Next-Generation Antimicrobial Susceptibility Testing
Alex van Belkum,aW. Michael Dunne, Jr.b
bioMérieux SA, Microbiology Unit, R&D Microbiology, La Balme Les Grottes, Francea
; bioMérieux, Inc., Microbiology Unit, R&D Microbiology, Durham, North Carolina, USAb
Antimicrobial resistance has emerged as one of the most-significant health care problems of the new millennium, and the clini-cal microbiology laboratory plays a central role in optimizing the therapeutic management of patients with infection. This mini-review explores the potential value of innovative methods for antimicrobial susceptibility testing of microorganisms that could provide valuable alternatives to existing methodologies in the very near future.
TODAY’S GLOBAL AST LANDSCAPE
A
limited number of methods for antimicrobial susceptibilitytesting (AST) of medically important microorganisms have survived the maturation of modern diagnostic clinical microbiol-ogy. Surprisingly, one of these is the disk diffusion method first
published in 1966 (1) and the various iterations thereof. Another
is broth microdilution (BMD) testing, which has attained refer-ence standard status to which all other AST methods are currently compared during development, verification, validation, and clin-ical trials. As such, BMD displaced agar dilution testing, the past gold standard methodology.
The most important outcome of any AST is the rapid and re-liable prediction of antimicrobial success in the treatment of in-fection. Currently, AST is typically accomplished using either clas-sical manual methods or growth-dependent automated systems, such as the Becton, Dickinson Phoenix, the Siemens Micoscan WalkAway, or the bioMérieux Vitek 2, all of which are based on BMD testing. The major limitations of these methods include the requirement for relatively large numbers of viable organisms, complicated preanalytical processing, limited organism spectrum,
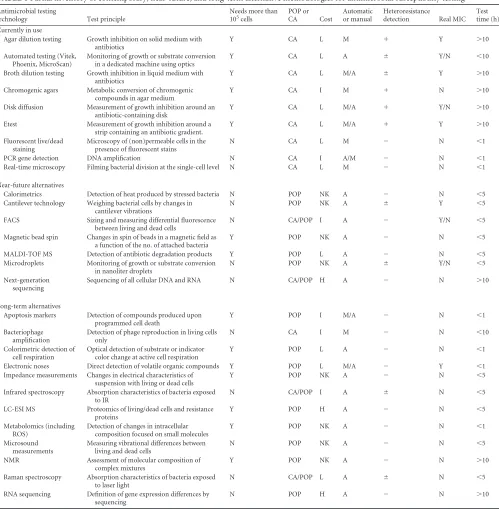
analytical variability, time to results, and cost.Table 1provides a
cursory review of current and future technologies and their strengths and weaknesses.
At present, nonphenotypic, mostly nucleic acid-based AST methods cannot detect all resistance markers, are expensive, and have not been widely adopted. Multiplex PCR detection of resis-tance determinants directly from positive blood cultures, how-ever, has been shown to substantially reduce the time to clinically
actionable results (2). Furthermore, digital PCR may allow for
better quantification of target molecules present in starting
mate-rial (3), and the refinement of aptamer technology
(single-stranded short RNA or DNA molecules with antibody-like
prop-erties) may further facilitate nucleic acid diagnostics (4). Clearly,
newer-generation, transcriptome, and whole-genome sequencing will provide near-future options to resistance prediction as data-bases mature. The per-strain assessment of detailed MICs for all relevant antimicrobials may be confounded by elevated degrees of genetic heterogeneity, as is, for instance, obvious among many Gram-negative bacterial species. This may still frustrate the genomic approach, but this would be the subject of an entire re-view by itself and will not be discussed here.
Novel options are available to supplant the existing toolbox, but the timing of such events is hard to foresee. As a disclaimer, we cannot provide a complete survey of all potential AST configura-tions but will try to highlight a number of phenotypic methods
that provide insight into the wealth of the possibilities to come. Below, we will briefly describe six technologies that could repre-sent competition for current reference standard methods— nucleic acid amplification and sequencing excluded.
NEAR-FUTURE ALTERNATIVES FOR ROUTINE AST
MS. Matrix-assisted laser desorption ionization–time of flight
mass spectrometry (MALDI-TOF MS), a powerful tool for the rapid identification of organisms with medical importance, may
also prove to be of value as an AST method (5). Several approaches
have been explored, including (i) documenting the activity of
an-tibiotic-inactivating enzymes (e.g.,-lactamases), (ii) confirming
the presence of a PCR product indicative of antimicrobial
resis-tance (e.g.,vanA,mecA, or NDM-1), and (iii) observing changes
in the protein spectrum of an organism in the presence or absence of an antimicrobial agent that correlate with susceptibility changes.
