Full-length amelogenin influences the differentiation of human dental pulp stem cells
Full text
Figure
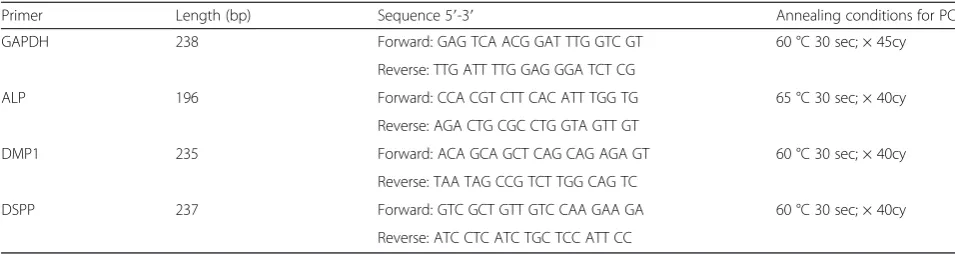
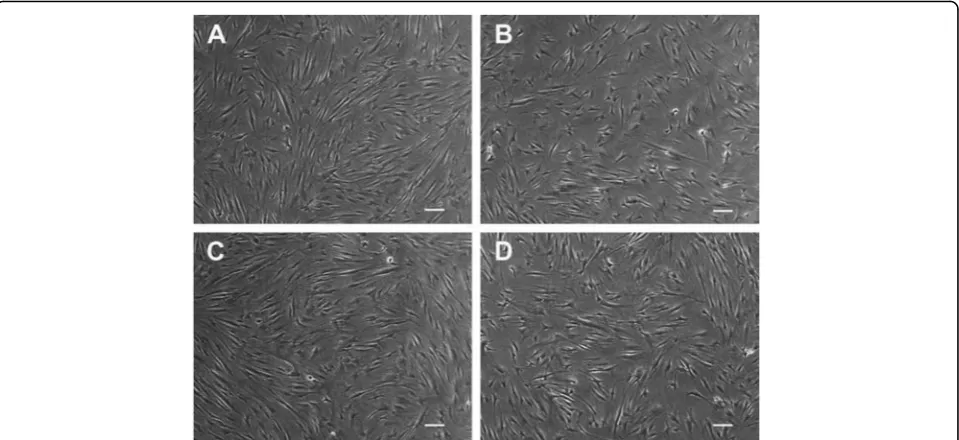
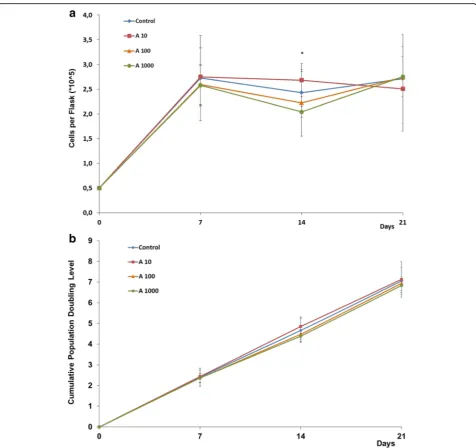
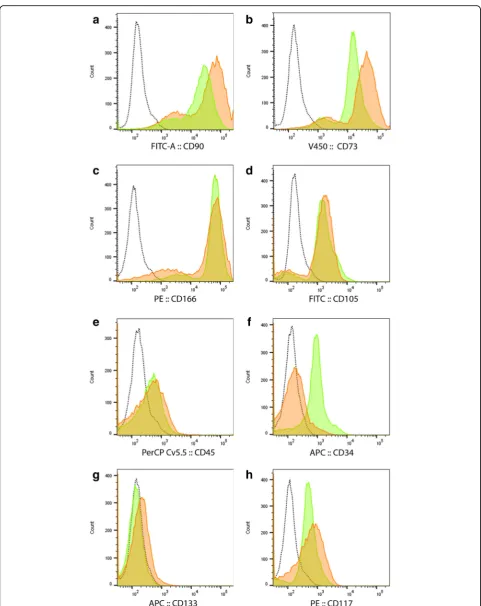
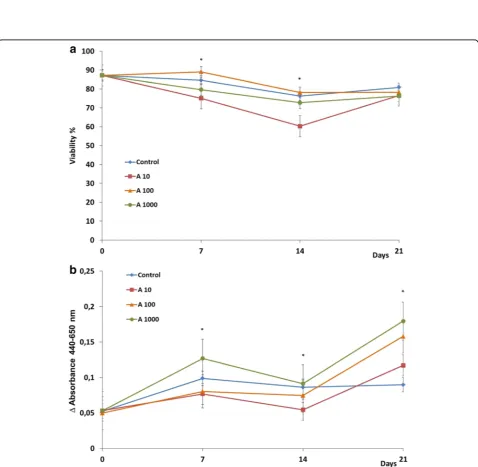

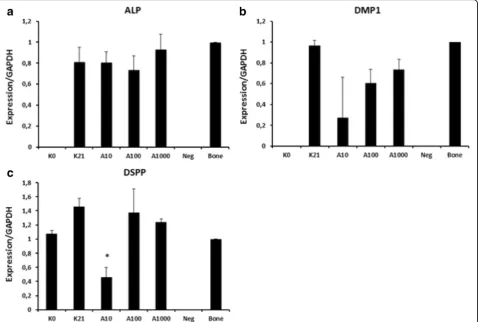
Related documents
hBM-MSCs: Human bone marrow-derived mesenchymal stem cells; hBM- MSC-Ex: Human bone marrow mesenchymal stem cell-derived exosomes; ALP: Alkaline phosphatase; ALT:
ALP = alkaline phosphatase; BMMSC = bone marrow mesenchymal stem cells; BrdU = bromodeoxyuridine; C3 = complement 3; CFU = colony forming units- fibroblastic; CTX =
ALP: alkaline phosphatase; AsA2-P: L-ascorbic acid 2-phosphate; β -GP: beta- glycerophosphate; BMP: bone morphogenetic protein; BMSC: bone marrow- derived mesenchymal stem cell;
ALP: Alkaline phosphatase; bFGF: Basic fibroblast growth factor; BMP2: Bone morphogenetic protein 2; BMSCs: Bone mesenchymal stem cells; BV/ TV: Trabecular bone volume per
bFGF: basic fibroblast growth factor; BM-MSCs: bone marrow-derived mesenchymal stem cells; DAPI: 4,6-diamidino-2-phenylindole; (D) MEM: (Dulbecco ’ s) modified Eagle ’ s medium;
3 ′ -UTR: 3 ′ Untranslated region; ALP: Alkaline phosphatase; DMP-1: Dentin matrix protein 1; DSPP: Dentin sialophosphoprotein; FACS: Fluorescence- activated cell sorting;
AIM: Adipogenic induction medium; ALP: Alkaline phosphatase; AP2: Adipocyte protein 2; ATSC: Adipose tissue-derived MSC; BMSC: Bone marrow-derived MSC; CEBP α :
ALP: Alkaline phosphatase; BM: Bone marrow; BMSC: Bone marrow-derived mesenchymal stem cell; CIA: Collagen-induced arthritis; DMSO: Dimethyl sulfoxide; FBS: Fetal bovine serum;
