Studies into the insecticidal activity and mode of action of monoterpenoid constituents of essential oils against the human louse, Pediculus humanus
Full text
Figure
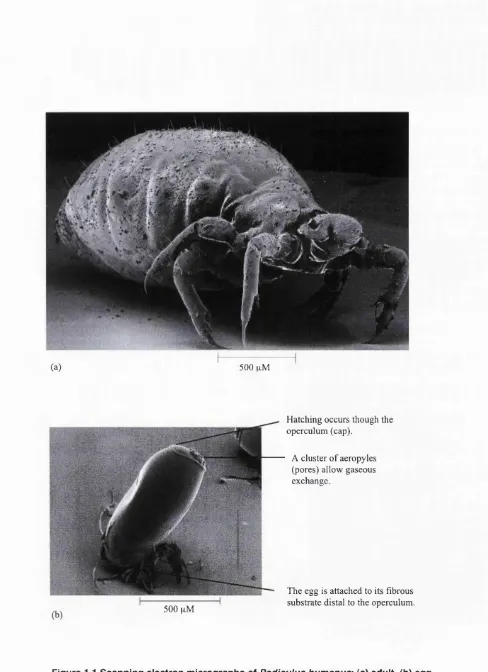
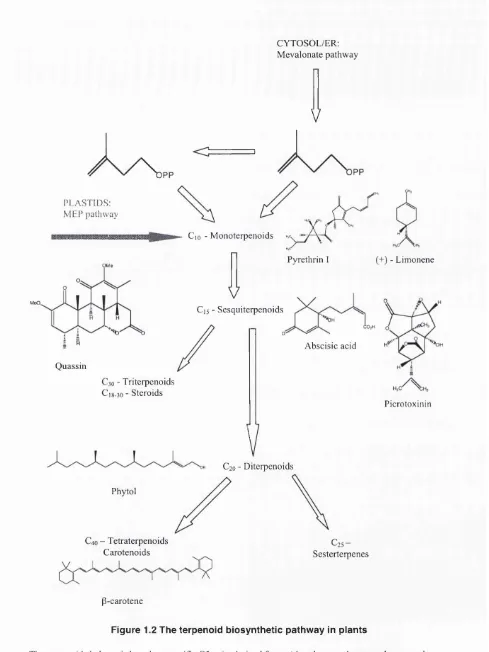
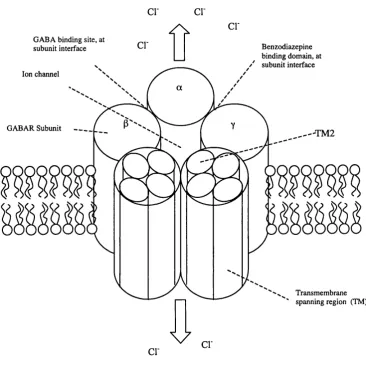
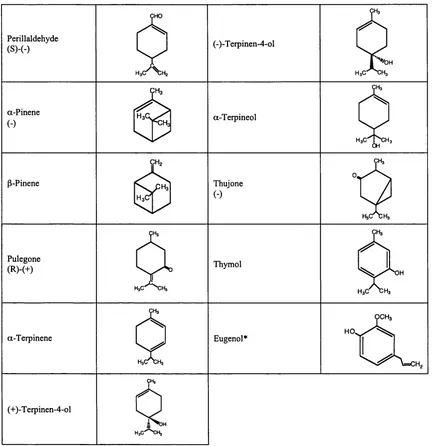
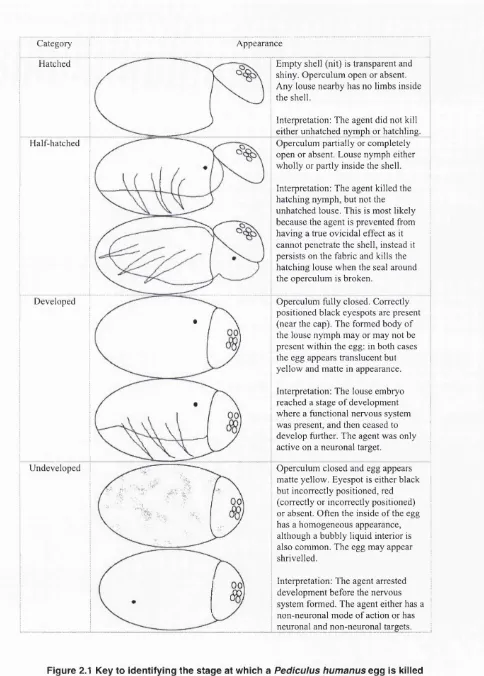
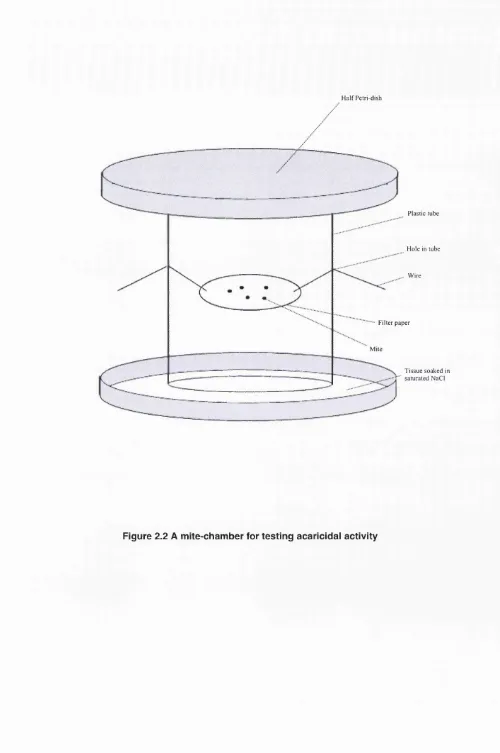
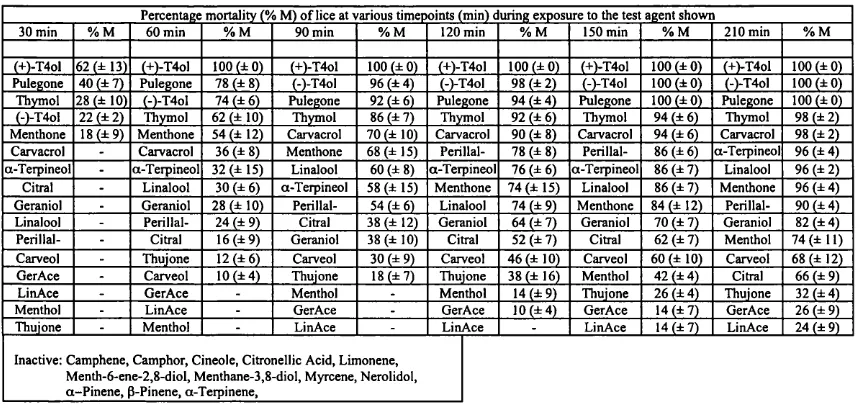
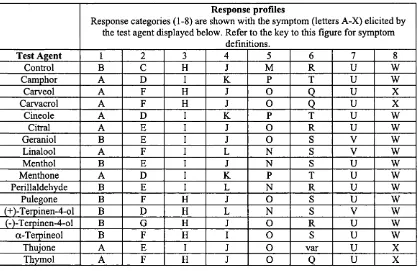
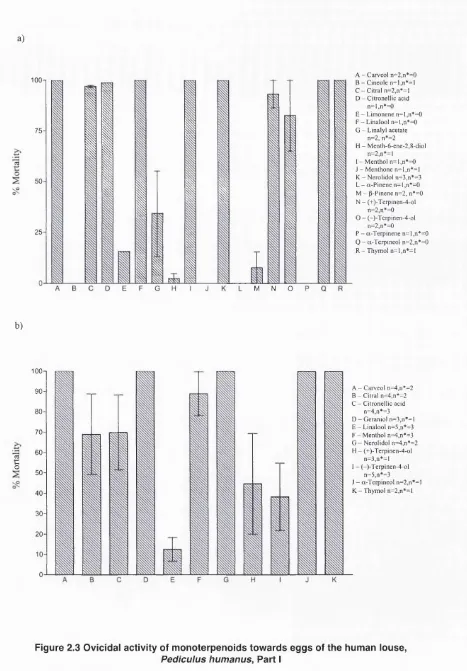
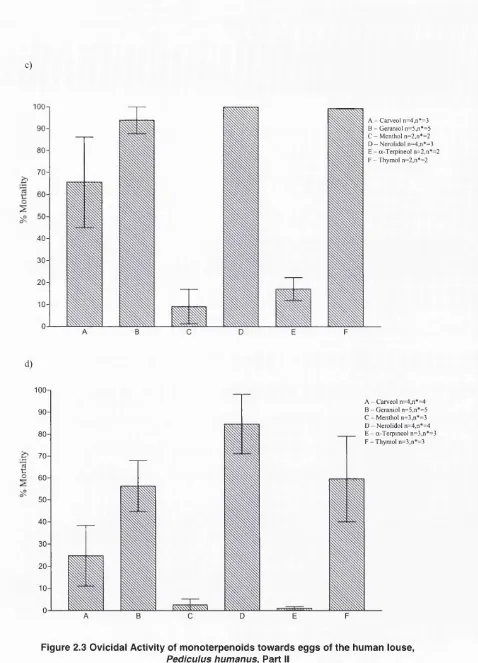
Outline
Related documents
The projected gains over the years 2000 to 2040 in life and active life expectancies, and expected years of dependency at age 65for males and females, for alternatives I, II, and
Twenty-five percent of our respondents listed unilateral hearing loss as an indication for BAHA im- plantation, and only 17% routinely offered this treatment to children with
• Low power and small die size of SAR present great potentials for spatial redundancy technique to be employed in single-event upset
(p-4) the measurement methodology used to determine information mentioned in maximum, idle, sleep, off mode power as defined in Point P9.1 in the Product IT Eco Declaration:.
19% serve a county. Fourteen per cent of the centers provide service for adjoining states in addition to the states in which they are located; usually these adjoining states have
To further examine whether the decreasing disease index in transgenic seedlings resulted from the silencing of VdAAC , we used qRT-PCR to investigate the relative expression level of
(37) also in effect argue for a phylogenetically based, unitary application of the name Fu- sarium, but they focus on a single clade nested within the terminal Fusarium clade where