As an example of the first, carbapenemase activity was detected in a variety of Gram-negative organisms using ertapenem as a
substrate (6). A bacterial suspension was incubated with
ertap-enem, and serial samples were examined by MALDI-TOF MS. Organisms producing either NDM-1 or IMP-1 completely hydro-lyzed ertapenem within 1 h. It should be noted that four distinct peaks were initially observed in the mass spectrum of the parent drug, which already included the inactive hydrolyzed form. Other carbapenemases (IMP-2, VIM-1, VIM-2, and KPC-2) hydrolyzed the drug more slowly. Similar data were generated using mero-penem as a substrate for enzymes including NDM-1, VIM-1,
KPC-2, KPC-3, OXA-48, and OXA-162 (7). The applicability of
this method using other substrates (penicillin G, ampicillin, cefoxitin, and imipenem) was demonstrated with and without
clavulanic acid as a means of distinguishing-lactamase classes
such as AmpC or TEM-1 (8). The possibility exists for
multiplex-ing the assay usmultiplex-ing combinations of-lactams and inhibitors,
which would allow classification of extended-spectrum
-lacta-mases as well.
The second approach was presented in a study that compared two methods of single-nucleotide polymorphism (SNP) analysis for epidemiological typing of 147 strains of methicillin-resistant
Published ahead of print13 March 2013
Address correspondence to Alex van Belkum, [email protected]. Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JCM.00313-13
on May 16, 2020 by guest
http://jcm.asm.org/
Staphylococcus aureus(MRSA) at 16 distinct loci (9). The predi-cate method of analysis was a real-time SYBR green PCR assay. The comparator method employed the Sequenom MassARRAY iPLEX SNP typing platform (Sequenom, Brisbane, Australia), which combines multiplexed single-base extension PCR with MALDI-TOF MS of amplicons to determine the location of SNPs.
Both methods proved comparable, and themecAPCR amplicon
was successfully identified by MALDI-TOF MS for all 147 strains.
A combination of a primer extension (PEX) reaction with MALDI-TOF MS also led to the detection of ganciclovir resistance mutations in cytomegalovirus (CMV) among viremic heart
trans-plant patients (10). Compared to a combination of real-time PCR
[image:2.585.44.543.73.582.2]and Sanger sequencing, the PEX/MALDI-TOF MS method dis-closed resistance mutations earlier without loss of specificity. Sim-ilar analyses of PCR-generated amplicons are the basis for resis-tance detection using the more-advanced electrospray ionization
TABLE 1Partial inventory of contemporary, near-future, and long-term alternative methodologies for antimicrobial susceptibility testinga
Antimicrobial testing
technology Test principle
Needs more than 105
cells
POP or CA Cost
Automatic or manual
Heteroresistance
detection Real MIC Test time (h) Currently in use
Agar dilution testing Growth inhibition on solid medium with antibiotics
Y CA L M ⫹ Y ⬎10
Automated testing (Vitek, Phoenix, MicroScan)
Monitoring of growth or substrate conversion in a dedicated machine using optics
Y CA L A ⫾ Y/N ⬍10
Broth dilution testing Growth inhibition in liquid medium with antibiotics
Y CA L M/A ⫾ Y ⬎10
Chromogenic agars Metabolic conversion of chromogenic compounds in agar medium
Y CA I M ⫹ N ⬎10
Disk diffusion Measurement of growth inhibition around an antibiotic-containing disk
Y CA L M/A ⫹ Y/N ⬎10
Etest Measurement of growth inhibition around a strip containing an antibiotic gradient.
Y CA L M/A ⫹ Y ⬎10
Fluorescent live/dead staining
Microscopy of (non)permeable cells in the presence of fluorescent stains
N CA L M ⫺ N ⬍1
PCR gene detection DNA amplification N CA I A/M ⫺ N ⬍1 Real-time microscopy Filming bacterial division at the single-cell level N CA L M ⫺ N ⬍1
Near-future alternatives
Calorimetrics Detection of heat produced by stressed bacteria N POP NK A ⫺ N ⬍5 Cantilever technology Weighing bacterial cells by changes in
cantilever vibrations
N POP NK A ⫾ Y ⬍5
FACS Sizing and measuring differential fluorescence between living and dead cells
N CA/POP I A ⫺ Y/N ⬍5
Magnetic bead spin Changes in spin of beads in a magnetic field as a function of the no. of attached bacteria
Y POP NK A ⫺ N ⬍5
MALDI-TOF MS Detection of antibiotic degradation products Y POP L A ⫺ N ⬍5 Microdroplets Monitoring of growth or substrate conversion
in nanoliter droplets
N POP NK A ⫾ Y/N ⬍5
Next-generation sequencing
Sequencing of all cellular DNA and RNA N CA/POP H A ⫺ N ⬎10
Long-term alternatives
Apoptosis markers Detection of compounds produced upon programmed cell death
Y POP I M/A ⫺ N ⬍1
Bacteriophage amplification
Detection of phage reproduction in living cells only
N CA I M ⫺ N ⬍10
Colorimetric detection of cell respiration
Optical detection of substrate or indicator color change at active cell respiration
Y POP L A ⫺ N ⬍1
Electronic noses Direct detection of volatile organic compounds Y POP L M/A ⫺ Y ⬍1 Impedance measurements Changes in electrical characteristics of
suspension with living or dead cells
Y POP NK A ⫺ N ⬍5
Infrared spectroscopy Absorption characteristics of bacteria exposed to IR
N CA/POP I A ⫾ N ⬍5
LC-ESI MS Proteomics of living/dead cells and resistance proteins
Y POP H A ⫺ N ⬍5
Metabolomics (including ROS)
Detection of changes in intracellular composition focused on small molecules
Y POP NK A ⫺ N ⬍1
Microsound measurements
Measuring vibrational differences between living and dead cells
N POP NK A ⫺ N ⬍5
NMR Assessment of molecular composition of complex mixtures
Y POP NK A ⫺ N ⬎10
Raman spectroscopy Absorption characteristics of bacteria exposed to laser light
N CA/POP L A ⫾ N ⬍5
RNA sequencing Definition of gene expression differences by sequencing
N POP H A ⫺ N ⬎10
a
FACS, fluorescence-activated cell sorting; MALDI-TOF MS, matrix-assisted laser desorption ionization–time of flight mass spectrometry; LC-ESI MS, liquid chromatography-electron spray ionization mass spectrometry; ROS, reactive oxygen species; NMR, nuclear magnetic resonance; Y, yes; N, no; POP, proof of principle; CA, commercially available; H, high; L, low; I, intermediate; NK, not known;⫹, detects heteroresistance;⫾, may detect heteroresistance;⫺, fails to detect heteroresistance.
