The Control of Cell Motility and Differentiation by Ras Pathways
Richard Ian Tuxworth
MRC Laboratory for Molecular Cell Biology
University College London
A Thesis Submitted to the University of London
ProQuest Number: 10014310
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10014310
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
Acknowledgements.
Abstract.
The Ras Family of GTP-binding proteins have been implicated in the control of cellular growth, differentiation and motility. The regulation of proliferation by Ras has been extensively studied in mammalian cells bu t the control of cell motility has proved more difficult to study in these cells due to the dom inant effects on cell proliferation that occur w ith the perturbation of Ras function. This thesis examines the functions of RasG and a putative Ras Exchange Factor Aimless, in the control of cell motility, and the functions of RasG in the control of the onset of differentiation of Dictyostelium discoideum.
To understand the role of Ras in the control of motility 1 have investigated the effects of disrupting the rasG gene. Analysis of rasG null cells indicates that RasG plays an im portant role in the control of the cell motility and in the onset of early multicellular development, but is not required for proliferation. The rasG null cells show aberrant regulation of the actin cytoskeleton, a loss of polarity, reduced speed of motility and a failure in the final stages of cytokinesis. rasG null cells also exhibit a delay in the initiation of development. W hen developm ent does occur, it is asynchronous.
Table of Contents.
ACKNOWLEDGEMENTS... 2
ABSTRACT... 3
TABLE OF CONTENTS... 4
LIST OF FIGURES... 10
ABBREVIATIONS... 12
CHAPTER ONE: INTRODUCTION... 16
1.1. Ra s Su p e r f a m il yof Sm a l l GTPa s e s... 16
1.2. GDP-GTP M o l e c u l a r S w i t c h ... 17
1.3. Mo l e c u l a r Ba s iso fth e Sw it c h...18
1.4. Str u c t u r ea n d Bioc he m ic al Propertieso f Ra s... 21
1.4.1. Mechanism ofG T P Hydrolysis...21
1.4.2. Mechanism o f GAP Action...21
1.4.3. Mechanism o f GEF Activity...22
1.4.4. Effector Binding...23
1.4.5. Post-translational Modification...23
1.5. The Bio log ica l Fu n c t io n sof Ra s...25
1.5.1. Cellular Transformation - the Control o f Cell Proliferation and Morphology...26
1.5.2. Differentiation and Cell Fate Choice...2 7 1.5.3. Ras in the Control o f Normal Cell Motility...28
1.5.4. Ras Function in Saccharomyces cerevisiae...2 9 1.5.5. Ras Function in Schizosaccharomyces pombe...30
1.5.6. ras Gene Targeting in Mice...32
1.6. Ra s Sig n a l l in g Pa t h w a y s...33
1.6.1. Activation o f Ras...33
1.6.1.1.RasG EF s...33
1.6.1.2. Adapter Proteins...35
1.6.1.3. Receptors...36
1.6.2. Alternative Pathways fo r Ras Activation...36
1.6.3.1. RasGAP and N F-1... 38
1.6.3.2. Raf-1... 40
1.6.3.3. PI 3-kinase... 41
1.6.3.4. Other Ras Effectors... 42
1.6.4. Pathways Downstream o f Ras Effectors...43
1.6.4.1. The MAP kinase Cascade...43
