit fly, activation of the Dorsal protein leads to a nuclear gradiait of
this morphogen, resulting in activation and repression of gene transcription (Roth et a l, 1989; Rushlow et a l, 1989; Steward, 1989). This gradient is formed due to the selective nuclear transport of dorsal to the ventral, but not the dorsal nuclei within the developing syncitium, upon binding of the ligand Spatzle to the
Toll receptor (Morisato and Anderson, 1994; Morisato and Anderson, 1995; Roth et al. 1989; Rushlow et al, 1989; Schneider et al, 1994; Steward, 1989). Toll activation correlates with the dissociation of cactus, an inhibitor protein that retains Dorsal as an inactive complex in the cytoplasm of cells (Belvin et a l, 1995; Geisler et a l, 1992; Roth et al., 1991). Recessive mutations of cactus result in a phenotype in which dorsal cell fetes are shifted ventrally (Govind and Steward, 1991). Molecular cloning of Dorsal and Cactus
revealed that these proteins were structurally, as well as functionally homologous to N FkB and IkB (Karin and Ben-Neriah, 2000). Genetic studies in Drosophila indicated tiiat products of the Tube and Pelle genes functioned downstream of the Toll receptor, but upstream of Dorsal and either parallel to, or upstream of
Cactus (Hecht and Anderson, 1993; Letsou et a l, 1991; Shelton and Wasserman, 1993). Tube and Pelle were thffefore likely candidate signal transducers for transmitting the signal required for the nuclear import
of the Dorsal protein (Hecht and Anderson, 1993; Letsou et a l, 1991; Shelton and Wasserman, 1993). Cloning and characterisation of Tube/Pelle was achieved before their functional counterparts in the