on May 16, 2020 by guest
http://jcm.asm.org/
mass spectrometry (PCR/ESI-MS). In one study, the quinolone
resistance-determining regions ofparCandgyrAof
multidrug-resistant strains ofAcinetobacter spp. were identified with
ade-quate correlation to BMD testing (11).
Finally, there are a number of examples of MALDI-TOF MS being used to highlight the effects of antimicrobial agents on the protein spectral profile of susceptible organisms. Comparisons of
the profiles ofCandida albicansgrown in the presence of
increas-ing concentrations of fluconazole led to the formulation of a min-imal profile change concentration (MPCC) that was defined as the lowest concentration of the drug at which a change in the profile
could be documented (12). The authors found a very high
con-cordance between the MPCC and the MIC values obtained by the CLSI broth-based reference method. Similarly, MALDI-TOF MS
was used to assess caspofungin resistance secondary tofks
muta-tions in 34Candidaspecies and 10Aspergillusisolates (13). Strains
were exposed to increasing concentrations of caspofungin in a BMD format, along with a drug-free control well, and incubated for 15 h prior to MALDI-TOF MS. For each drug concentration, an MPCC was calculated for each strain. This group found 100% essential agreement for all of the isolates using CLSI breakpoints for MIC or minimal effective concentration (MEC). Only two
Candidaisolates were incorrectly interpreted as nonsusceptible, generating a categorical agreement of 94.1%.
FC.Flow cytometry (FC) permits changes in the morphology,
physiological and metabolic activity, and viability of microorgan-isms to be followed after exposure to antibiotics. Through a pro-cess of staining with nucleic acid dyes that do not permeate the cell walls of healthy organisms, the proportion of cells in a dying or dead state (and everything in between) can be rapidly assessed by examining emission spectra after the cells pass individually
through a flow channel and when the dye is excited by a laser (14).
In early studies, a primitive flow cytometer constructed from a fluorescence microscope was used to assess cellular morphology or DNA after bacteria were exposed to antimicrobial agents. It was concluded that the effects of antimicrobial treatment could be detected within a few hours, suggesting a promising application for AST. The earlier studies were also useful for elucidating vari-ous dye/fixation combinations that would better differentiate the
state of cell viability by FC (15). In 1997, propidium iodide was
used to differentiate live/dead C. albicans cells treated with
amphotericin B or fluconazole, which could then be rapidly and
sensitively quantified by FC (16). Similar assays were developed
for the echinocandins, caspofungin, and additional azoles (17).
The results of these AST strategies were 96 to 99% concordant
with other forms of AST (18). The overall duration of AST could
be reduced from overnight incubations to 1 to 2 h. Also in 1997, bis(1,3-dibutylbarbituric acid) trimethine oxonol (DiBAC4[3]) was used to visualize anionic membrane potential changes using
fluorescence, which proved to be very adequate for AST of
Esche-richia coli(19). Tests of organisms causing urinary tract infection showed 94% agreement between classical disk diffusion testing
and DiBAC4[3]-FC testing (20). FC AST was also described for
Mycobacterium tuberculosis. Pyrizinamide susceptibility testing by FC was 93% concordant with the Bactec MGIT assay, and the
former was conclusive within 24 h (21). ForYersiniaspp. also,
significantly faster testing was achieved using FC (22). FC AST was
at least 20% faster than classical methods forE. coli,Pseudomonas
aeruginosa, andS. aureus, and extended-spectrum-lactamases
could be reliably detected by FC in 1 to 2 h (23). Although
signif-icant advances in the design and performance of FC instrumenta-tion have occurred, the technology has not yet emerged as a major player in the AST market, although commercial assays have been launched. Some of the perceived hurdles for the methodology are, among others, the ability to differentiate cellular damage caused by cidal versus static antibiotics, autofluorescence of certain bac-terial species, and the tremendous amount of work required for verification/validation of the clinical database and the method it-self.
Microbial cell weighing by vibrating cantilevers.Cantilevers containing small canals which facilitate microbial passage can be made to vibrate continuously. When bacteria pass through, their weight (in the femtogram range) will cause a change in the
fre-quency of cantilever movement (24). Less-dense cells will cause a
different change than more-dense cells. When cells are treated with antimicrobial agents, their buoyant mass density changes,
and this is measurable (25). The principle has been proven using
ampicillin-resistant and -susceptible variants ofCitrobacter
roden-tium. It was also shown that resuscitation of both phenotypes after
osmotic shock in the presence or absence of ampicillin allows rapid differentiation in a reduced time span. Cantilevers can be multiplexed using nanotechnology such that multiple antibiotics in various concentrations could be tested for a single growing culture simultaneously.
A rapid biosensor for the detection of bacterial growth was developed using vibrating cantilevers containing a certain number
of fixed but still viable bacteria (26). The change in resonance
frequency as a function of the increasing mass on the cantilever forms the basis of the detection scheme. The calculated mass sen-sitivity according to the mechanical properties of the cantilever sensor is approximately 50 pg/Hz; this mass corresponds to about 100E. colicells. The sensor was able to detect active growth ofE.
colicells within 1 h. The number ofE. colicells initially attached to
the cantilever was on average 1,000 cells. Furthermore, the non-inhibited growth of resistant cells could be documented within 2 h
after the addition of antibiotics (27). Cantilever technology has
also been used to assess vancomycin binding to cell wall
precur-sors (28) and to measure the effects of colistin onP. aeruginosa
(29).
Using suspended nanochannel resonators (SNRs), it was dem-onstrated that the measurement of bacterial mass in solution was even more precise. The SNR consisted of a cantilever with an embedded nanochannel. In addition, a new method was intro-duced that uses centrifugal force caused by vibration of the
canti-lever to trap particles at the free end of the SNR (30). This
ap-proach eliminates the intrinsic position-dependent error of the SNR and also improves the mass resolution by increasing the av-erage “time of presence” for each particle. In addition, it would facilitate the continuous mass monitoring of a limited number of bacteria during (changing) exposure to antibiotics. Clearly, can-tilever systems and precise weight measurements provide an in-teresting option for the development of multiplexed AST.
IMC.Isothermal microcalorimetry (IMC) is a dynamic
tech-nique that allows the measurement of heat production either as a
flow rate (W/unit time) or as total accumulation over time
(Joules/unit time) stemming from the metabolism of actively growing cells. Cumulative heat production generally parallels conventional growth curves in that the slope and shape of the accumulating heat production correspond with classical lag, log, and stationary phases. Maximum heat values represent the total
on May 16, 2020 by guest
http://jcm.asm.org/
number of cells produced over time (31). The method has been successfully adapted to small culture volumes (e.g., 1 to 3 ml) and can be used in conjunction with either solid or liquid culture
me-dium (32). Using IMC, bacterial species identification from urine
specimens was performed even at low bacterial counts within 3 h
on the basis of dynamic heat flow patterns (33). When adapted to
AST, measurements of heat production are made passively from sealed vials containing the organism, growth medium, and anti-microbial agents in doubling-dilution concentrations. The mini-mal heat inhibition concentration (MHIC) can be defined as the lowest drug concentration to either inhibit 50% of total heat pro-duction or result in a 50% repro-duction in heat flow rate, depending on the drug being tested. The process requires specific IMC instru-mentation (e.g., TAM III; TA Instruments, New Castle, DE) for real-time measurement of heat generation with a detection limit
in the 0.2-W range.
The use of IMC for susceptibility testing is not new but has
been successfully adapted for AST of bacteria (31), mycobacteria,
includingM. tuberculosis(32), and fungi (34,35). The advantages
of this technique include the following: (i) testing is conducted in sealed ampules, alleviating safety concerns when evaluating
high-risk organisms, such asM. tuberculosisor fungal species; (ii) all
monitoring during testing is passive and requires no manual ma-nipulation of the test vials; and (iii) the completed analysis pro-vides information about the maximum growth rate of the organ-ism, static versus cidal activity of an agent, and delays to log-phase growth (extended lag phase) caused by the agent. The latter pro-vides useful information concerning antimicrobial activity at sub-inhibitory concentrations of the drug and can help predict the actual MIC when inhibitory concentrations have not been
devel-oped (31). This is pronounced with fungal testing, where delays or
an abbreviation of maximum heat flow can be readily appreciated
with subinhibitory concentrations of antifungal agents (34,35).
Furthermore, IMC is not prone to subjective interpretations, such as trailing MBD wells or the determination of MECs based on morphological changes at a microscopic level. IMC correlates well with BMD testing and CLSI and/or EUCAST breakpoints when net heat production over time is used as a surrogate for growth. As a bonus, IMC can be used to evaluate the synergistic activity of antimicrobial combinations. Chip calorimetry is a monitoring tool for determining the physiological state of biofilms. Its poten-tial use for the study of the effects of antibiotics was tested using an established model. The real-time monitoring potential of chip cal-orimetry was successfully demonstrated: a dosage of antibiotics initially increased the heat production rate, probably due to
activ-ity of energy-dependent resistance mechanisms (36). The
subse-quent reduction in heat production was probably due to the loss of activity and the death of the biofilm. This new analytical tool pro-vided fast, quantitative, and mechanistic insights into the effects of antibiotics on biofilm activity. In short, the maximum bacterial growth rate and the start of the lag phase can be quantified by microcalorimetric technology in an affordable and sensitive man-ner. Hence, antibiotic-associated changes in these parameters can be efficiently measured as well.