1.6.4.2. PI 3-kinase Pathways... 45
1.7. De v e l o p m e n to f Dic t y o ste liu md is c o id e u m...48
1.7.1. The Stages o f Development....48
1.7.2. Differentiation and Morphogenesis...51
1.7.3. Signalling at the Onset o f Development....53
1.7.3.1. Pre-Starvation and Starvation Response o f Dictyostelium... 53
1.7.3.2. Pre-starvation Factor... 54
1.7.3.3. Conditioned Medium Factor... 55
1.7.3.4. CMF Signalling...56
1.7.4. Signalling Pathways During Aggregation...57
1.7.4.1. cAMP Receptors...57
1.7.4.2. Responses to Extracellular cA M P...58
1.7.4.3. Adaptation of the cAMP Response... 59
7.7.5. Signalling Pathways Controlling Differentiation...59
1.8. Dic t y o st e l iu m Ra s Pa t h w a y s...62
1.8.1. Functions o f Dictyostelium Ras Proteins...66
1.8.1.1.R as D ...66
1.8.1.2. RasG... 67
1.8.1.3. Other Dictyostelium Ras Proteins...67
1.9. Th e Dic t y o s t e l iu m RasGEP,A im l e s s ... 68
1.9.1. Disruption o f the aimless Gene...68
1.9.2. Specificity o f Aimless... 72
CHAPTER TWO: MATERIALS AND METHODS... 75
2.0. Mo l e c u l a r Bio l o g y... 75
2.0.1. Bacterial Strains and Propagation... 75
2.0.2. Preparation o f Competent Bacteria...75
2.0.3. Transformation o f Bacteria... 76
2.0.4. DNA Preparation... 76
2.0.5. Restriction Digestion o f DNA... 77
2.0.6. Agarose Gel Electrophoresis...78
2.0.7. Purification o f DNA Fragments from Agarose...78
2.0.8. Ligations...78
2.0.9. Polymerase Chain Reaction (PGR)...79
2.0.10. Sequencing o f DNA Constructs...79
2.0.11. Southern Blotting...79
2.1. Cl o n in g St r a t eg ie sa n d Pr im er s Us e d... 80
2.1.1. rasG Disruption...80
2.1.2. Aimless-CAAX in an Extrachromosomal Vector....80
2.1.3. Aimless-CAAX in discoidin-lnducible Vector....81
2.1.4. GST-Aimless...81
2.2.C e l l S t r a i n s , C u l t u r e a n d T r a n s f e c t i o n ... 82
2.2.1. Axenic Culture...82
2.2.2. Bacterial Culture...82
2.2.3. Transfection...83
2.3. Bio c h e m is t r y... 83
2.3.0. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)...83
2.3.1. Western Blotting...84
2.3.2. Immunological Detection o f Transferred Proteins...84
2.3.3. Luciferase Assays...85
2.4. Pu r ific a t io nof Rec o m b in a n t Pro te in sfr om Ba c t e r ia...85
2.4.1. Expression ofGST-fusion Proteins...85
2.4.2. Lysis o f the Bacteria...86
2.4.3. Purification o f the GST-fusion Protein with Glutathione-agarose...86
2.4.4. Gel Filtration o f the Purified GST-Ras Proteins...87
2.4.5. Cleavage o f GST-Aimless...87
2.5. Ex c h a n g e Fa c to r As s a y s... 87
2.5.1. Loading o f the GST-Ras Protein...87
2.5.2. Exchange fo r GTP...88
2.6. Mic r o sc o p y...88
2.6.1. Morphology and Movement o f Living Cells...88
2.6.2. Traction-Mediated Cytofission...89
2.6.3. Chemotaxis Assays...89
2.6.5. Effects ofA zide...90
2.6.6. Immunofluorescence o f Fixed Cells...90
2.1. D /cryo5r£L/[/M D e v e lo p m e n t ... 91
2.7.1. Induction o f Development....91
2.7.2. Assay o f PSFActivity...91
2.7.3. Preparation and Assay o f Conditioned Medium...91
