AFFECTING A QUANTITATIVE CHARACTER I N YEAST
W. R. SCOWCROFT AND A. P. JAMES*
Division of Plant Industry, CSIRO, Canberra, A.C.T., Australia
Received June 26, 1970
H E strategy applied to the genetic improvement of a population should in. clude knowledge of the nature of intra- and interallelic gene action. In this context the current theory of genotypic improvement assumes that dominance and epistasis are of little consequence. It has been claimed (ROBERTSON 1968) that the gene action extant in a population is a consequence of natural selection in the past; characters closely related to fitness primarily show nonadditive gene action, while genetic variability for traits of peripheral relevance to fitness are largely additive. Considerations of naturally occurring or newly induced genetic variability for characters such as viability, which are directly related to fitness, throw some light on the nature of gene action.
WALLACE
(1958, 1959, 1963),
CRENSHAW (1965), MUKAI and YAMAZAKI (1968),
andWILLS
(1968) have demonstrated that newly induced viability mutants show varying degrees of partial dominance. For example, MUKAI and YAMAZAKI (1968) found the average degree of dominance for newly arisen mutant polygenes controlling viability in Drosophila melanogaster to be of the order of 0.4.Such mutant viability genes also display a synergistic interaction in multiple homozygotes (MUKAI 196913) i.e., two mutants were more deleterious than if they acted independently. This nonlinear relationship between viability and gene number also holds for genotypes heterozygous for several recessive lethals
(KITAGAWA 1967).
In view of this evidence one might ask whether gene action of the more classi- cal quantitatively varying traits is similar to that found for viability genes. If this were true, then it may be deduced that nonadditive gene action is of consid- erable consequence, assuming that newly induced variability is similar to that which occurs in nature. With yeast, techniques have been developed (JAMES
1959) which allow the isolation of mutants in an otherwise homozygous back- ground and their subsequent precise measurement. In effect it is possible to remove the genes from a category which requires quantitative analytical tech- niques to one which permits the use of qualitative techniques.
The present study concerned (a) the isolation and characterization of a ran- dom sample of growth-rate mutants in an otherwise homozygous background; (b) the determination of their intra-allelic interactions; and (c) a n assessment
* Biology & Health Physics Division, C R N L , Chalk River, Ontario, Canada This research was done while A P J. was on leave in CSIRO, Canberra, A C T , Australia
60 W. R. SCOWCROFT A N D A. P. J A M E S
of interallelic interactions for all possible dihybrid combinations. The interactive models against which the dihybrid combinations were tested were:
( 1 ) Epistatic-the phenotype of the dihybrid is equal to that of the more detri- mental monohybrid contributing to the hybrid duplex.
(2) Additiue-the combined effect of the two mutant genes is the sum of their individual effects as monohybrids.
( 3 ) MuZtipZicatiue-the effects of the two mutant genes are completely inde- pendent.
The results indicated (i) that in general deleterious growth-rate mutants dis- play partial dominance and (ii) behavior of such mutants in dihybrid combina- tions suggests that they interact in a multiplicative fashion.
,
MATERIALS A N D M E T H ODS
Character: The character has been termed relative colony yield (yield). This metric was evaluated by measuring the diameter of the colonies produced by individual or budding cells after incubation f o r a specified time period. Yield is expressed as the mean diameter of the mutant colonies relative to the contemporary control wild-type genotypes. The diameter of a colony varies directly with the number of cells, and the time period chosen conforms to the exponential growth phase for which the plot of log colony diameter is essentially linear with time (JAMES, MACNUIT and MORSE 1965).
The yeast strain. A maximum degree of homozygosity a t loci other than those under investi- gation was essential to the experimental procedure. This requirement was satisfied by using a homothallic strain of yeast, Saccharomyces cereuisiae var. ellipsoides. In this strain the four spore-colonies produced after sporulation and dissection are diploid rather than haploid, and are homozygous at all loci other than sex. In fact, the four meiotic products germinate as haploids but, under the influence of a dominant gene D, conversion a t the sex locus occurs at an early vegetative generation and is quickly followed by the production of a diploid zygote (HAWTHORNE 1963). The system is particularly advantageous since accumulated heterozygosity resulting from spontaneous mutations can be eliminated at any time by resporulation and extraction of indi- vidual diploid spore colonies.
set of values. Finally, the segregation from the dihybrids between these four mutants in each case conformed to that expected from allelic segregation. The data from these four presumptive mutants have been pooled to provide greater accuracy i n the evaluation of the spontaneous mutant. The induced mutants are identified as a, b, c, e, k, and h, while the spontaneous mutant is identified as s.
Production of dihybrids: Matings to produce dihybrids were achieved by placing haploid spores of the desired genotypes adjacent to each other on agar medium. In a fraction of instances mating reactions were followed by the obvious production d diploid zygotes, and these were isolated as heterozygotes. This procedure was completely successful despite the seeming possi- bility that supposed heterozygotes were in fact homozygotes derived from single spores. The reliability of the technique was tested by crossing a pink adenine-dependent strain with white adenine-sufficient strains. Nine of ten matings were successful, producing white diploid lines which in turn produced white and pink segregants i n the ratio of 2:2. The exception was a single zygote which produced a pink and white mosaic colony. The white sector proved to be homozygous for adenine sufficiency and the mosaicism was attributed to a mitotic crossover soon after zygote formation.
Experimental design and analysis: Relative colony yield of the variants was determined by incubating the cells on the surface of slabs of chemically defined solid media (JAMES and SPENCER 1958) on glass microscope slides. The slabs were prepared using the apparatus designed by HAEFNER (1967). Six genotypes were placed on each slide. These were wild type, i.e., the homozygous dominant, three monohybrids, and two of the three possible dihybrids. The geno- types were randomized in six rows in each of four blocks per slide. Each row contained 10-15 single or budding cells isolated by micromanipulation from a colony in logarithmic phase of growth. The slides were sealed in the inverted position over a plexiglass box and incubated a t
30°C. At 3 hr, the number of cells per clone was noted. Diameters of the clones were measured to the nearest 0.05 mm at 27 h r using a micrometer eye piece.
The colony diameter a t 27 hr is influenced by the mitotic phase of the progenitor cell, i.e., the average correlation of diameter at 27 hr with cell number per colony a t 3 h r was 0.77 zk 0.01. Consequently, each genotype per slide was adjusted according to the mean cell number a t 3 h r for that particular slide. It is possible that standardization of colony diameter may partially obscure real genotypic differences. Some genotypes did in fact have significantly different cell numbers at 3 hr, but this was not consistently so from slide to slide.
Each slide presumably presents a unique environment to the genotypes thereon. Of the total variation displayed by the control genotype both within and between slides, 94.9% was attributed to variation between slides. Consequently, colony diameter of the mutant genotypes on a given slide was evaluated relative to their contemporary control genotype.
RESULTS
Characterization of mutant alleles: The seven homozygous mutant alleles caused a reduction in yield ranging from 8% to 46% (Table 1 ) . All mutants were significantly different from the control and from each other. The induced mutants closely resembled wild type in colony shape and outline. There was some variation in the spontaneous mutant, yielding some colonies which tended to be ragged in outline rather than smooth.
62 W. R. SCOWCROFT A N D A. P. J A M E S
TABLE 1
Relative colony yield of homozygotes and heterozygotes for the isolated mutants and the degree of dominance (d/a+) displayed b y each
Homozygotes Heterozygotes
Locus (Het) d/dr
C 0.748 t 0.0096 1.006 i 0.0024 (12)' 1.05
B 0.874 k 0.0064 1.001 i 0.0036 (8) 1.02
A 0.925 2 0.0074 1.002 t 0.0035 (8) 1.05
K 0.625 2 0.0127 0.986 +- 0.0024 (9) " 0.93
E 0.712 t 0.0114 0.815 2 0.0043 (11) -0.28
H 0.541 t 0.0030 0.811 t 0.0063 (9) 0.18
S 0.559 -C 0.0088 0.781 i 0.0024 (41) 0.01
* Figures in parentheses are number of estimates contributing to the mean 2 Het - (1
+
Hom)1 - Hom
+
See FALCONER (1960) : calculated asnant, while the spontaneous variant, s, displayed additive gene action.
The two remaining mutants, c and a, are of particular interest because of the overdominance displayed by the heterozygote. This is particularly strong for c since the mean yield of the heterozygote was significantly greater than wild type even though the degree of overdominance was only 0.6%. In the case of a, the mean yield of the heterozygote was statistically the same as wild type, so that any overdominance is probably more apparent than real. It is possible that such cases of overdominance are due to linked mutations having an effect which re- sults in a greater yield in the heterozygote. If so, the linkage was relatively tight since it was not broken in several segregations. In any case, gene action for c and possibly a was functionally single-locus overdominance.
An inspection of Table 1 indicated that there is no relationship between the effect of the yield mutant in the homozygous condition and its performance as a heterozygote. Mutant c, although depressing the yield by 25
%
as a homozygote, was overdominant in the heterozygote. This differs from the completely reces- sive mutant b, for example, which only depressed the yield by 13% when homo- zygous. This contrasts with JAMES' (1960) more extensive analysis in which he found a negative relationship-less detrimental alleles displaying a greater pro- portion of their effect in the heterozygote.63
TABLE 2
Relative colony yield of all possible dihybrids between the seuen isolated mutants
Ao Bb K k Ee H h ss
Cc 1.000 t .0084 1.012 t ,0076 0.999 t .0066 0.834 t .0070 0.84-1. -I: .0074 0.773 t . O O B
Aa 0.965 i: .0085 0.970 f .OO?j3 0.807 t ,0096 0.824 t .0032 0.778 f .0071
Bb 0.954 t ,0070 0.806 +. .0103 0.815 rt .0051 0.773 f .0028
K k 0.818 .0077 0.799 zk .0071 0.773 k .0081
Ee 0.705 k .0071 0.674 k .0119
H h 0.661 t BO28
spection of Table 3 and the magnitude of the standard errors about the means that it would be virtually impossible to construct a general model that would account for all dihybrid combinations. It is difficult to discriminate between the models where the mutants contributing to the dihybrid (cy a, by and h ) behave essentially as complete recessives. This of course would be expected if there were no interaction between the mutants. Where there is a large effect of the mutant in the monohybrid (mutants e, h, and s), it is obvious from Table 3 that the multiplicative model provides a better fit to the dihybrid data than the additive model which in turn is superior to that of the epistatic. Although a significant deviation of observed from expected was found only in the case of the epistatic model, the additive model gave a deviation some 28 times greater than the multi- plicative model. In general therefore, the relative colony yield of genotypes heterozygous at two loci is best described by the product of the respective mono- hybrids.
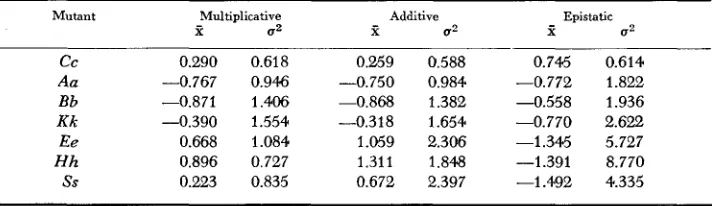
Intervariant homogeneity: Given that there is a spectrum of gene effects among these seven variants, it is pertinent to inquire as to the homogeneity of relative yield for each mutant (Table 4). The mean standardized deviation and variance refer to the mean effect of the nominated mutant in all possible dihybrid combi- nations with the other six and the variation about that mean, respectively, for each of the three models. Considering all mutants, the multiplicative model pro- vided the greatest amount of homogeneity as evidenced by the smaller deviations and relatively smaller variances. This is most apparent for the considerably smaller mean deviations of the heterozygotes Ee, H h , and Ss, for which the rela- tively smaller variances of the multiplicative model indicate a more consistent deviation from expectation. The multiplicative model was not ill-suited to any of the variants and was superior in those instances in which the data could be expected to discriminate between the models.
The heterotic nature of Cc>s supported by the fact that it maintains its heterosis for relative colspy yield independently of the background genotype. The mu- tants which were completely recessive in the monohybrid condition, a or b, or near recessive, IC (see Table I ) , each depressed the relative growth rate relative to expectation when in combination with other heterozygotes. Mutants e, h, and s,
64 W. R. SCOWCROFT A N D A. P. JAMES
TABLE 3
Standardized deviation' of relative yield of individual dihybrids from their expected values for the three modelse
An Bb Kk Ee Hh ss
M
c c A
E M
Aa A
E M
Bb A
E
M
K k A
E M
Ee A
E
M
Hh A
E
-0.52 0.33 0.56
-0.52 0.33 0.56
-0.12 0.83 1.31
-2.32* -1.45 --2.32* -I.& -2.80** -2.03'
-2.36* -2.34* -3.22* *
0.81 0.73 1.16 -0.53 - 0 . 5 1 -0.48 -0.52 -0.50 -0.51 0.83 0.97 0.19
1.30 -0.74 1.23 -0.77 1.60 -0.31 0.44 -0.23 0.51 -0.22 0.68 0.12 0.14 -0.50 0.09 -0.47
-0.05 0.13 0.09 0.26
2.14* 1.28 3.19** 2.48* -5.24*** -3.19** 1 .'lo 2.76* * -4.96*** 0.17 -0.34
-0.60 -0.27
Standardized deviation = [observed - expected] /SE. [obs-expl.
2 M multiplicative; A additive; E
=
epistatic.* P<0.05; ** P < O . O l ; * * * P<O.OOl.
DISCUSSION
From the study of a random sample of seven mutants (six induced by ionizing radiation and one spontaneous) with altered colony yield in yeast, the following generalizations may be made. (1) Mutants depress the relative colony yield (as measured by relative colony diameter) from 7.5% to 46% of wild type. (2) The
TABLE 4
Homogeneity of relative yield for each heterozygous mutant for the three models
Mutant Multiplicative Additive Epistatic
t 0 2 t 0 2 X a2
c c 0.290 0.618 0.259 0.588 0.745 0.614
Aa -0.767 0.946 -0.750 0.984 -0.772 1.822
Bb -0.871 1.406 -0.868 1.382 -0.558 1.936
K k -0.390 1.554 -0.318 1.654 -0.770 2.622
Ee 0.668 1.084 1.059 2.306 -1.345 5.727
Hh 0.896 0.727 1.311 1.848 -1.391 8.770
ss 0.223 0.835 0.672 2.397 -1.492 4.335
65
behavior of such mutants in heterozygotes covers the spectrum of gene action from overdominance through strict additivity to incomplete dominance of the mutant. ( 3 ) All possible dihybrid combinations suggest that in heterozygotes, relative colony yield mutants act independently, i.e., in a multiplicative fashion. There are several limitations in this study which must be taken into account when considering the general nature of quantitative genetic variability. The sample size was small, though ROBERTSON (1968) suggests that the number of loci that are responsible for the difference between extreme lines selected for a given quantitative character may be no more than ten. Consequently, it could be argued that the number of mutants studied in this sample might be characteristic of that encountered in a selection program.
Six of the seven mutants were induced by ionizing radiation and hence may not be a representative sample of those which occur in nature. Both completely recessive mutants and those exhibiting partial dominance with respect to fitness have been induced in both Drosophila and yeast. The mutants we have investi- gated would appear to fall into this category. Despite these weaknesses the pre- cision with which relative colony yield can be evaluated by the technique employed could prove valuable in characterizing quantitative genetic variability. Although the mean deviation in yield from that expected of a multiplicative model is relatively small, it is evident that the model is not exact, but only pro- vides a prediction which is superior to models based on either additive gene action or epistasis. In the case of the recessive mutant genes a and b, the multiplicative model overestimates the dihybrid yield. On the other hand it is clear from Tables 3 and 4 that, in general, the model provides underestimates of the yields of di- hybrids involving combinations of mutants e , h, and s, which had a distinct de- pressing effect as monohybrids. This pattern of deviation invites the interesting speculation that dihybridization tends to increase selection pressures against minor variants, and conversely, to decrease the selection intensity that can be applied to major mutants. Although MUKAI’S (1969a,b) evidence suggests that for multiple homozygotes, two or more mutants were more deleterious than if they were acting independently, our limited study indicates that deleterious mutants, at least as heterozygotes, act independently. I n fact the data, particu- larly those involving mutants e, h, and s, suggest that the effect of a mutant as a
heterozygote is mitigated by the presence of another deleterious mutant.
SUM MARY
The intra- and interallelic behavior of seven mutants affecting colony yield was determined in the homothallic strain of yeast Saccharomyces cerevisiae var. ellipsiodes. All mutants depressed colony yield as homozygotes. In the heterozy- gous condition the mutants each displayed either overdominance, complete recessiveness, incomplete recessiveness, additivity, or partial dominance. The
66 W. R. SCOWCROFT A N D A. P. J A M E S
L I TE R AT UR E CITED
CRENSHAW, J. W., 1965
FALCONER, D. S., 1960 Introduction to Quantitatiue Genetics. Ronald Press, New York. HAEFNER, K., 1967 A simple apparatus for producing agar layers of uniform thickness for
microbiological micromanipulator work. Z. Allg. Mikrobiol. 7 : 229-231.
HAWTHORNE, D. C., 1963 A deletion in yeast and its bearing on the structure of the mating- type locus. Genetics 48 : 1727-1 729.
JAMES, A. P., 1959 The spectrum of seventy of mutant effects. I: Haploid effects in yeast. Genetics 44: 1309-1324. -, 1960 The spectrum of seventy of mutant effects. 11: Heterozygous effects in yeast. Genetics 45: 1627-1648.
The influence of dose on the spectrum of radiation-induced mutants affecting a quantitative character in yeast. Genetics 52: 21-29.
The process of spontaneous extranuclear mutation in yeast. Genetics 43: 317-331.
Interaction in fitness between lethal genes in heterozygous condition in
Drosophila melanogaster. Genetics 57 : 809-820.
Maintenance of polygenic and isoallelic variation in populations. Proc. 12th Intern. Congr. Genet. 3: 293-308.
-
, 1969b The genetic structure of natural popu- lations of Drosophila melanogaster. VII: Synergistic interaction of spontaneous mutant poly- genes controlling viability. Genetics 61 : 74.9-761.The genetic structure of natural populations of Drosophila melanogaster. V: Coupling-repulsion effect of spontaneous mutant polygenes controlling viability. Genetics 59: 513-535.
The spectrum of genetic variation. pp. 5-16. In: Population Biology and Euolution. Edited by R. C. LEWONTIN. Syracuse Univ. Press, Syracuse, N.Y.
The average effect of radiation-induced mutation on viability in Drosophila melanogaster. Evolution 12 2 532-556, - , 1959 The role of heterozygosity in Dro-
sophila populations. Proc. 10th Intern. Congr. Genet. 1: 408419. - , 1963 Further data on the overdominance of induced mutations. Genetics 4.8: 633-651.
Yeast partial dominance : effect of environment and background genotype. Radiation-induced increases in fitness in the flour beetle Tribolium confusum. Science 149: 42f5-4-27.
JAMES, A. P., M. M. MACNUTT and P. M. MORSE, 1965
JAMES, A. P. and P. E. SPENCER, 1958 KITAGAWA, O., 1967
MUKAI, T., 1969a
MUKAI, T. and T. YAMAZAKI, 1968
ROBERTSON, A., 1968
WALLACE, B., 1958
WILLS, C., 1968