Magnetic bead rotation.When magnetic beads are brought into a revolving magnetic field, they self-assemble and assume a specific rotational spin. The frequency of rotation can be influ-enced by the binding of molecules, viruses, or bacteria. So, if the beads are equipped with a ligand that specifically captures bacte-rial cells, the rotation of the beads changes at the moment of
cap-ture. This change can be measured. If all beads in a broth culture are paired with one or two cells, something that can be accom-plished by incubating ligand-modified beads with a diluted bacte-rial suspension followed by washing, they will resume a constant rotational frequency. As bacteria start to divide, the rotation fre-quency changes. If cell division is inhibited or blocked by antimi-crobials due to susceptibility, the change is arrested. If the bacteria are resistant to the antimicrobial applied, again, a change in rota-tional frequency occurs. In this way, antimicrobial resistance can be detected and precisely quantified.
A growth-based antimicrobial susceptibility assay based on asynchronous magnetic bead rotation (AMBR) biosensors has
been described (37). In this system, the effects of bacterial growth
on the rotation and shape of a cluster of self-assembling magnetic microbeads in a rotating magnetic field can be observed over time. The rotational period (RP) is indirectly proportional to the drag coefficient of the surrounding medium containing bacteria, broth medium, and antimicrobial agents at various dilutions. The RP increases as organisms multiply and attach to the organism-spe-cific antibody-coated beads or if the viscosity of the growth me-dium is altered. The addition of antimicrobial agents in increasing concentrations prevents an increase in RP over time. This process can be observed directly by illuminating the culture broth (in the form of a hanging drop) with a light-emitting diode (LED) or laser. The hanging-drop format also acts as a lens to create
mag-nification of up to 100⫻such that the structure and rotational rate
of the bead aggregates can be observed microscopically or when projected onto a detector. Serial 2-fold dilutions of streptomycin and gentamicin were added to Mueller-Hinton broth containing antibody-sensitized magnetic beads prebound to a standardized
inoculum ofE. coli. A hanging drop was formed with the mixture
that was subjected to an oscillating magnetic field. Changes in the RP were observed microscopically over time and recorded. As expected, the RP increased relative to bacterial growth, with solu-tions containing higher concentrasolu-tions of antibiotic demonstrat-ing the lowest increases. This method can be miniaturized to nanoliter-volume water-in-oil droplets containing 50 or fewer
bacterial cells per droplet (38). These changes substantially
re-duced the duration of a test.
Testing in microdroplets.Micro- or nanodroplets can be used as small individual reaction wells. The droplets can be individually manipulated, and when they contain bacteria in sufficient numbers, the metabolic activity and viability of cells can be monitored. The development of this system became feasible once the emulsification process was refined and the
long-term stability of the droplets could be guaranteed (39). A
system consisting of 100-nl droplets containing 103 bacteria
per droplet and differing concentrations of antibiotics was
de-veloped (40). By following the droplets over time using
epifluo-rescence, growth curves can be monitored at each drug concen-tration. More recently, droplets were prepared that contain a
single bacterial cell (41). This technology can be miniaturized
and easily multiplexed with respect to the number of antibiot-ics tested per organism The duration of testing can be as short as a single or a few bacterial replication cycles. Obviously, the assessment of technical reproducibility and development of adequate reference MIC databases will be necessary before this approach can be evaluated as a routine AST tool.
on May 16, 2020 by guest
http://jcm.asm.org/
TECHNOLOGIES FOR APPLICATION IN THE MORE-DISTANT FUTURE
Several innovative AST methods have been designed and
pre-sented over the past decade (Table 1, Long-term alternatives), but
most are far from introduction into routine clinical microbiology. Some, including bacteriophage-based AST, may be closer than the others. The bacteriophage-based strategy has been successfully
adapted for testing of M. tuberculosis (42), with optimization
achieved by including recombinant phages containing luciferase
genes (43). It was shown that the luciferase assay could be
per-formed at low cost in 2 days (44) and could be applied successfully
in laboratories in developing countries (45). Unfortunately, this
innovative approach has not been broadly accepted in diagnostic
microbiology laboratories, probably due to contamination (46) or
phage resistance issues. As stated above, as promising as some innovative methods may seem to be, broad acceptance has not yet been achieved.
Real-time microscopy is one of the innovative technologies that may be applicable to AST in the not-too-distant future. High-resolution camera-based systems have been commercialized, and
these show a high degree of efficacy (47). This technology has been
extended for use with microcolonies and serial photography in the presence or absence of antibiotics. Such technologies can generate a “1-day AST” for bacteria from positive blood culture bottles
(48). This demonstrates that simple, seemingly old-fashioned test
formats can still be adapted into systems that better serve the pa-tient’s needs.
Several modern technologies have been proposed as being pos-sible long-term future alternatives to today’s technologies in the
clinical microbiology laboratory; these are briefly described in
Ta-ble 1. Proof of principle has been demonstrated, and in some cases, the first trials of feasibility have been published. However, for all of these strategies, database development has yet to be com-pleted, and until then, the prospects of these technologies remain nebulous.
CONCLUSIONS
Conventional AST has its diagnostic limitations: it is generally time-consuming, and actionable results have a tendency for late arrival. The current methods are very solid and well-respected and do generally have CE and FDA certification. Any new technology has to compete with current reference standards, and the method that shows significant improvements has yet to be published. As we described here, several technologies may be knocking on the door shortly.
ACKNOWLEDGMENTS
The authors are employees of bioMérieux, a company creating and devel-oping infectious disease diagnostics. No further potential conflicts of in-terest relevant to this article are reported.
The classification of the various innovative AST systems in the three categories suggested inTable 1represents the authors’ personal opinions. REFERENCES
1.Bauer AW, Kirby WMM, Sherris JC, Turck M.1966. Antibiotic suscep-tibility testing by a standardized single disc method. Am. J. Clin. Pathol. 45:493– 496.
2.Waldeisen JR, Wang T, Mitra D, Lee LP.2011. A real-time PCR antibi-ogram for drug-resistant sepsis. PLoS One6:e28528. doi:10.1371/journal .pone.0028528.
3.Hatch AC, Fisher JS, Tovar AR, Hsieh AT, Lin R, Pentoney SL, Yang
DL, Lee AP.2011. 1-Million droplet array with wide-field fluorescence imaging for digital PCR. Lab Chip11:3838 –3845.
4.DeGrasse JA.2012. A single-stranded DNA aptamer that selectively binds toStaphylococcus aureusenterotoxin B. PLoS One7:e33410. doi:10.1371 /journal.pone.0033410.
5.van Belkum A, Welker M, Erhard M, Chatellier S.2012. Biomedical mass spectrometry in today’s and tomorrow’s clinical microbiology labo-ratories. J. Clin. Microbiol.50:1513–1517.
6.Burckhardt I, Zimmermann S.2011. Using matrix-assisted laser desorp-tion ionizadesorp-tion–time of flight mass spectrometry to detect carbapenem resistance within 1 to 2.5 hours. J. Clin. Microbiol.49:3321–3324. 7.Hrabák J, Walková R, Studentová V, Chudácková E, Bergerová T.2011.
Carbapenemase activity detection by matrix-assisted laser desorption ion-ization–time of flight mass spectrometry. J. Clin. Microbiol.49:3222– 3227.
8.Hooff GP, van Kampen JJA, Meesters RJW, van Belkum A, Goessens WHF, Luider TM.2012. Characterization of-lactamase enzyme activity in bacterial lysates using MALDI-mass spectrometry. J. Proteome Res. 11:79 – 84.
9.Syrmis MW, Moser RJ, Whiley DM, Vaska V, Coombs GW, Nissen MD, Sloots TP, Nimmo GR. 2011. Comparison of a multiplexed MassARRAY system with real-time allele-specific PCR technology for genotyping of methicillin-resistantStaphylococcus aureus. Clin. Micro-biol. Infect.17:1804 –1810.
10. Zürcher S, Mooser C, Lüthi AU, Mühlemann K, Barbani MT, Mohacsi P, Garzoni C, Gorgievski-Hrisoho M, Schaller A, Flatz L.2012. Sensitive and rapid detection of ganciclovir resistance by PCR based MALDI-TOF analysis. J. Clin. Virol.54:359 –363.
11. Hujer KM, Jujer AM, Endimiani A, Thomson JM, Adams MD, Goglin K, Eshoo PN, Sampath R, Blyn LB, Ecker DJ, Bonomo RA.2009. Rapid determination of quinolone resistance inAcinetobacterspp. J. Clin. Mi-crobiol.47:1436 –1442.
12. Marinach C, Alanio A, Palous M, Kwesek S, Fekkar A, Brossas J-Y, Brun S, Snounou G, Hennequin C, Sanglard D, Datry A, Golmard J-L, Mazier D.2009. MALDI-TOF MS-based drug susceptibility testing of pathogens: The example ofCandida albicansand fluconazole. Proteomics 9:4627– 4631.
13. De Carolis E, Vella A, Florio AR, Posteraro P, Perlin DS, Sanuinetti M, Posteraro B.2012. Use of matrix-assisted laser desorption ionization– time of flight mass spectrometry for caspofungin susceptibility testing of
CandidaandAspergillusspecies. J. Clin. Microbiol.50:2479 –2483. 14. Soejima T, Minami J, Iwatsuki K.2012. The exclusive use of flow
cytom-etry to evaluate the antibiotic-susceptibility. Biochim. Biophys. Acta1820: 1980 –1986.
15. Alvarez-Barrientos A, Arroyo J, Cantón R, Nombela C, Sánchez-Pérez M.2000. Applications of flow cytometry to clinical microbiology. Clin. Microbiol. Rev.13:167–195.
16. Ramani R, Ramani A, Wong SJ.1997. Rapid flow cytometric suscepti-bility testing ofCandida albicans. J. Clin. Microbiol.35:2320 –2324. 17. Rudensky B, Broidie E, Yinnon AM, Weitzman T, Paz E, Keller N,
Raveh D.2005. Rapid flow-cytometric susceptibility testing ofCandida
species. J. Antimicrob. Chemother.5:106 –109.
18. Chaturvedi V, Ramani R, Pfaller MA.2004. Collaborative study of the NCCLS and flow cytometry methods for antifungal susceptibility testing ofCandida albicans. J. Clin. Microbiol.42:2249 –2251.
19. Jepras RI, Paul FE, Pearson SC, Wilkinson MJ.1997. Rapid assessment of antibiotic effects onEscherichia coliby bis-(1,3-dibutylbarbituric acid) trimethine oxonol and flow cytometry. Antimicrob. Agents Chemother. 41:2001–2005.
20. Gauthier C, St-Pierre Y, Villemur R.2002. Rapid antimicrobial suscep-tibility testing of urinary tract isolates and samples by flow cytometry. J. Med. Microbiol.51:192–200.
21. Fredricks BA, DeCoster DJ, Kim Y, Sparks N, Callister SM, Schell RF. 2006. Rapid pyrazinamide susceptibility testing ofMycobacterium tuber-culosisby flow cytometry. J. Microbiol. Methods67:266 –272.
22. Steinberger-Levy I, Zahavy E, Cohen S, Flashner Y, Mamroud E, Aftalion M, Gur D, Ber R.2007. Enrichment ofYersinia pestisfrom blood cultures enables rapid antimicrobial susceptibility determination by flow cytometry. Adv. Exp. Med. Biol.603:339 –350.
23. Faria-Ramos I, Espinar MJ, Rocha R, Santos-Antunes J, Rodrigues AG, Cantón R, Pina-Vaz C.2013. A novel flow cytometric assay for rapid detection of extended-spectrum beta-lactamases. Clin. Microbiol. Infect. 19:E8 –E15.
on May 16, 2020 by guest
http://jcm.asm.org/
24. Godin M, Delgado FF, Son S, Grover WH, Bryan AK, Tzur A, Jor-gensen P, Payer K, Grossman AD, Kirschner MW, Manalis SR.2010. Using buoyant mass to measure the growth of single cells. Nat. Methods 7:387–390.
25. Knudsen SM, von Muhlen MG, Schauer DB, Manalis SR.2009. Deter-mination of bacterial antibiotic resistance based on osmotic shock re-sponse. Anal. Chem.81:7087–7090.
26. Gfeller KY, Nugaeva N, Hegner M.2005. Rapid biosensor for detection of antibiotic-selective growth ofEscherichia coli. Appl. Environ. Micro-biol.71:2626 –2631.
27. Gfeller, KY, Nugaeva N, Hegner M.2005. Micromechanical oscillators as rapid biosensor for the detection of active growth ofEscherichia coli. Bio-sens. Bioelectron.21:528 –533.
28. Ndieyira JW, Watari M, Barrera AD, Zhou D, Vögtli M, Batchelor M, Cooper MA, Strunz T, Horton MA, Abell C, Rayment T, Aeppli G, McKendry RA. 2008. Nanomechanical detection of antibiotic-mucopeptide binding in a model for superbug drug resistance. Nat. Nano-technol.3:691– 696.
29. Mortensen NP, Fowlkes JD, Sullivan CJ, Allison DP, Larsen NB, Molin S, Doktycz MJ.2009. Effects of colistin on surface ultrastructure and nanomechanics ofPseudomonas aeruginosacells. Langmuir25:3728 – 3733.
30. Lee J, Shen W, Payer K, Burg TP, Manalis SR.2010. Toward attogram mass measurements in solution with suspended nanochannel resonators. Nano Lett.10:2537–2542.
31. von Ah U, Wirz D, Daniels AU.2009. Isothermal micro calorimetry—a new method for MIC determinations: results for 12 antibiotics and refer-ence strains ofEscherichia coliandStaphylococcus aureus. BMC Microbiol. 9:106. doi:10.1186/1471-2180-9-106.
32. Howell M, Wirz D, Daniels AU, Braissant O.2012. Application of a microcalorimetric method for determining drug susceptibility in Myco-bacteriumspecies. J. Clin. Microbiol.50:16 –20.
33. Bonkat G, Braissant O, Widmer AF, Frei R, Rieken M, Wyler S, Gasser TC, Wirz D, Daniels AU, Bachmann A.2012. Rapid detection of urinary tract pathogens using microcalorimetry: principle, technique and first re-sults. BJU Int.110:892– 897.
34. Furustrand Tafin U, Meis JF, Trampuz AJ.2012. Isothermal microcalo-rimetry for antifungal susceptibility testing ofMucorales,Fusariumspp., andScedosporiumspp. Diagn. Microbiol. Infect. Dis.73:330 –337. 35. Furustrand Tafin U, Clauss M, Hauser PM, Bille J, Meis JF, Trampuz
A.2012. Isothermal microcalorimetry: a novel method for real-time de-termination of antifungal susceptibility ofAspergillusspecies. Clin. Micro-biol. Infect.18:e241– e245. doi:10.1111/j.1469-0691.2012.03854.x. 36. Buchholz F, Wolf A, Lerchner J, Mertens F, Harms H, Maskow T.2010.
Chip calorimetry for fast and reliable evaluation of bactericidal and bac-teriostatic treatments of biofilms. Antimicrob. Agents Chemother.54: 312–319.
37. Kinnunen P, McNaughton BH, Albertson T, Sinn I, Mofakham S, Elbez R, Newton DW, Hunt A, Kopleman R.2012. Self-assembled magnetic bead biosensor for measuring bacterial growth and antimicrobial suscep-tibility testing. Small8:2477–2482.
38. Sinn I, Albertson T, Kinnunen P, Breslauer DN, McNaughton BH, Burns MA, Kopelman R.2012. Asynchronous magnetic bead rotation microviscometer for rapid, sensitive, and label-free studies of bacterial growth and drug sensitivity. Anal. Chem.84:5250 –5256.
39. Delmas T, Piraux H, Couffin AC, Texier I, Vinet F, Poulin P, Cates ME, Bibette J.2011. How to prepare and stabilize very small nanoemulsions. Langmuir27:1683–1692.
40. Baraban L, Bertholle F, Salverda ML, Bremond N, Panizza P, Baudry J, de Visser JA, Bibette J.2011. Millifluidic droplet analyser for microbiol-ogy. Lab Chip11:4057– 4062.
41. Boitard L, Cottinet D, Kleinschmitt C, Bremond N, Baudry J, Yvert G, Bibette J.2012. Monitoring single-cell bioenergetics via the coarsening of emulsion droplets. Proc. Natl. Acad. Sci. U. S. A.109:7181–7186. 42. Wilson SM, al-Suwaidi Z, McNerney R, Porter J, Drobniewski F.1997.
Evaluation of a new rapid bacteriophage-based method for the drug sus-ceptibility testing ofMycobacterium tuberculosis. Nat. Med.3:465– 468. 43. Banaiee N, Bobadilla-Del-Valle M, Bardarov S, Jr, Riska PF, Small PM,
Ponce-De-Leon A, Jacobs WR, Jr, Hatfull GF, Sifuentes-Osornio J. 2001. Luciferase reporter mycobacteriophages for detection, identifica-tion, and antibiotic susceptibility testing ofMycobacterium tuberculosisin Mexico. J. Clin. Microbiol.39:3883–3888.
44. Smietana M, Bock WJ, Mikulic P, Ng A, Chinnappan R, Zourob M. 2011. Detection of bacteria using bacteriophages as recognition elements immobilized on long-period fiber gratings. Opt. Express19:7971–7978. 45. Traore H, Ogwang S, Mallard K, Joloba ML, Mumbowa F, Narayan K,
Kayes S, Jones-Lopez EC, Smith PG, Ellner JJ, Mugerwa RD, Eisenach KD, McNerney R.2007. Low-cost rapid detection of rifampicin resistant tuberculosis using bacteriophage in Kampala, Uganda. Ann. Clin. Micro-biol. Antimicrob.6:1. doi:10.1186/1476-0711-6-1.
46. Mole R, Trollip A, Abrahams C, Bosman M, Albert H.2007. Improved contamination control for a rapid phage-based rifampicin resistance test forMycobacterium tuberculosis. J. Med. Microbiol.56:1334 –1339. 47. Jorgensen JH, Ferraro MJ.2009. Antimicrobial susceptibility testing: a
review of general principles and contemporary practices. Clin. Infect. Dis. 49:1749 –1755.
48. Douglas IS, Price C, Overdier K, Thompson K, Wolken B, Metzger S, Howson D.2011. Rapid microbiological identification and major drug resistance phenotyping with novel multiplexed automated digital micros-copy (MADM) for ventilator-associated pneumonia (VAP) surveillance, poster A3928. ATS 2011 International Conference, Denver, CO, 13 to 15 May 2011.
on May 16, 2020 by guest
http://jcm.asm.org/
Alex van Belkumgraduated as a biologist at the University of Leiden, The Netherlands, in 1983. In 1988, Professor van Belkum did his Ph.D. examination in Biochemistry at the same uni-versity. In 1996, he received a second Ph.D. in Molecular Microbiology at the Erasmus Uni-versity, Rotterdam, The Netherlands. Between 1988 and 1990, he was involved in malaria vac-cine research as a research scientist at the Bio-medical Primate Research Centre (BPRC-TNO), Department of Infectious Diseases,
Rijswijk, The Netherlands. Between 1990 and 1991, he was the Head of the Department of Infectious Diseases, MedScand Ingeny B.V., Leiden, The Netherlands, after which he joined the Department of Molecular Biology, Diagnostic Centre (SSDZ), Delft, The Netherlands (1991 to 1994), as a staff member. In both positions, his focus was on the development of molecular tests for the detection and characterization of infectious pathogens. From 1994 until 2010, he was a staff member at the Erasmus University Medical Center Rotterdam (EMCR), Department of Medical Microbiology & Infec-tious Diseases, Rotterdam, The Netherlands. Between 2002 and 2010, van Belkum was the head of the Unit for Research and Development. Since 2003, he has been a Professor of Molecular Microbiology at Erasmus MC. From 2010 to 2011, he worked for bioMérieux as R&D Director in the La Balme Microbiology Unit. In 2011, Dr. van Belkum became the Corporate Vice President for Microbiology R&D at bioMérieux (La Balme les Grottes, France). In his current position at bioMérieux, he heads an international team of microbiology researchers in the field ofin vitrodiagnostics of bacte-rial diseases. Professor van Belkum has authored or coauthored more than 440 peer-reviewed publications, 100 chapters in books, and a variety of edi-torials, letters, etc. Dr. van Belkum is Editor in Chief of theEuropean Journal of Clinical Microbiology and Infectious Diseases.