2.7.4. Analysis o f Motility Changes...92
CHAPTER THREE: RASG IS A REGULATOR OF THE ACTIN CYTOSKELETON BUT IS NOT REQUIRED FOR GROWTH...94
3.1. In t r o d u c t io n... 94
3.2. G e n e r a t i o n o f r a sG n u l l c e l l l i n e s ...95
3.3. T h e E f f e c t s o f D is r u p t in g t h e r a sG g e n e ... 99
3.3.1. Growth Rate on Surfaces...99
3.3.2. Disruption o f Cell Polarity and Morphology...lOI 3.3.3. Disruption o f the Actin Cytoskeleton...103
3.3.3.1. Confocal M icroscopy... 103
3.3.3.2. Epifluorescence Microscopy... 105
3.3.3.3. Three Dimensional Renderings o f Confocal Z-series... 107
3.3.4. Failure in Cytokinesis...109
3.3.4.1. rasG' Cells Cannot Proliferate in Shaken Culture...109
3.3.4.2. rasG' Cells Form Huge Multinucleate Cells After Shaking...111
3.3.5. rasG' Cells Fail at the Final Stage o f Cytokinesis...113
3.3.5.1. Synchronisation of Cells...113
3.3.5.2. Microscopy of Cells in Cytokinesis...114
3.3.6. Myosin II Function is Normal in rasG' Cells...117
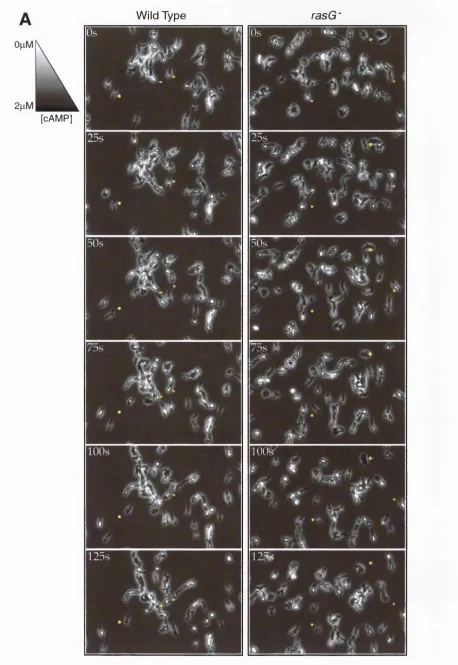
3.3.7. Chemotaxis is Normal in rasG' Cells...119
3.4 Dis c u s s io n... 123
3.4.1. RasG and Proliferation...173
3.4.2. Redundancy...174
3.4.2.1. Mouse Knock-outs... 125
3.4.3. Control o f the Actin Cytoskeleton...176
3.4.4. Loss o f Polarity...176
3.4.5. Alterations in F -Actin Structures...177
3.4.6. Cytokinesis Fails at the Final Stage...179
3.4.7. Chemotaxis Appears Normal...13]
CHAPTER FOUR: RASG IS REQUIRED FOR THE CORRECT INITIATION OF DICTYOSTELIUM DEVELOPMENT...134
4.1 . In t r o d u c t io n... 134
4.2. C o l o n y M o r p h o l o g y is A b e r r a n t in t h e rasC C e l l s ... 134
4.3 . PSF S e c r e t i o n a n d R e s p o n s e is N o r m a l in rasG' C e l l s ... 137
4.4. C M F S e c r e t i o n is N o r m a l in rasG C e l l s ... 139
4.5. rasG' C e l l s D o N o t R e sp o n d t o C o n d it io n e d M e d iu m ... 141
4.6. C h a n g e s in M o r p h o l o g y in R e s p o n s e t o C M F a r e R e d u c e d in rasG C e l l s ...141
4.7. AIMLESS N u l l C e l l s F a i l t o R e sp o n d t o C o n d it io n e d M e d iu m ... 145
4 .8 . A g g r e g a t i o n o f rasG' C e l l s is E s s e n t i a l l y N o r m a l ...148
4 .9 . Lat er De v e l o pm e n tis Mo rph o l o g ic a lly No r m a lb u tis As y n c h r o n o u s... 150
4 .1 0 . Dis c u s s io n...152
4.8.1. A Possible Defect in CMF Signalling...152
4.8.2. Signal Transduction from RasG...153
4.8.3. Comparison o f Differentiation in Dictyostelium and S. pombe...154
4.8.4. Differentiation o f PC 12 Cells...156
CHAPTER FIVE: ANALYSIS OF THE FUNCTIONS OF THE AIMLESS RAS EXCHANGE FACTOR... 157
5.1. In t r o d u c t io n... 157
5.2. Co n st r u c t io nofa Me m b r a n e Ta r g e t e d Aim less Pr o t e in... 157
5.3. E x p r e s s io n o f A im le s s -C A A X in Dictyostelium C e l l s ...160
5.4. The Effe c tof Ex p r e ssin gth e Aim l e s s-C A A X Proteinu n d e ra n In d u c ib l e Pr o m o t e r...163
5.4.1. Induction o f Aimless-CAAX Expression...163
5.4.2. Colony Morphology...165
5.4.3. Cell Morphology...165
5.4.4. Cell Motility...168
5.4.5. Cell Morphology by Immunofluorescence...172
5.5 The Specificityo f Ai m l e s s...174
5.5.1. Purification o f GST-Ras Proteins...174
5.5.2. Preparation o f GST-Aimless Proteins...174
5.6. Dis c u s s io n... 178
5.6.1. Activation o f Aimless...178
5.6.2. Potential Role fo r Aimless in Chemotaxis...179
5.6.3. Alternative Model fo r Aimless Function...181
5.6.4. The Target fo r Aim less?...181
CHAPTER SIX; FINAL DISCUSSION AND PROSPECTS... 184
L ist o f Figures»
Figure 1,1. The GTPase cycle... 20
Figure 1.2. Stages of the Dictyostelium Life Cycle... 50
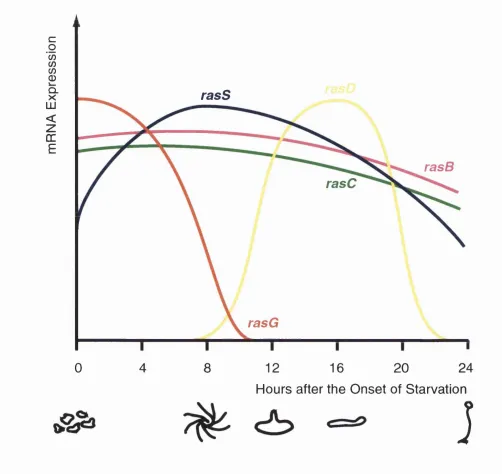
Figure 1.3. Expression of the ras Genes Through Dictyostelium Multicellular Development...63
Figure 1.4. AUgnment of Fluman Ha-Ras w ith the Dictyostelium Ras Proteins... 64
Figure 1.5. Alignments of Dictyostelium RasD and RasG w ith Fluman Ha-Ras... 65
Figure 1.6. Residues Im portant for Interaction between Ras Proteins and RasGEFs are Conserved in Aimless and Dictyostelium Ras Proteins...71
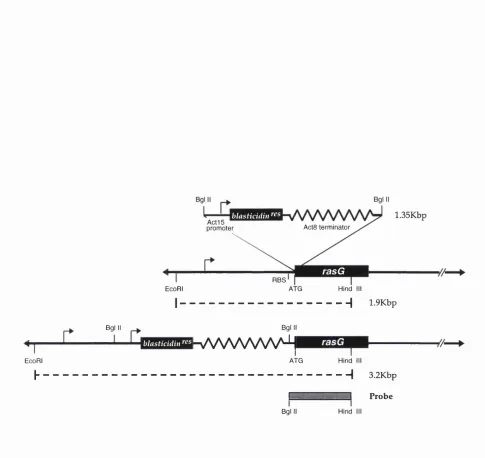
Figure 3.1. Schematic Representation of the Strategy Used to D isrupt the rasG Gene... 97
Figure 3.2. Analysis of Putative rasG- Clones...98
Figure 3.3. Normal Growth Rate of rasG- Cells on Surfaces...100
Figure 3.4. Loss of Polarity in rasG- Cells... 102
Figure 3.5. Aberrant Actin Cytoskeleton in rasG- Cells... 104
Figure 3.6. Distorted Actin Cortex in rasG- Cells... 106
Figure 3.7. Aberrant Actin Cortex in Flattened rasG- Cells... 108
Figure 3.8. Impaired Growth of rasG- Cells in Shaken Culture... 110
Figure 3.9. Multinucleate rasG- Cells Divide by Traction M ediated Cytofission 112 Figure 3.10. rasG- Cells Fail to Complete the Final Stage of Cytokinesis...116
Figure 3.11. Myosin II Activity is Present in rasG- Cells... 118
Figure 3.12. Normal Chemotaxis of rasG- Cells to Folate... 121
Figure 3.13. Normal Chemotaxis of rasG- Cells Towards cAMP... 122
Figure 4.1. Aberrant Colony Morphology in rasG- Cells... 136
Figure 4.2. Normal Induction of discoidin-1 y-luciferase by PSF-Conditioned M edium in rasG- Cells...138
Figure 4.3. Induction of the discoidin-ly Promoter by Conditioned M edium is Reduced in rasG- Cells... 140
Figure 4.4. Reduced Morphological Response to CMF in rasG- Cells...144
Figure 4.6. Reduced Morphological Changes in aimless Null Cells in Response to
Conditioned M edium ... 147
Figure 4.7. Normal Morphology and Aggregation of Developed rasG- Cells... 149
Figure 4.8. Morphologically Normal bu t Asynchronous D evelopment in rasG-CeUs...151
Figure 4.9. Comparison of the S. pombe Pheromone Pathway w ith the Dictyostelium CMF Pathw ay... 155
Figure 5.1. Construction of a Membrane-targetted Aimless Protein... 159
Figure 5.2. Aimless-CAAX Expression from the actinl5 Promoter... 162
Figure 5.3. Induction of Aimless-CAAX Expression by Conditioned M edium ...164
Figure 5.4. Failure to Aggregate in CeUs Expressing Aimless-CAAX...166
Figure 5.5. Aimless-CAAX Cells Display Multiple Actin-Rich Spikes... 167
Figure 5.6. Hypermotility and Multiple Pseudopod Extension in Cells Expressing Aimless-CAAX... 170
Figure 5.7. Aimless-CAAX Protein is Localised to the End of Actin Spikes... 171
Figure 5.8. GST-Ras and GST-Rapl Protein Preparations... 173
Figure 5.9. Preparation of the GST-Aimless Catalytic Domain...176
Figure 5.10. Effect of Aimless on GDP Exchange of Dictyostelium Ras and Rap Proteins...177
A bbreviations.
7-TM-R: seven-span transmembrane receptor ADP: adenosine diphosphate
AEBSF: 4-(2-aminoethyl)benzenesulphonyl flouride ARF; ADP-ribosylation factor
APS: ammonium persulphate Asm asparagine
ATP: adenosine triphosphate
cAMP: cyclic adenosine monophosphate cARl: cyclic AMP receptor 1
cAR2: cyclic AMP receptor 2 cAR3: cyclic AMP receptor 3 cAR4: cyclic AMP receptor 4
cDNA: complementary deoxyribonucleic acid CDKl: cyclin-dependent kinase inhibitors cGMP: cyclic guanosine monophosphate CMF: conditioned medium factor CSF-1: colony stimulating factor-1 dATP: deoxyadenosine triphosphate dCTP: deoxycytidine triphosphate dGTP: deoxyguanosine triphosphate dTTP: deoxythymidine triphosphate DIG: differential interference contrast DIF: differentiation inducing factor DNA: deoxyribonucleic acid
dsDNA: double stranded deoxyribonucleic acid DTT: dithiothreitol
ECM: extracellular matrix ecmA: extracellular matrix A ecmB: extracellular matrix B
EDTA: ethylenediaminetetraacetic acid EGF: epidermal growth factor
FGF: fibroblast growth factor
f-Met-Leu-Phe: formylated-methione-leucine-phenylalanine tripeptide GAP: GTPase activating protein
GBF: G-box factor
GFP: green fluorescent protein Gin: glycine
GSK3: glycogen synthase kinase 3 GST: glutathione-S-transferase GDP: guanosine diphosphate GEF: GTPase exchange factor Gin: glutamine
Gly: glycine
GTP: guanosine triphosphate HRP: horseradish peroxidase IP3: inositol trisphosphate
IPTG: isopropyl P-o-thiogalactopyranoside LB: Luria broth
NF-1: neurofibromatosis type 1 NGF: neural growth factor
PAGE: polyacrylamide gel electrophoresis PB: phosphate buffer
PBS: phosphate-buffered saline PGR: polymerase chain reaction PDGF: platelet-derived growth factor PEG: polyethylene glycol
PFÎ: pleckstrin homology PI: phosphatidylinositol
PIP: phosphatidylinositol monophosphate PIP2: phosphatidylinositol 3,4-bisphosphate
PIP3: phosphatidylinositol 3,4,5-trisphosphate
PKA: protein kinase A PKB: protein kinase B PEG: phospholipase G PSF: pre-starvation factor pstA: prestalk A
pstB: prestalk B pstO: prestalk O Rb: retinoblastoma RNA: ribonucleic acid
RTK: tyrosine kinase receptor SAP: stress activated protein SDS: sodium dodecyl sulphate Ser: serine
SH2: Src homology 2 SH3: Src homology 3
STAT: signal transducer and activator of transcription TEMED: N, N, N% N'-tetramethylethylenediamine Thr: threonine
TMGF: traction-mediated cytofission tRNA: transfer ribonucleic acid Tyr: tyrosine
Val: valine
Chapter One: Introduction,
1.1. Ras Superfamily of Small GTPases.
The Ras superfamily of small GTPases comprises a large num ber of closely related proteins that have been implicated in the control of a wide range of cellular processes including the control of proliferation and differentiation, the regulation of vesicle transport and the control of cell motility and polarity. Members of the superfam ily can be divided by sequence hom ology into five sub-families of proteins - the Ras, Rho, Rab, Arf and Ran sub-families.
Ras: The functions of Ras proteins have been studied intensively since the discovery in the early 1980s that mutations in ras genes underlie some retrovirally induced tum ours (Ellis et a l, 1981). Ras proteins play fundam ental roles in the control of cellular growth, differentiation and motility and will be discussed in detail below.
Rho: M embers of the Rho subfam ily were discovered by low stringency screening for Ras-like genes (Madaule and Axel, 1985). The family is further subdivided into groups, the best characterised consisting of multiple Rho, Rac and Cdc42 proteins that have been shown to control cell motility, adhesion and polarity th ro u g h m od u latio n of the actin cytoskeleton. M ore recently, roles in transcriptional activation and cellular transform ation have also been highlighted (Hall, 1998).
Rab: The Rab family of GTPases has been implicated in the regulation of vesicle transport w ithin the cell. When GTP is bound the Rab proteins can regulate the fusion of vesicles w ith their target membranes. The fusion stim ulates GTP hydrolysis and causes the dissociation of the GDP-bound Rab from the target membrane. The Rab is then recycled back to the donor membrane where exchange for GTP occurs and the cycle repeats (Zerial, 1995).
trafficking of vesicles, including intra-Golgi transport, endoplasmic reticulum-Golgi transport and the fusion of endosomes. Like Ran proteins, the cycling between the GDP-GTP bound states is im portant for Arf function, w ith the GTP-bound form only having a high affinity for membranes (Kahn, 1995).
Ran: The Ran proteins are unique members of the superfamily of small GTPases in that they reside in the nucleus rather than the cytoplasm. The proteins are thought to regulate the entry of nuclear localisation sequence-containing proteins into the nucleus and the export of mRNA from the nucleus. Ran proteins may also function in the coupling of mitosis to the completion of DNA synthesis (Birschoff et a l, 1995).
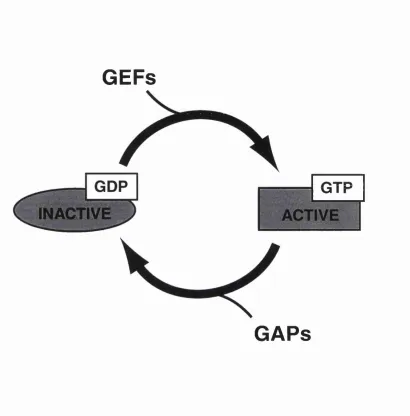
1.2. GDP-GTP Molecular Switch.
The Ras superfamily is part of a much larger family of proteins that use a highly conserved m echanism of action to regulate w idely different processes. The mechanism is commonly referred to as a "m olecular switch" since it involves cycling of the protein betw een an inactive and an active conformation. Every protein binds the nucleotides guanosine diphosphate (GDP) and guanosine triphosphate (GTP), They are inactive w hen GDP is bound but become active and are able to bind effector molecules w hen the GDP is exchanged for GTP. An intrinsic GTPase activity that hydrolyses the bound GTP to GDP forces the cycle to proceed in one direction only (Bourne et a l, 1990).
A short description of three of the more divergent proteins that utilise the GDP- GTP switch illustrates the processes that are controlled by this mechanism. Firstly, elongation factor EF-Tu from Escherichia coli uses a GDP-GTP switch during ribosomal protein synthesis to control binding of aminoacyl-tRNA molecules in the ribosome and to proof-read the selection of the appropriate tRNA molecule for the codon. Only w ith GTP bound are EF-Tu molecules able to bind the aminoacyl- tRNA. When a EF-Tu-GTP-tRNA complex forms the aminoacyl-tRNA is brought to the mRNA in the ribosome where the anticodon can associate w ith the mRNA codon. The association of EF-Tu with the ribosome triggers GTP hydrolysis, bu t the speed of GTP hydrolysis allows a kinetic proof-reading step to take place - the hydrolysis step is slow and m ism atched anticodons and codons will usually dissociate before hydrolysis is complete thus rejecting the incorrect aminoacyl- tRNA (Thompson et a l, 1986; Thompson, 1988).
Another system that uses the switch mechanism is the transduction of signals from transmembrane hormone receptors or from photoreceptors. The a-subunits of heterotrimeric G-proteins only bind to the receptor when it is activated (in these cases by ligand binding to the extracellular domain, or by photon activation). Only the a-subunit of the G-protein binds guanine nucleotides, and it is the interaction of the G-protein w ith the receptor that allows exchange of GDP for GTP to occur on the a-su b u n it and consequent activation of the G-protein. W hen inactive (with GDP bound), the a-subunit is associated w ith the py-dimer, but activation leads to dissociation, leaving both the a-subunit and the py-dimer free to bind to and activate downstream effector molecules. The intrinsic hydrolysis of GTP bound to the a-subunit limits the period of activation of the G-protein but provides sufficient time for a large amplification of the signal to occur (Neer, 1995).
Microtubule polymerisation and depolymerisation is also controlled by a GDP- GTP switch involving the tubulin dimers that make up the filaments. a-P dimers are able to add to the plus end of the growing microtubule filament more rapidly and form tighter interactions with neighbouring dimers w hen GTP is bound to the p-subunit. After polymerisation, the GTP is slowly hydrolysed to GDP by the intrinsic GTPase activity of the subunit. With GDP bound, the a-P dimers interact less strongly with other dimers and are more likely to dissociate from the filament, w ith dissociation occurring primarily at the minus (Mitchison and Kirschner, 1984; Alberts et a l, 1994).
Actin filament polymerisation and depolymerisation is regulated by a similar mechanism to microtubules. However, in the case of actin, ATP rather than GTP binds to the actin monomers and is hydrolysed to ADP following polymerisation of the monomers. The ATP bound actin monomers have an increased affinity for other monomers as with GTP-bound tubulin dimers. Similarly, hydrolysis to ADP decreases the affinity and promotes depolymerisation. The structure of actin bears little homology to that of tubulin despite the similarity in mechanism and probably represents a case of convergent evolution.
1.3. Molecular Basis of the Switch.
that hydrolyses the GTP molecule and returns the protein to the inactive form (figure 1.1; Bourne et a l, 1991).
The affinity of GTPases for guanine nucleotides is extremely high (Kd values are between 10-n and 10-^ M) and since the cytosolic concentrations of GDP and GTP are considerably higher than the Kd values of the GTPase (lO-^ M for GDP and approximately 10 times higher for GTP), nucleotides will always be bound in vivo (Bourne et ah, 1991). Consequently the switch from GDP to GTP cannot be made by the GTPase simply responding to small changes in the concentrations of the nucleotides, rather the switch m ust be catalysed by a second protein. The class of proteins that catalyse the exchange - GTPase exchange factors (GEFs) - bind to the GTPase in its GDP-bound form and allow the GDP to dissociate (figure 1.1). The GTPase is now in the nucleotide-free form and can form a stable complex w ith the GEF although GTP will usually enter the nucleotide binding site rapidly, causing the GEF to dissociate (Lai et a l, 1993). The higher level of GTP over GDP in the cytosol means that GTP will normally enter the binding site in preference to GDP.
The GTP-binding protein is now in its activated confirmation and is capable of interacting with effector molecules but does not remain so indefinitely. The period of activation is limited by the intrinsic GTP hydrolysing activity of the molecule. The intrinsic GTP hydrolysis rate varies in different systems. Hydrolysis of GTP bound to the a-subunits of heterotrimeric G-protems following receptor activation varies but is generally slow (a half life of 0.5 - 10s), allowing time for the interaction to occur between the a-subunit and its effector and amplify the original signal (Carty et a l, 1990). In the case of EF-Tu GTP hydrolysis does not occur at all until the EF-Tu-GTP-aminoacy 1-tRNA complex has associated w ith the ribosome because prem ature hydrolysis w ould lead to dissociation of the complex and release of the aminoacyl-tRNA cargo. However, once associated w ith the ribosome hydrolysis occurs rapidly (Thompson, 1988).
The intrinsic GTP hydrolysis rate can be stim ulated by GTPase activating proteins (GAPs) which return the GTPase protein to its inactive conform ation (figure 1.1). Both GEFs and GAPs are im portant regulators of GTPase function. The loss of GAP activity can results in a GTPase protein that is constitutively in its activated GTP-bound conformation, w ith consequences that can be as severe as cellular transformation and tum our formation. For example the loss of a RasGAP, encoded by the NF-1 gene, either through naturally occurring hum an mutations or by gene targeting in mice, leads to a predisposition to neurofibromas (Brarman et a l, 1994).
GEFs
INACTIVE
\JI 1 “
GAPs
Figure 1.1. The GTPase cycle.
The GTPase protein cycles between an inactive state w ith GDP bound and an activated form w ith GTP bound. Only when GTP is bound can the GTPase interact with effector molecules. The exchange of GDP for GTP is catalysed by GTP exchange factors (GEFs). GTPase activating proteins (GAPs) stimulate the hydrolysis of GTP to GDP and thus promote the inactivation of the GTPase protein.
1.4. Structure and Biochemical Properties of Ras.
The families of proteins that use the GDP-GTP switch share little prim ary sequence hom ology but, unsurprisingly considering the sim ilarities in their mechanisms of action, they share closely related three dimensional structures. The structure of Ras and how the structure relates to its function will be described below.
1.4.1. Mechanism of GTP Hydrolysis.
The three dim ensional structure of Ras bound to a non-hydrolysable GTP analogue, GppNHp, was solved to a resolution of 1.35Â in 1990 (Pai et ah, 1990) and allowed six conserved motifs to be identified that correspond to motifs found in other GTP-binding proteins. These motifs are involved in binding of the nucleotide and the essential Mg^+ ion, and also in the hydrolysis of GTP. A mechanism for the hydrolysis of GTP was suggested that requires a catalytic water molecule (W atl 75) to be positioned by the glutamine residue at position 61. The w ater molecule launches a nucleophilic attack on the y-phosphate w ith the Gln61 residue also stabilising the leaving phosphoryl group (Pai et ah, 1990). The Gln61 plays an im portant role in the GTP hydrolysis reaction, suggested not only by the crystal structure but also by the strong transforming ability of Ras proteins m utated at this residue. It is surprising, therefore, that it is not present in all GTPases. In Rap, which shares large homology with Ras and appears to bind the same effectors, the glutamine residue is replaced by threonine. In EF-Tu, histidine is present. These differences may reflect different mechanisms of GTP hydrolysis or may suggest that the proposed mechanism is incorrect.
1.4.2. Mechanism of GAP Action.
The m echanism by which GAP stimulates the hydrolysis of GTP has been proposed for Ras following the solving of the crystal structures of the catalytic dom ain of p i 20 RasGAP alone, and in complex w ith Rapl-GTP (Scheffzek et al., 1996), and subsequently of RasGAP in complex w ith Ras-GDP and AIF4- (Scheffzek
et al., 1997). AIF4- has long be know n to be capable of activating GDP-bound heterotrimeric G-proteins but not small GTPases such as Ras. The reason for this anomaly is that the a-subunits of heterotrimeric G-proteins contain a small GAP domain that provides a relatively rapid hydrolysis of GTP w ithout the requirement
for a second protein. Complexing Ras-GDP w ith GAP provides the GAP domain found in the a-subunits and allows AIF4- to bind (Scheffzek et a l, 1997).
The structure of Ras and p i 20 RasGAP provided a model for GAP activity in w hich an "arginine finger" is inserted by GAP into the catalytic site of Ras and stabilises the transition state of the hydrolysis reaction. The im portance of the arginine residue in the finger is demonstrated by its conservation in GAP molecules th at interact w ith Ras or w ith other small GTPases, and also by the inability to substitute this residue w ith any other (even a conservative substitution such as lysine) w ithout losing GAP activity (Ahmadian et a l, 1997). