Copyright 0 1995 by the Genetics Society of America
Mutations Affecting Donor Preference During Mating Type
Interconversion
in
Saccharomyces cereuisiae
Karen
S. Weiler,’
Lisa Szetoand
James R. Broach
Department of Molecular Biology, Princeton University, Princeton, New Jersey 08544-1014 Manuscript received October 26, 1994
Accepted for publication December 15, 1994
ABSTRACT
Homothallic strains of Saccharomyces cereuisiae can convert mating type from a to a or (Y to a as often as every generation, by replacing genetic information specifjmg one mating type at the expressor locus, MAT, with information specifymg the opposite mating type. The cryptic mating type information that is copied and inserted at MAT is contained in either of two loci, H M L or H M R . The particular locus selected as donor during mating type interconversion is regulated by the allele expressed at MAT. MATa cells usually select HML, and MATa cells usually select H M R , a process referred to as donor preference. To identify factors required for donor preference, we isolated and characterized a number of mutants that frequently selected the nonpreferred donor locus during mating type interconversion. Many of these mutants were found to harbor chromosome rearrangements or mutations at MAT or H M L that interfered with the switching process. However, one mutant carried a recessive allele of C H L l , a gene previously shown to be required for efficient chromosome segregation during mitosis. Homothallic strains of yeast containing a null allele of CHLl exhibited almost random selection of the donor locus in a MATa background but were normal in their ability to select H M R in a MATa background. Our results indicate that Chllp participates in the process of donor selection and are consistent with a model in which Chllp helps establish an intrinsic bias in donor preference.
C
ELLS of the yeast Saccharomyces cermisiae are able to alternate efficiently between two distinct cell types. The cell type of a haploid yeast cell is determined by which of two different alleles is present at the mating type locus, MAT, on chromosome ZZZ. Cells carrying the MATa allele have an a mating type, while cells car- rying the MATa allele have an a mating type. MATa and MATa contain allele-specific sequences that en- code distinct transcription factors responsible for regu- lating the sets of a- and a-specific genes that determine the mating phenotype of the cell (reviewed in HERSKO- WITZ et al. 1992). Each cell also retains single copies of the a and a mating type genes at two other loci, HMR and H ML , located near the right and left ends of chro- mosome 111, respectively (HAFLMHIMA et al. 1974; NAS MYTH and TATCHELL 1980; STRATHERN et al. 1980). The mating type genes resident at these loci, though, do not contribute to the phenotype of the cell because they are maintained in a transcriptionally inactive state (KLAR et al. 1981; NASMYTH et al. 1981). The function of these silent loci is to serve as templates for gene conversion of the MAT locus during a mating type switching event (OSHIMA and TAKANO 1971; HICKS et al. 1977b; reviewed in HERSKOWITZ et al. 1992).Mating type switching occurs in haploid cells when
Corresponding author: James R. Broach, Department of Molecular Biology, Lewis Thomas Laboratory, Princeton University, Princeton, NJ 085441014. E-mail: [email protected]
Present address: Department of Zoology, NJ-15, University of Wash- ington, Seattle, WA 98195.
Genetics 139: 1495-1510 (April, 1995)
the allele sequence present at MATis replaced with the opposite allele sequence copied from HMRa or HMLa. Only strains carrying the HO gene (homothallic cells) are capable of mating type interconversion, and these cells can switch as often as every generation (HICKS and HERSKOWITZ 1976; HICKS et al. 1977a; STRATHERN and HERSKOWITZ 1979). Switching is initiated when the product of the HO gene, a site-specific endonuclease, catalyzes a double-strand DNA cleavage within the MAT locus ( STRATHERN et al. 1982; KOSTRIKEN et al. 1983; KOSTRIKEN and HEFFRON 1984). The double-strand break at MAT is then repaired by gene conversion, us- ing either HMLa or HMRa as the template, and results in the unidirectional transposition of the mating type information resident at the donor locus to MAT.
1496 K. S. Weiler, L. Szeto and J. R. Broach
dom. If it were, then productive switching, leading to a change in mating type, would occur no more than 50% of the time. That switching usually leads to a change in mating type indicates that cell type dictates donor locus selection. MATa cells preferentially utilize HMLCV as a donor locus, and MATa cells preferentially utilize HMRa as a donor locus. The mechanism by which the cell distinguishes the two donor loci during interconversion is not known (WEILER and BROACH 1992 )
.
Mutational analyses of the MAT locus suggested that a product of MATa, the a 2 protein, directly or indi- rectly specifies which donor locus will be selected. RINE
et al. ( 1981 ) showed that donor locus selection was wild type in a strain in which the a 1 protein was mutationally inactivated. Similarly, JENSEN and HERSKOWITZ (1984) showed that the product of MATa, the a1 protein, was not required for proper donor locus selection. They proposed a role for the a2 protein in donor locus selec- tion based on observations by TANAKA et al. (1984) of a hmku2 HMRa HO strain. TANAKA et al. (1984) deter- mined that a culture of this strain was composed of -90% mat412 cells and -10% MATa cells. These data led JENSEN and HERSKOWITZ (1984) to conclude that
the a 2 protein promoted the selection of HMRa and that H M h was selected in the absence of the a 2 pro- tein (in MATa or m a b 2 cells). Therefore, either the a 2 protein is a component of the interconversion ma- chinery or it regulates the expression of one such com- ponent in its capacity to repress transcription of a-spe- cific genes.
We have attempted to identify genetically some of the other transacting factors required for donor selection. Previous attempts to do so have not yielded informative mutants. These previous mutant isolation schemes were founded on the observation that a spore clone from a wild-type homothallic strain ( H M k HMRa) yields a nonmating colony, since the spore clone rapidly pro- duces equal numbers of a cells and a cells that conju- gate to form sterile a / a diploids. Consequently, the general approach undertaken independently in several laboratories was to screen for mutant isolates whose spores gave rise to colonies that still contained a signifi- cant number of mating-competent cells. These screens yielded mutations in the HO gene and genes whose products regulate HO expression ( HABER and GARVIK 1977; OSHIMA and TAKANO 1980; STERN et al. 1984), mutations at MAT that reduced or inhibited HO endo- nuclease cleavage ( MASCIOLI and WER 1980; OSHIMA and TAKANO 1980; HABER et al. 1980), gene conversions of one donor locus to the opposite mating type allele (OSHIMA and TAKANO 1980) and mutations causing cell type-specific sterility ( OSHIMA and TAKANO 1980). Noticeably absent from the collection of mutants were mutations in genes whose products were directly in- volved in the interconversion process, including RAD51, RAD52 and RAD54, that are required for com-
pletion of mating type interconversion (MALONE and
ESPOSITO 1980; FRIEDBERG et al. 1991 )
.
Nor did a direct screen of known UV-sensitive strains yield mutants de- fective in mating type switching ( HABER et al. 1980).We report here the results of a novel screen to iden- tify components of the cellular machinery responsible for mating type interconversion. Specifically, we sought mutants that frequently selected HMRa as a donor locus when cell type dictated that H M h be used as the donor template. Analogous to previous screens, some of the mutants isolated carried mutations at MAT or H M L , and mutants with chromosomal rearrangements were found at high frequency. However, one isolate carried a mutation, ddpl-1, that appears to perturb proper donor locus selection. DDPl is allelic to a previously identified gene, CHLl ( HABER 1974; identified as C F l by SPEN-
CER et al. 1990)
,
and inactivation of the gene has a clear and distinct effect on the fidelity of donor selection during mating type interconversion.MATERZALS AND METHODS
Media and culture conditions: Solid and liquid medium were prepared as described by S H E W et al. ( 1979). Yeast strains were usually grown on YEPD. For transformations and prototrophic selection during matings, the cells were grown on synthetic minimal medium or synthetic medium supple- mented with uracil and all amino acids but those omitted for selection. a-Amino adipate plates were used for determining the mutation frequency to cells capable of using a-amino adipate as nitrogen source ( lys2 or lys5) after EMS mutagene- sis ( CHATTOO et al. 1979). For 5-fluoro-orotic acid (5-FOA) selection, cells were grown on synthetic medium plates supple- mented with 0.1% 5-FOA.
Plasmids and plasmid constructions: Plasmid afz46 was generously provided by S. FIELDS (FIELDS and HERSKOWITZ 1985). It contains the MFaI gene fused to bacterial lac2 at nucleotide 23 of the m a l coding sequence, in vector YCp50.
The construction of plasmid pKSW32 was described in WEILER and BROACH ( 1992) . It carries the MAT locus, bear- ing a deletion of the Ya-Z1 region marked by the LEU2 gene. Plasmid pKSW39 was derived from plasmid pKSW34 ( WEILER and BROACH 1992) by inserting the 1.1-kb URA? gene, as a BglII fragment, into the unique EcoRI site of the plasmid, eliminating both BglII and EcoRI restriction sites in the process. Plasmid pKSW34 carries the MATa,,,, locus and a lys2-inverted cassette for directing plasmid integration at the LYS2 locus during yeast transformation. Linearization of plasmid pKSW39 with BglII produces a fragment that is essen- tially the lys2 gene interrupted with MATcr,,,,, URA? and vec- tor sequence.
Plasmid pKSW70 was constructed by inserting the 1.8-kb BamHI fragment of the H I S 3 gene into the unique StuI site of plasmid X35/ 313 ( GAUDET and FITZGERAI.D-HAYES 1987), eliminating both restrictions sites. Plasmid X35/313 (gener- ously provided by M. FITZGERALD-HAYES) carries a 4.8-kb re- gion spanning E N ? , into which has been introduced the cen?X35 mutation. It also carries the URA? gene, inserted as a 1.1-kb Hind111 fragment adjacent to cen?X?5, to serve as a selectable marker for transformation. However, the insertion of the HZS? gene to produce plasmid pKSW70 inactivated the
URA3 gene.
Donor Preference Mutants 1497
the BamHI cleavage site at position +710, inserted as a BamHI fragment into the vector pUC18. This B a d 1 ho fragment was isolated from plasmid pKSW24, which contains HO sequence from the BspMI cleavage site at +240 to the BspMI cleavage site at +1737, inserted into the SmaI site of vector pUC18. The B a d 1 site within the pUC18 polylinker of plasmid pKSW24 and the BamHI site at +240 of the HO coding se- quence were utilized to subclone the ho sequence. The URA3 gene was then inserted as a 1.1-kb Hind111 fragment into the HindIII site to create plasmid pKSW72.
Plasmids pSG4 and pSG6 were obtained from P. HIETER. Plasmid pSG4 contains the 3' end of CHLl from the Xbal to the EcoRI restriction sites ligated to the 5' end of CHLl from the EcoRI to the SpeI restriction sites (GERRING et al. 1990) and inserted into the HZS3integrating plasmid pRS313 (SI- KORSKI and HIETER 1989). Plasmid pSG6 carries the same CHLl sequences inserted into the URA3 vector pRS316. Transformation of yeast strains with either plasmid digested with EcoRI results in replacement of the endogenous CHLl gene by the plasmid vector and selectable marker.
Yeast strains and strain constructions: All of the strains used in this study are listed in Table 1. Except as noted, they were constructed in this laboratory and derive from a common HO haploid precursor strain, KSW3-20C, the con- struction of which was described in WEILER and BROACH (1992). Strain KSW3-20C contains a frame shift mutation at codon 26 in the a 2 coding sequence at both HML and MAT.
Since the a2 mutation rendered cells sterile, strain KSWS 20C was composed of a population of mah2 cells and MATa
cells.
Strain KSW3-20C mj& 1-lacZwas constructed by transplacing the MFal gene with the mjkl-lac2 fusion derived from plas- mid afz46. Plasmid afz46 DNA was digested with BglII and HindIII, which cut the MFal gene in the 5 ' regulatory se- quence and within the coding sequence 3' to the lacZ gene, respectively, and was cotransformed into strain KSW32OC with vector YEp51, selecting for Leu+ transformants. YEp51 transformants bearing the mfal-LucZ fusion were identified using the ,&gal filter assay, described below, and confirmed by Southern analysis. One isolate bearing the mfal-laczfusion and from which the YEp51 plasmid had been lost by segrega- tion was saved as strain KSW3-20C mjkl-lad.
Strain KSW4, described in WEILER and BROACH ( 1992), was isogenic to strain KSW3-20C, except the hmh2 allele was replaced with the HMLa allele, which was subsequently trans- posed to the MAT locus by Hamediated mating type inter- conversion. Consequently, strain KSW4 persisted as a diploid. Strains A9D6/KSW4 and AlOGlO/KSW4 were made by mating cells of mutants A9D6 and AlOG10, respectively, with spores of strain KSW4.
Strain KSW13 was constructed from strain KSW3-2OC- matA::LEU2 ( WEILER and BROACH 1992), which differed from strain KSW3-20C in that part of the mah2 locus, includ- ing the a1422 promoter and the HO endonuclease cleavage site, was replaced with LEU2. Strain KSWl3 was obtained by transforming strain KSW3-20C-matA::LEU2 to Ura+ Lys- with BglII-digested pKSW39 DNA, which had MATainC and
URA3 inserted in the middle of lys2 coding sequence. Strains A2G4/KSWl3 and A5ElO/KSWl3 were con- structed by transforming the mutant haploid strains A2G4 and A5E10 with vector YEp57 to make them His+ and mating them to strain KSW13, by selecting for growth of the diploids on medium lacking histidine and leucine.
Construction of strain KSW25 was described in WEILER and BROACH ( 1992)
.
Strain KSW25 was isogenic with strain KSW4, except that it was haploid and had a deletion of the MATlocus ( matA::LEU2) and an insertion of MATainc at the LYS2 locus ( lys2::MATainC).
Strain A3ClO/KSW25 was made by mating mutant strain A3C10 with strain KSW25, selecting for Leu+ Lys+ diploids. This diploid was sporulated, and the progeny were screened for leucine and lysine prototrophies and mating competence.
As described in RESULTS, a spore clone of the desired genotype was retained as mutant strain KSW38 ( =A3ClO/KSW258A). Strain KSW38 was diploid due to Hamediated mating type interconversion.
Strain KSW29 was constructed by transforming strain KSWl3 to His+ with EcoRIdigested pKSW70 DNA, which re- placed CEN3 with cen3X35-ura3::HZS3.
Strains KSWS2OC/KSW29 and A3C10/KSW29 were made by mating strains KSW320C and A3C10, respectively, with strain KSW29, selecting for diploids on medium lacking lysine and histidine. Diploid colonies composed of cells having lost the copy of chromosome ZZZ bearing the ca3X35-ura3::HZS3 mutation and the matA::LEU2 allele were identified by their inability to grow on medium lacking histidine and leucine. A single monosomic isolate from each strain was saved as strain KSW41 ( =KSW3-2OC/KSW29 monosomic for chromo- some ZZZ) and strain KSW39 ( =A3ClO/KSW29 monosomic for chromosome ZZZ)
.
The two aneuploid strains were then subjected to 5-FOA selection to isolate recombinants having replaced the lys2::MATai,,,-URA3 allele with LYS2+ by gene conversion, using the LYS2 locus on the homologous chromo- some as a template. The fact that the 5-FOA resistant isolates mated as a cells confirmed that the desired recombination event had taken place. A single 5-FOA resistant isolate from strain KSW41 was retained as strain KSW42, and a single 5- FOA resistant isolate from strain KSW39 was retained as strain KSW40.Strain KSW30 was produced from a cross between spores of strains KSW38 and KSW4. Strain KSW30 was diploid as a consequence of Hamediated mating type interconversion, and only differed from strain KSW4 in being homozygous for the ddpl-1 (chll-200) allele.
Strains KSW32 and KSW33 were heterothallic haploid deriv- atives of strain KSW4. Strain KSW4 was transformed to Ura+ with DNA of plasmid pKSW72, linearized at the unique Ed7111 site within the ho sequence, to direct its integration at the HO gene. One transformant was sporulated, and the progeny from 10 tetrads were scored for uracil prototrophy and mating ability. The tetrads were observed to segregate 2:2 Ura+:Ura- with half of the Ura+ progeny exhibiting a
mating and half exhibiting a mating, confirming that the transformant carried the ho::CiRA3 disruption. One MATa
ho::URA3 spore clone was saved as strain KSW32, and one
MATa h o : : m 3 spore clone was saved as strain KSW33. Strains KSW30-B ( MATa ho::U&i3) and KSW30-C (MATa
ho::CilRA3) were derived from strain KSW30 and constructed in the same manner as described for strains KSW32 and KSW33. Strain KSW30-B/C was made by cell to cell mating of strains KSW30-B and KSW3O-C. Strain KSW30-C/KSW4 was made by spore to cell mating of strains KSW4 and KSW3OC.
Strains KSW34 and KSW35 are heterothallic derivatives of strain KSW3-20C. Strain KSW3-20C was transformed to Ura+ with DNA of plasmid pKSW72, linearized at the unique Eco47III site within the ho sequence, to direct its integration at the HO gene. Transformants bearing the ho::URA3 disruption were identified as stable MATa and matcu2 isolates, using the limited mating assay. A MATa ho::URA3 transformant was re- tained as strain KSW34 and a mah 2 ho::URA3 strain was retained as transformant KSW35.
Strains CTF1::TnlS and CTFl-R:Tn43 were constructed in the laboratory of P. HIETER and were provided by the labora- tory of M. ROSE. The CHLl alleles carried by these strains are described in GERRINC et al. ( 1990).
1498 K. S. Weiler, L. Szeto and J. R. Broach
TABLE 1
Experimental yeast strains
Strain Mating type loci Additional markers
KSW3-20C
KSW3-20C mfal-lacZ
KSw4 A9D6 KSw4 AlOGlO - KSw4 KSWl3 A2G4 KSWl3 A5E10 KSWl3 KSW25 A3C10 KSW25 KSW29 A3C10 KSW29 KSW3-2OC KSW29 KSW30 KSW30-B KSW30-C KSWSO-B/C KSW30-C KSw4 KSW32 KSw33 KSw34 KSw35 KSW38 KSW38 KSw4 KsW39' KSW40' KSW4 1 KSW42' CTF1::TnlSd CTFl-R::Tn43d
ma& 2/MATa h m h 2 HMRa ma& 2/MATa h m h 2 HMRa MATa HMLa HMRa MATa HMLa HMRa
-"
MATa h m h 2 HMRa" MATa HMLa HMRa
"
-MATa h m h 2 H M l W MATa HMLa HMRa matA::LEU2 h m h 2 HMRa
MATa h m h 2 HMRa matA::LEU2 h m h 2 HMRa MATa h m h 2 HMRa matA::LEUZ h m h 2 HMRa matA::LEU2 H M k HMRa
MATa h m h 2 HMRa matA::LEU2 HMLa HMRa matA::LEUZ h m h 2 HMRa
MATa h m h 2 HMRa matA::LEU2 h m h 2 HMRa MATa h m h 2 HMRa matA::LEUZ h m h 2 HMRa MATa HMLa HMRa MATa HMLa HMRa MATa HMLa HMRa MATa HMLa HMRa MATa H M h HMRa MATa HMLa HMRa MATa HMLa HMRa MATa HMLa HMRa MATa HMLa HMRa MATa HMLa HMRa MATa h m h 2 HMRa matcr2 h m h 2 HMRa MATa H M h HMRa MATa HMLa HMRa
MATa HMLa HMRa MATa HMLa HMRa
MATa h m h 2 HMRa
"
--__
" "-___
" " -"-"
-" -"-MATa h m h 2 HMRa
MATa h m h 2 HMRa
MATa h m h 2 HMRa MATa HMLa HMRa MATa H M k HMRa
ura3-52 leu2-3,112 his3Al trpl-289 HO As KSW3-2OC; mfa 1-lacZ
As KSw320c
As KSW3-2OC; mfa 1-lacZ
m a 1 mfa 1-lacZ
m a 1
As KSW3-2OC;
As KSW3-2OC; ~~s~::MATcu,,,-URA~
As KSW3-2OC; mfal-lacZ L YS2
m a l ~~S~::MATCU;,,,-URA~
As KSW3-2OC; m f i 1-lacZ L YS2
MFa 1 LYsZ::MATCU,,,,-URA~
As KSW3-2OC; L Y S ~ : : M A T O ~ ~ , ~
As KSw320c;
As KSW3-2OC; lys2::MATai,,,-URA3 cen3I05-HIS3 mfoll-lacZ LYS2 chll-200
MFa 1 lys2::MATainC CHLl
As JSSW3-2OC; rnfa 1-lacZ L YS2 Chll-200 CEN3 MFQ 1 ~YS~::MATCU,,,,-URA~ CHLl ~ ~ 3 2 3 5 - H I S 3
As KSw320c; L YS2 E N 3
lys2::MATai,,-URA3 cen3X35-HIS3
chll-2Ooh
AS KsW3-2OC: ~ chll-200
As KSW3-2OC; chll-200 ho::UmJb As KSW3-20C; chll-200 ho::Um3'
chll-200 ho::URAJb AS KSW3-2OC; ~ ~
chll-200 hO::URA3
As KSW3-2OC; ___ ___ chll-200 ho::URAJb CHLl HO
AS KSw320c; ho::Um3
AS KSW3-2OC; hO::URA3 AS KsW3-2OC; ho::URA3
As KSW3-2OC; hO::URA3 chll-200b
AS KSW3-2OC; ___ chll-200
AS KSW3-2OC; ~
chll-200" CHLl
As K s w 3 2 0 c ; rnfa 1-lacZ LYSZ ~hll-200 MFa1 lys2::MAT~ui,,-URA3 CHLl CEN3 mfal-lacZ chll-200
MFal CHLl L YS2
lys2::MATc~i,,,-URA3
As KSW3-2OC; CEN3
AS Ksw3-2oc;
AS JSSW3-20C; CEN3
CEN3
Donor Preference Mutants 1499
TABLE 1
Continued
Genotype Strain
Mating type loci Additional markers
LS204 MATa h m h l a Z,,, hmra,,, MATa hmlcv la.&,, hmrairrL
LS342 MATa h m h la Z,,,, hmra,,, MATa hmlala&,, hmra,,,
LS349 MATa hmlcvla Z,,, hmra,,, MATa hn&ku&,, hmrainc
As m w 4
As Ksw4; chll::URA3 CHLl
As Ksw4; chll::URA3 chll::HIS3
~
a Chromosome 111 is involved in a reciprocal translocation, as described in RESULTS.
'The presence or absence of the mfal-LucZ allele was not determined in the strain. "The strain is diploid, but monosomic for chromosome III.
The strain was obtained from the HIETER laboratory via the ROSE laboratory.
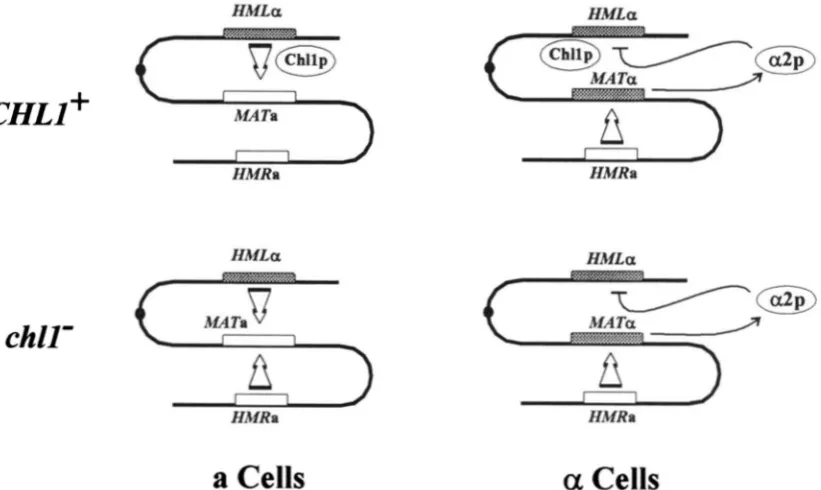
of the HMLLV loci with an hmlcvla~,, allele and replacement of the HMRa loci with an hrnq,, allele (L. SZETO and J. R. BROACH, unpublished results)
.
An inc allele is a mutation of the HO endonuclease cutting site; it does not affect the ability of HML or HMR to serve as a donor locus and is always transferred to MATafter HOinduced mating type conversion. Once resident at MAT, the inc mutation prevents any further HOinduced switching. The hrnlcvla.2 allele is a 290-bp dele- tion that eliminates the a l a 2 promoter, and the hmm allele is a 460-bp deletion within the a1 coding region.Strain LS342 was heterozygous for a disruption of CHLl and was derived by transformation of strain LS204 to Ura+ with EcoRIdigested pSG6 DNA.
Strain LS349 carried disruptions of both CHLl loci and was obtained by transforming strain LS342 to His+ with EcoRI digested pSG4 DNA, screening for transformants that also remained Ura+
.
Mutagenesis: Strain KSW320C m fa I-lacZ was grown over- night in YEPD liquid medium to a density of -1 X 10' cells per ml. Cells from 1.0 ml were mutagenized with EMS essen- tially as described (SHERMAN et al. 1979). The mutagenesis resulted in a 50% decrease in viability and a -20-fold increase in the frequency of cells capable of growing on a-amino adi- pate.
Mutant isolation: About 14,000 colonies grown on YEPD at 30" were screened by a limited mating assay and a p-galac- tosidase filter assay, both described in detail below, for pheno- types reflecting an increase in the fraction of MATa cells per colony as compared to the parental strain. Nine hundred seventy-two colonies qualified as potential mutants by at least one assay and were pursued further. Each isolate was cultured in 200 p1 YEPD in a microtiter dish, inoculated onto a YEPD plate using a multiprong applicator, grown overnight and then retested by both assays. Three hundred thirty-one iso- lates that were initially identified by both screens or exhibited a strong phenotype during a secondary screen were streaked out for single colonies and reexamined by the mating assay. Of these isolates, 140 appeared mutant and were further ana- lyzed by an a-factor confrontation assay to determine the percentage of a cells in cultures of each putative mutant. The sample size for the initial confrontation assays was -50 cells in early experiments and 100-125 cells in later experiments. Genetic techniques: Yeast cells were transformed by the LAC method of ITO et al. ( 1983), using the one step trans placement method of ROTHSTEIN (1983) for chromosomal gene replacements.
For spore to spore matings, each diploid was sporulated, and the asci were dissected on YEPD. Multiple mating pairs were arranged by micromanipulating two spores of unde- termined mating type, one derived from each diploid, adja- cent to each other. Each potential mating pair was exam- ined microscopically over several hours for the formation of a zygote, indicating successful diploid formation. Cell to spore and cell to cell matings were performed in an analo- gous manner.
A limited mating assay on plates provided discrimination between yeast strains with subtle differences in a mating abil- ity. A fresh lawn of strain DC17 ( MATa his1 ) cells was sequen- tially replica printed to two velvets, and the second velvet was then printed to a YEPD plate. The colonies to be tested were similarly printed to two velvets and the second velvet printed on top of the lawn of a cells. The cells were allowed to mate and grow without selection for 5 hr at 30", after which time they were replica printed to minimal medium for diploid selection. Differences in relative diploid growth were scoreable for a short window of time, after -12 hr of incuba- tion at 30".
a-Factor confrontation assays were used to quantitate the number of a cells present in a population of cells. A thick bisecting line of the a tester strain, DC17, was applied to a YEPD plate and incubated overnight. Single cells of the strain to be assayed were micromanipulated to within 0.5 mm of the mass of a cells. The plate was incubated at 30" for several hours, and then the cells were scored for their response to the a-factor secreted into the medium by the DC17 a cells. Cells that divided, i.e., were a-factor resistant, were scored as matcu2. Cells that were a-factor sensitive, as indicated by G1 arrest and shmoo formation, were scored at MATa. Cells that neither divided nor responded to the a-factor by shmooing were considered inviable.
Two types of pedigree analyses were performed to deter- mine the frequencies of switching from MATa to MATa and MATa to MATa. The first method relied on pheromone re- sponsiveness as a metric of the status of the MAT locus of
1500 K. S. Weiler, L. Szeto and J. R. Broach
cessful a to a switching was signaled by two cells at the four cell stage exhibiting growth arrest and shmoo formation. For MATa to h4ATa switches, tetrads were dissected on a YEPD plate impregnated with a crude preparation of a-factor. a spores were identified as those that continued to grow after germination. When such spores had divided once and both mother and daughter had budded, the two budded cells were placed near a source of a-factor and monitored microscopi- cally. A successful a to a switch was signaled by the arrest of
two of the four subsequent cells; the absence of a switch was signaled by the arrest of all four cells.
The second method for detecting mating type switches re- lied on PCR analysis of spore clones derived from strains carrying modified donor loci. These strains were constructed such that ( 1 ) the two donor loci alleles could be distinguished by PCR analysis from each other and from either allele ( a or
a ) initially resident at MAT, and ( 2 ) upon transfer of either of the donor locus alleles to MAT, the strain could not un- dergo further mating type interconversion. Sporulated test strains were dissected in the presence of a source of a-factor, and the mating type of individual spores was determined after 4 hr by microscopic examination of the response of each spore to the pheromone. Each spore that had either budded or exhibited anisotropic growth (shmooed) was removed from the source of a-factor and allowed to form a colony. PCR analysis was then performed on each spore clone to determine the allele present at MAT in the majority of the cells in each colony. This analysis readily distinguished among all the possible MAT alleles: a , a , a 1 and a l a 2 . Colonies predominantly exhibiting the mntal allele were scored as
selecting HMR as donor, colonies predominantly exhibiting matcvla2were scored as selecting H M L as donor and colonies that failed to exhibit one predominant allele were scored as ambiguous.
Molecular techniques: The P-galactosidase filter assay, adapted from the protocol of BREEDEN and NASMWH ( 1985), was used as a quick assay of yeast colonies (or patches) for P-gal activity. The plate of cells to be assayed was replica- printed to a velvet, which was used to print to a round, prewet- ted nitrocellulose filter. The filter was suspended over liquid nitrogen for 90 sec, immersed for 30 sec and then allowed to warm to room temperature. The filter was then placed in a
petri dish (cell-side facing upward) on top of a 3MM What- man filter presoaked in 2 ml Z buffer
+
X-Gal (625 pg) . The petri dish was covered, sealed and placed at 30” until the desired level of color was obtained.For karyotypic analysis of mutant strains by pulsed field gel electrophoresis, chromosomal DNA was isolated as described in ROSE et nl. (1989). One percent SeaKem GTG (FMC) agarose gels in 0.5X TBE were usually run in a CHEF appara- tus at 200 volts for 14 hr at a 60-sec pulse frequency, followed by 10 hr at a 90-sec pulse frequency. Gels were stained for 30 min in 0.5 yg/ml EtBr, destained for 1 hr and photographed under UV illumination.
PCR analysis to determine the allele resident at MA?’ in individual spore clones was performed by resuspending cells directly from the colony into PCR buffer and then running the PCR amplification as specified (Promega) . Primers were
5 “CTTGAGACGATTTGGCCCTG3
‘
and 5 “CCGTCCCGT- ATAGCCAATTCGJ’, corresponding to sites within W and centromere distal to MAT, respectively.RESULTS
We conducted a novel genetic screen for mutants that select the preferred donor locus with decreased fidelity during mating type interconversion. The most
important consideration in devising a new mutant isola- tion scheme to identify factors involved in donor prefer- ence was to require that the mutants be capable of switching. This requirement was best fulfilled by using a h m h 2 matcu2 HMRa HO strain, which undergoes futile cycling of the a 2 allele from HMI, to M T s i n c e mutcu2 strains preferentially select HML as donor. The mutant isolation scheme was based upon the following premise: a mutation in a gene whose product is required to select
H M h preferentially should result in the use of HMRa
as donor at increased frequency, compared to the par- ent. The mutant would be expected to transpose and express the a allele at MAT more frequently than the parental strain. T o identify such mutants, we screened colonies for those showing an increase in the fraction of a cells per colony. Mutations that only decrease the frequency of switching should not be recovered in the screen, since expression of the mutant phenotype re- quired that mutcu2 cells more frequently switch to MATa.
Isolation of mutants: T h e h m h 2 mutcu2 HMRa HO strain used for mutagenesis, KSW3-20C mfal-lucZ, was composed of 95% mutcu2 cells and 5% MATa cells (as determined by confrontation analysis described in MA- TERIALS AND M E T H O D S ) . Primarily due to the percent- age of a cells, but also to a partial a phenocopy of mutcu2 mutants, this strain exhibited a weak a mating character. This weak a mating ability was distinguish- able from the strong a mating ability of a MATa strain by a limited mating assay, as shown in Figure 1A. Al- though the difference in levels of mating observed when comparing strain KSW3-20C m fa 1-lacZand a n iso- genic MATa strain, KSW34, was not great, mutants showing a few fold more a cells than strain KSW3-20C
mfal-lucZ (having 5% a cells) were distinguishable from it by this limited mating assay (see below) . To increase the probability of identifying mutants, we si- multaneously employed a second screen for decreased P-galactosidase activity produced from the m fa 1-lucZ fusion present in the strain.
Strain KSW3-20C m,fal-lacZ was mutagenized and screened on YEPD at 30” for either an increase in a mating character by the limited mating assay o r a de- crease in P-galactosidase synthesis by the P-gal filter assay. As described in MATERIAIS AND METHODS, of the -14,000 colonies screened by both assays, 331 emerged as possible mutants. Of these, 140 appeared to exhibit greater a character than strain KSW3-2OC when exam- ined more closely using the limited mating assay. T h e mating ability of a subset of these mutants is shown in Figure 1B.
1502 K. S. Weiler, L. Szeto and J. R. Broach
TABLE 2
Mutants defective in selection of HML as donor locus
Mutant Phenotype Description of mutation
A5D4 97% a Translocation of chromosome III to
A5F7 80% a
A9D6
Translocation of chromosome /I/ to V
80% a Translocation of chromosome III to V
AlOGlO 80% a M E 1 0
Translocation of chromosome 111 to V
76% a MATaY8C-T
A2G4 42% a
A3C10
Chromosome III rearrangement
21% a chll-200 1/71 or XV
20C, and are listed in Table 2. Mutant A3C10, having 21% a cells, was included in this class since the first results from confrontation analysis suggested that it had 31% a cells. The higher initial value reflected the ambi- guity of distinguishing between inviable, anisomorphic cells and cells responding to a-factor. These mutants are described in more detail in the following.
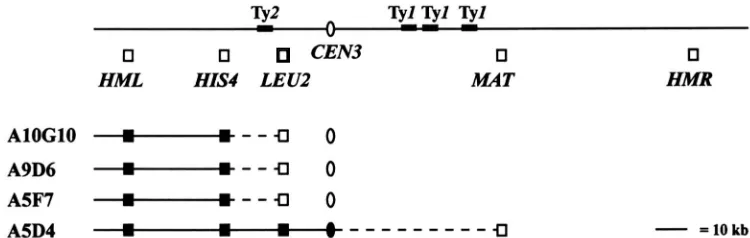
Translocations of the HML locus lead to a strong mutant phenotype: Analyses of four mutants having a strong a phenotype indicated that they carried translo- cations of H M L . First, heterozygous diploids obtained by crossing two of the mutants (A9D6 and AlOGlO) to strain KSW4 exhibited low ( -50% ) spore viability with the vast majority of the tetrads exhibiting a viab1e:invia- ble segregation pattern of 4:0, 2:2 or 0:4. Second, the two mutations were tightly linked to either h m h 2 or MATa (data not shown). Third, karyotype analysis by pulsed field gel electrophoresis demonstrated loss of H M L from chromosome ZZZ in all four strains and its appearance on a second chromosome (Figure 2 ) . From these data we concluded that mutants AlOG10, A9D6 and A5F7 all carried a ZZZ:Vreciprocal transloca- tion and mutant A5D4 carried a ZIZ:VZZor IIIXVrecipro- cal translocation. The translocation breakpoints of the four mutants were approximately located by probing the karyotype filter with DNA corresponding to differ- ent chromosome ZZZgenes, the results of which are sum- marized in Figure 3. The limits of breakpoint regions for all four translocations encompass several Ty and
6
elements and have been noted as hot spots for Ty transposition ( WARMINGTON et al. 1986, 1 9 8 7). Recom- bination events involving these repeated elements were probably responsible for the translocations.A chromosome ZZZ rearrangement involving the HML locus affects its activity as a donor: Mutant A2G4 (42% a ) had several unusual properties. First, chro- mosome
ZZZ
in mutant A2G4 carried additional DNA, roughly estimated at 100 kb (lane7,
Figure 2 A ) . This represents a duplication or insertion of genetic mate- rial into the chromosome, since MAT, H M L and H M R still reside on the enlarged chromosomeZZI.
The source of this extra material on chromosomeZZZ
is unknown. Second, the H M L locus had undergone a rearrangement, as judged by Southern analysis. Thenature of this rearrangement is unknown, although it is not a simple deletion. Finally, sporulation and dissection of diploids obtained between mutant A2G4 and strain KSWl3 (see Table 1 ) yielded only 2 : 2 via- b1e:inviable spore tetrads. Since the rnatA::ZEU2 allele from strain KSWl3 was inherited by all of the viable spores (data not shown), the diploid behaved as if it carried a haplolethal mutation tightly linked to the MATa allele derived from mutant A2G4. Obviously, though, this haplolethal phenotype was not manifest in the original mutant. We do not yet know which, if either, of these properties are responsible for the switching phenotype of the strain.
A mutation in Ya of MAT inhibits switching: Mutant A5E10 had both a strong mutant phenotype (76% a cells) and a wild-type karyotype (Figure 2, lane 6 ) . In addition, spores from diploids formed between mutant A5E10 and strain KSWl3 showed normal viability, and tetrad analysis from this cross indicated that the mutant phenotype cosegregated with the MATa locus (or Z,YS2) from mutant A5E10. Sequencing the MA79 al- lele from mutant A5E10 and from wild-type strain KSW3-20C revealed that the mutant had a single base change in the Ya region of MAT, eight nucleotides away from the Y/ Z1 border (Figure 4 )
.
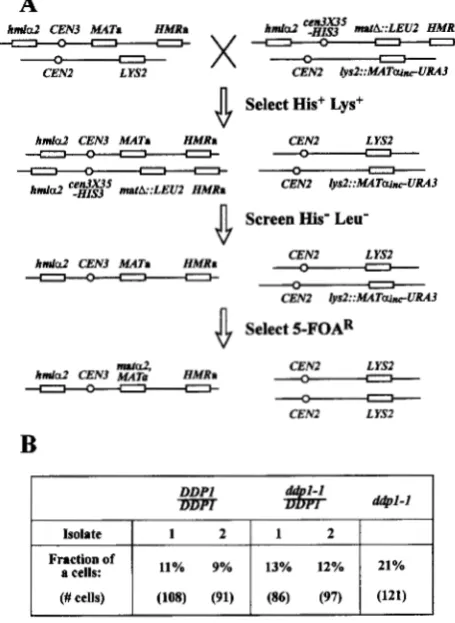
Since position Y8 is within the region recognized by the HO endonu- clease ( NICKOLOFF et al. 199 0 ) , we surmised that this mutation impaired the ability of the HO endonuclease to cleave MATa. Preliminary pedigree analysis data from a strain carrying HMLCY and this mutation at MA79 supported this conclusion.A recessive mutation, ddpl-1 (chll-200), reduces the fidelity of preferential donor locus selection: Charac- terization of mutant A3C10 (21% a ) indicated that it contained a recessive mutation, provisionally desig- nated d d p l - 1 , that affected donor locus selection. The strain appeared normal by PFGE analysis (Figure 2, lane 8 ) and typically yielded four viable spores per tet- rad when crossed to wild-type strain KSW13. As deter- mined by the complementation test detailed in Figure 5, the mutation responsible for the increase percentage of a cells was recessive. This analysis also showed that d d p l - 1 was not present on chromosome ZZZ.
Donor Preference Mutants 1503
A
B
1
2
3
1
4
2
5
3
6
4
7
5
8
6
7
8
chrom
I
C
D
E
FIGURE 2.-Analysis of mutant strains by PFGE reveals that sev- eral mutants carry chromosome III rearrangements. ( A ) Intact chromosomes were isolated from yeast cells and fractionated by pulsed field gel electrophoresis as described in MATERIALSAND METH- ODS. The ethidium bromide- stained gel is shown. The identity of the chromosome bands is indi- cated to the right. The strains are as follows: lane 1, KSW3-20C; lane 2, A5D4; lane 3, A5F7; lane 4, A9D6; lane 5, AlOG10; lane 6, A5E10; lane 7, A 2 G 4 ; lane 8, A3C10.
(B-E)
The gel shown in A was processed for Southern analysis and sequentially probed with the 1.4-kb HindIII-BamHI fragment centromere-proximal toHML
(B),
the HIS4 gene frag-ment ( C ) , the LEU2 gene frag- ment ( D ) and the EcoRI-Hind111 1.9-kb A region to the left of CEN3 ( E ) .
and, in addition, was masked in strains capable of
HO
notypesof
d d p l - 1 strains and those of chll (chromo- mediated mating type interconversion (Figure 6 ) . This some loss 1 ) strains, identified through mutations that chromosome loss phenotype was confirmed by the de- caused chromosome loss ( WER 1974; LIW et al. 1978; termination, described below, that d d p l is allelic to SPENCER et al. 1990), prompted us to test for allelism chll, mutations of which have been shown to cause of the two genes. The possibility of allelism was strength- mitotic chromosome loss. ened by the observation that both chll and d d p l - 1 ex-DDPZ is allelic to
GHLZ:
The similarities in the phe- hibited centromere linkage (data not shown). To testm2
mzm1 mz
0 0
CEN3
0 13HML
HIS4
LEU2
MAT
HMR
AlOGlO
"--I)---)-- U
0
A9D6
---€I0
A5F7
-
- -
U
0
A5D4
-"---", €I-
= 10 kb1504 K. S. Weiler, L. Szeto and J. R. Broach
l o 9 8 7 6 5 4 3 2 1 1 2 3 4 5 6 7 8
Ya
I
z1T T C A G C T T T C C G C A A ~ G T A i Wild Type
A A G T C G A A A G G C k G T T C A T
*
T T T A G C T T T C C G C A A ~ G T A
A A A T C G A A A G G C G T T C A T
dk
FIGURE 4.-Mutant A5E10 has a single base change in Ya of MAT. The DNA sequence of the Ya-Z1 junction region of A4ATa is compared for wild-type strain KSW3-20C and mutant strain A5E10. The position of the HO endonuclease cleavage site is illustrated. Strain A5E10 has a single base change of C
to T at position Y8.
whether ddpl-1 was an allele of CHLl, we crossed strain CTFl::TnlS, carrying a HIS3marked gene disruption of CHLl, to the ddpl-1 ho strain KSW30-C and tested the diploid for mating competence. The heteroallelic diploid exhibited weak a and a mating, indicating that the chl::Inl9HIS?and ddpl-1 mutations did not comple- ment (data not shown). In contrast, the two heterozy- gous mutant diploids produced by crossing each mu- tant to a wild-type strain of the opposing genetic background did not exhibit chromosome loss. To con- firm that ddpl-1 was an allele of CHLl, we crossed strain KSW30-C to strain CTFl-R:Tn43, which carries an in- sertion of HIS? into the adjacent, noncoding region of CHLl ( CHLl::In4?HIS?). The diploid was sporulated and all the His- progeny of each tetrad (of 10 exam- ined) were tested for the presence of the ddpl-1 allele as follows. Those His- progeny that carried the ho::URA? allele were incubated on 5-FOA medium to select for HO' recombinants having excised the URA? gene by intramolecular recombination. These 5-FOAR isolates were diploid due to HOmediated mating type interconversion and thus could be tested directly for the presence of ddpl-l as indicated by a and a mating ability. For those His- progeny that were Ura- and thus did not carry the ho::URA? allele, we tested for the presence of the ddpl-1 allele by complementation. These strains were crossed to either MATa strain KSW30-B or MATa strain KSW30-C, and the diploids were tested for weak mating ability. All 20 His- progeny of strain KSWJO-C/CTFl-R:Tn43 were found to have inherited the ddpl-1 allele. Therefore, ddpl-l is allelic to CHLl and will henceforth be referred to as the chll- 200 allele.
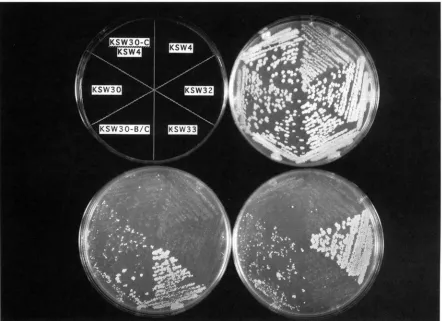
Disruption of chll abolishes donor preference in
MATa but not MATa cells: To determine the effect of
the chll-200 mutation on mating type interconversion, we subjected strain KSW30 to traditional pedigree anal- ysis, as described in MATERIALS AND METHODS. As shown
in Table 3, we found that the frequencies of switching from MATa to MATa and MATa to MATa were lower than those obtained for the wild-type strain, KSW4.
A
h m l d CEN3 MAT. HMRa h m l d
"3'
W k : L E U Z IfMRa-x-%&iEz
CENZ LYSZ4
Select His+ Lys' hndu.2 CEN3 MAT. HMRa CENZ LYSZhndu.2
'fgZ5
w k : L E U Z HMRa CENZ lysz::MATainrURA3-e
Screen His- Leu-h m l d CEN3 MAT. HMRa CENZ LYSZ
CENZ lysz::MATaineURA3
4
Select 5-FOARhndu.2 CEN3 HMRs CENZ LYSZ
P
CENZ LYSZ
B
1
Isolate1
a cells:
(# cells) (108) (91) (121)
FIGURE 5.-The ddpl-1 (chll-200) mutation of strain A3C10 is recessive to wild type. (A) The genetic scheme for complementation analysis is shown. Strains A3C10 and KSW3- 20C (represented on the left) were crossed to strain KSW29 (represented on the right). Only chromosomes ZZand ZIZare illustrated. Strain KSW29 carried a deletion of the MATlocus ( matA::LEU2), complemented by a MATcu,,,, allele at the lys2 locus, and a HZS-marked centromere mutation, cen3X35, that causes mitotic loss of chromosome ZZZ at a frequency of 2- 4% per generation ( GAUDET and FITZGERAI.D-HAYES 1987). One chromosome ZZZ monosomic isolate of diploid A3C10/ KSW29, detected as a His Leu colony, was retained as strain KSW39. As a control, the isogenic wild-type strain was con- structed in parallel from strains KSW3-20C and KSW29 to yield strain KSW41. Mating type interconversion was initiated in strains KSW39 and KSW41 by applying 5-FOA selection for gene conversion of lys2::MAT~u~,,~-UuRA3 to LYS2+, and confrontation analysis was performed on two 5-FOA resistant isolates from each aneuploid strain. (B) The results from confrontation analysis performed on two 5-FOA' isolates of each chromosome ZZZ monosomic strain (KSW39 and KsW41) are shown. One 5-FOAK isolate from strain KSW39 was retained as strain KSW40, and one 5-FOAR isolate from strain KSW41 was retained as strain KSW42.
However, the mutation appeared to have a slightly greater effect on a to a switching, that is, on selection of the HML locus. We cannot distinguish from these results whether the diminished switching efficiency ob- served for both cell types was a result of a decrease in the initiation of switching or from increased selection of the nonpreferred donor locus.
1505 Donor Preference Mutants
FIGURE 6.- d d j ~ l - l / ddj11-1 ( chll-200/ chll-200) diploids exhibit mating ability, which is enhanced in a /lo/ ho background. A standard mating assay was performed on the strains indicated in the upper left of the figure. Strains KSW32 and KSW33 were included as positive controls for haploid a and a mating, respectively. The remaining strains are all a / a diploid. The growth of the strains on YEPD before the assay is shown on the upper right. a mating is reflected in the growth of diploid colonies after mating with MATa tester strain DC14, shown in the plate on the lower left. a mating ability is reflected in the growth of diploid colonies after mating with MATa tester strain DC17, shown in the plate on the lower right.
we analyzed donor preference using a modified pedi- gree assay. This pedigree assay, described in detail in MATERIALS AND METHODS, has the advantage that selection of both the preferred and the nonpreferred donor could be monitored. In brief, the diploid
strains used for this analysis had alleles at H M L and
H M R that could be distinguished by size from each
other and from the allele initially present at M A T .
In addition, each donor locus allele carried an inc mutation to prevent further conversions of M A T . Ac- cordingly, the particular donor locus selected during
TABLE 3
The effect of chll-200 on mating type interconversion
MATa + MATa +
Strain Genotype MATa (%) MATa (%)
KSW4" CHLl/CHLl 88 (41)" 84 (86)
KSW30 ~hll-20O/~hll-200 58 (50) 69 (51)
Taken from WEILER and BROACH (1992).
"The number of cells examined for a mating type intercon- version event is indicated in parentheses.
the first switching event within a spore clone could be determined simply by performing PCR analysis on a sample of DNA from the colony.
We performed this PCR-based pedigree analysis on spores from strains homozygous for C H L l , for strains heterozygous for a URA3marked disruption of CHI-1
and for strains carrying homozygous disruptions of
C H L l . These results are presented in Table 4. Both
MATa CHLl and M A T a C H L l spores, derived either
from a strain homozygous for CHLJ o r from a strain heterozygous for c h l l , exhibited appropriate donor locus selection. The frequencies of correct donor lo- cus selection determined by this assay were essentially the same as those determined by the standard pedi- gree analysis based on pheromone challenge of indi- vidual cells at the four cell stage (cf. Table 3 ) . In contrast, MATa chll cells inappropriately selected
H M R as the donor locus almost as often as they se-
lected H M L . The same was not observed with M A T a
chll cells; they selected HMRwith the same efficiency
K. S. Weiler, L. Szeto and J. R. Broach 1506
TABLE 4
The effect of chllA on donor locus selection during mating type interconversion
Donor"
Strain Spore genotype HML HMR Ambiguous
LS204 MATaCHLl 76 (83%) 16 (17%) 0
MATaCHLl 6 (5%) 106 (95%) 0
LS342 MATaCHIl.1 32 (84%) 5 (13%) 1 (3%) MATaCHLl 0 42 (98%) 1 (2%) MATachll::URA3 21 (53%) 16 (40%) 3 (7%) MATachllr:URA3 4 (9%) 39 (89%) 1 (2%) LS349 MATachllAb 22 (63%) 13 (37%) 0
MATachllAb 1 (3%) 31 (97%) 0
The donor locus for the initial switch in the progeny of each spore was determined by PCR analysis of the MAT locus of the spore colony as described in MATERIALS AND METHODS. Number of spores using each donor are indicated with per- centages noted in parentheses. Ambiguous indicates that the PCR analysis did not yield a single predominant fragment. By
X' analysis, the difference between the results obtained with
MATaCHLl us. MATachll strains is statistically significant
( p
< 0.001), whereas the difference between the results obtained with MATaCHLl us. MATachll strains is not statistically sig- nificant.
"Allele disrupted by either URA3 or HIS?.
of the gene has an asymmetric effect on the selection process.
DISCUSSION
We report here the results of a novel mutant isolation scheme designed to identify components of the cellular machinery responsible for the selection of a donor lo- cus during mating type interconversion. We looked spe- cifically for mutants that frequently selected HMRa as a donor when cell type dictated HML as the preferred donor locus. Although some of the mutants obtained carried mutations at MAT a n d / o r HML, the mutant isolation scheme also yielded at least one trans-acting factor involved in mating type interconversion. As evi- denced by the mutant phenotypes of strains carrying the chll-200 ( d d p l - I ) mutation, the CHLl gene prod- uct is involved in selecting the appropriate donor locus. In addition, chromosomal rearrangements were ob- tained at high frequency from this screen, highlighting the fact that the integrity of chromosome IZZis crucial for efficient switching.
All translocation mutants belong to the class of muta-
tions that strongly impair the selection of the HML lo-
cus: The translocations present in mutants A5D4, A5F7, A9D6 and AlOGlO had the effect of reducing the frequency with which the HML locus was used as a donor locus. Given that the
ZZI;
VI1 or X V translocation (mutant A5D4) had a slightly more severe effect than the ZZI;V translocation (mutants A5F7, A9D6 and AlOGlO) , we wondered whether translocations involv- ing other chromosomes might differ with respect tothe subsequent level of participation of HML in the switching process. In other words, might the transloca- tion of HML to one chromosome handicap it only slightly while its translocation to another chromosome completely prevented it from being utilized as a donor locus? Two mutant strains that carried a copy of the HML locus translocated to either chromosome Z or chromosome X I 1 were examined by HABER et al. (1981 ) , but they retained the normal HML locus as well as the translocated copy and were thus not informa- tive with respect to this question. As one means to ask if chromosomal translocations might differ in phenotype, mutants with a very low percentage of a cells as well as mutants from the 100% a class were examined for the presence of translocations by PFGE. We found that none of the mutants examined had suffered a chromo- somal translocation ( data not shown)
.
The fact that four of seven mutants of strong but intermediate phe- notype carried a translocation while none of 19 weak mutants or 25 100% a mutants did so suggests that neither of these phenotypes are readily obtained by translocation. One interpretation of this result is that any translocation eventper
se severely inhibits, but does not completely abolish, the utilization of the transposed locus as a donor template during switching and that the new chromosomal environment of the translocated donor locus has only a limited influence. An alternative explanation is that different translocations could have markedly different effects on donor preference but that only a limited number of chromosome rearrangements are possible in our strain. For instance, if translocations resulted predominantly from recombination between Ty elements, as proposed above, the chromosomal dis- tribution of these retrotransposons would dictate the spectrum of translocations obtained.Allelespecific MAT sequence affects the frequency
of HO endonuclease cleavage: A single bp change at
Y8 of MATa resulted in the mutant phenotype exhibited by strain A5E10. The isolation of a mutation affecting HO cleavage of MATa in the allele-specific (Y) region was not entirely surprising, as a 24 bp sequence, includ- ing 11 bp of Y, was defined by NICKOLOFF et al. ( 1986) as the minimal sequence permitting H O cleavage in vivo. Moreover, mutational analyses of the H O endonu- clease recognition site by NICKOLOFF et al. (1990) indi- cated that the Y regions influenced HO endonuclease recognition. However, they found only one point muta- tion in Ya and one point mutation in Ycu, both at posi- tion Y4, that affected HO cleavage by their assay; other mutations resulting in decreased cleavage had 2 5 bp changes ( 2 4 bp changes within Y)
.
In fact, the Ya 8C to T mutation present in mutant A5E10 was tested in their assay and found to have no detectable effect. A likely explanation for this discrepancy could be the na- ture and sensitivity of the assay used by NICKOLOFF etDonor Preference Mutants 1507
soma1 ura3 gene, disrupted by an insertion of the syn- thetic
H O
cleavage site being examined, and a chromo- somal copy of ura3, as scored by the number of Ura+ papillae arising afterH O
endonuclease synthesis. The ura3 sequences surrounding the 3@bp synthetic cleav- age sites might have influenced recognition by the HO endonuclease, especially since the HO endonuclease footprint of MAT extends slightly beyond this 30-bp core sequence ( NICKOLOFF et al. 1990). Of perhaps greater significance, though, is the fact that synthesis of theH O
endonuclease was induced using a galactose- inducibleH O
gene for 24 hr before transferring the cells to glucose (repressive) medium. This is in stark contrast to the limited synthesis of this enzyme when transcription is controlled by theH O
gene promoter, as in our studies.The effect of d l 1 mutations on mating type intercon- version: Results of pedigree analyses of strains carrying deletions of chll demonstrated that, in the absence of
CHLl
function, MATa cells randomly select the donor locus during mating type interconversion, while MATa cells are unimpaired in their ability to select HMR pref- erentially. Slightly different results were obtained with strains carrying the chll-200 allele. While such strains showed a slightly greater defect for donor preference in a MATa background than in a MATa background, productive switching was diminished in both cell types. This difference in phenotype could reflect differences in the behavior of the two alleles. In fact, diploid strains homozygous for chll-200 exhibit poor spore viability, an effect that is not seen with strains homozygous for a c h l l A allele. Alternatively, the differences in donor preference phenotype may reflect differences in the two assays used to evaluate donor preference. That is, if, in addition to the donor preference defect, chll strains were slightly less efficient thanCHLl
strains in initiating mating type interconversion, such a defect would not be readily evident in the PCR-based assay. However, this defect would register in the traditional pedigree assay as a reduction in the percentage of pro- ductive switches at the four-cell stage. Thus, chll strains may be slightly defective in initiation of mating type interconversion (formation of the HO-mediated dou- ble strand break, for example) in addition to being clearly defective in donor selection.The CHLl gene product and donor locus selec- tion: Chllp is required for efficient chromosome segre- gation during mitosis ( LIRAS et al. 1978; SPENCER et al. 1990). While the original chll-1 mutation was reported to cause an increase in mitotic recombination (LIRAS et al. 1978), null mutations do not have a significant effect on recombination ( GERRING et al. 1990). Se- quence analysis of
CHLl
revealed its homology to RAD3 ( GERRING et al. 1990), which encodes an ATPdepen- dent helicase with DNA binding activity. Rad3p is a component of transcription factor b and is required for excision repair in yeast ( GUZDER et al. 1994; WANG etal. 1994). Mutational inactivation of the putative ATP- binding domain within
CHLl
abolishes its activity. Fi- nally, chll mutants have been shown to be synthetically lethal with kar3, which encodes a kinesin-like protein involved in mitosis and required for karyogamy ( MELUH and ROSE 1990; MELUH 1992),
and with mad2, required for a checkpoint mechanism apparently used for assessing formation of a functional mitotic spindle( LI and MURRAY 1991 )
.
We can speculate as to the role Chllp might play in the cell to account for the phenotypes of chll strains. LIRAS et al. (1978) suggested a connection between en- hanced recombination and the chromosome loss phe- notype of chll-1 strains. The fact that Chllp has helicase activity might suggest that it could be involved in resolu- tion of recombination intermediates such as cross- stranded structures. Failure to resolve recombination intermediates between sister chromatids in mitosis could account for the chromosome missegregation phe- notype. However, this explanation fails to account for loss of donor selectivity in MATa cells, an event that precedes initial formation of recombination intermedi- ates, or for the synthetic lethality of chll with kar3 or mad2.
A second possibility is that Chllp is directly involved in the mechanics of chromosome dynamics. The pro- tein could facilitate kinetochore assembly or function, connect DNA sequences to other supermolecular struc- tures in the nucleus or facilitate specific chromosome organization or assembly. A direct role in chromosome dynamics of Chllp would be consistent with its function- ing in the same process as KarSp, a putative microtu- bule-associated molecular motor, and for precluding defects surveyed by the mitosis checkpoint apparatus. In this context, the defect of chll in donor preference could be in assembly of the particular three-dimen- sional organization of chromosome IIIthat would facili- tate specific interactions of donor loci with MAT or in actual movement of the appropriate donor locus into spatial proximity of MAT.
A third possibility is that Chllp is required for proper assembly of the chromatin structure of some, if not all, chromosomes. Chromosome missegregation could be a consequence of an altered chromatin structure, which might impede the association of requisite factors with the centromeres to form a functional spindle. This would account for the G2/M delay seen with chll strains ( GERRING et al. 1990), as well as for the synthetic lethality of chll and mad2. The fact that mutations of
SZiVl, which encodes a protein homologous to HMGl,