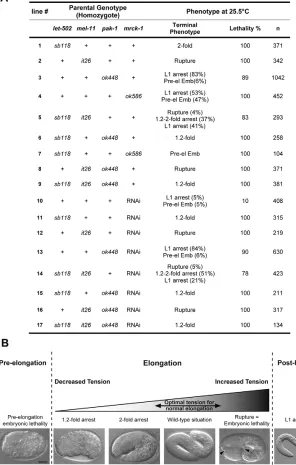
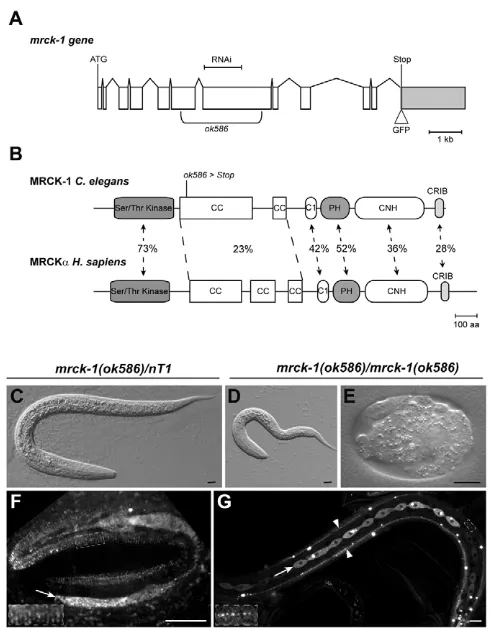
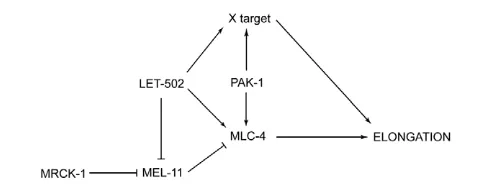
Myosin II regulation during C elegans embryonic elongation: LET 502/ROCK, MRCK 1 and PAK 1, three kinases with different roles
Full text
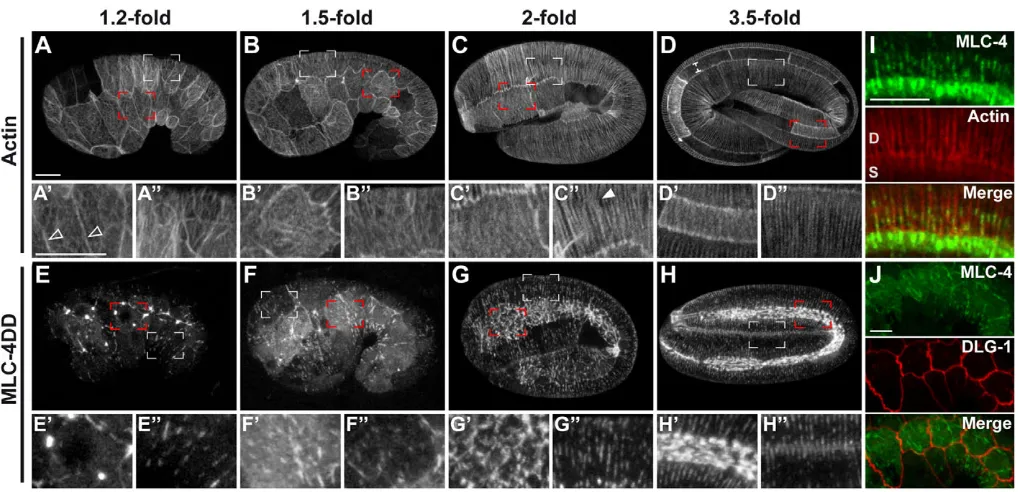
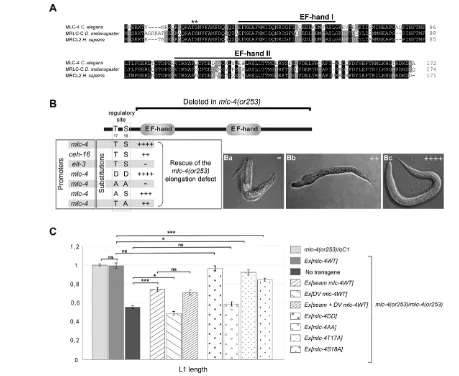
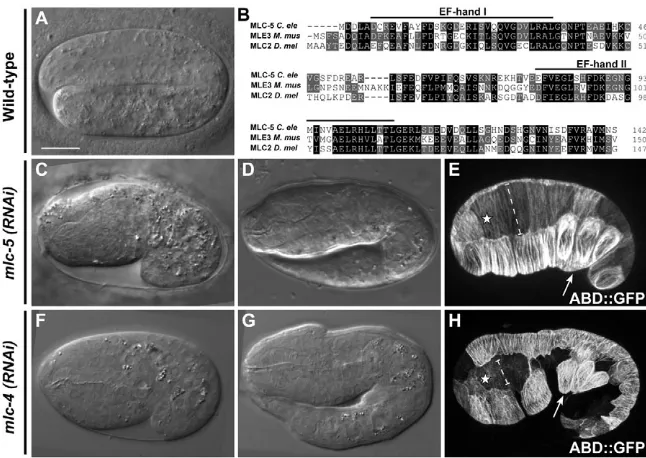
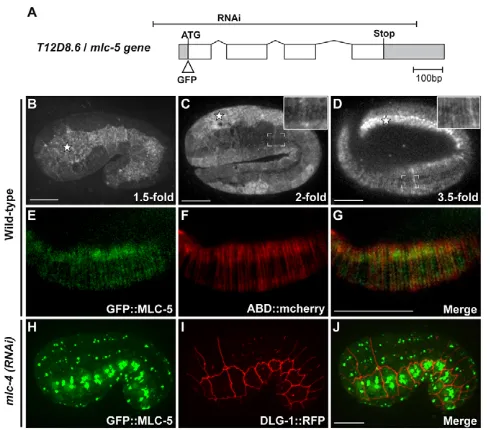
Figure
Related documents
The SNP position in the coding region and non-coding regions were then determined by using CLC Sequence Viewer 6.0 for all the 15 partial NmSusy1 genomic sequences (Table
The findings of the study reveal that gender and experience influence the Values in school- teachers although qualification influence the theoretical and aesthetic dimensions
normal levels of blood sugars in infants of.. diabetic mothers or
Abstract — This paper uses Multi-Layer Perceptron Neural Network (MLPNN) for comparing the linear dimensionality reduction techniques (DRTs) for fault diagnosis
Recen- temente, o Aphasia Rapid Test (ART) 11 foi proposto como um teste de cabeceira para avaliar a gravidade da afasia, baseando-se nos parâmetros neurológicos que são
and consider alternative options including oropharyngeal airway, nasopharyngeal airway, laryngeal mask airway, transtracheal jet ventilation, cricothyrotomy, and awake
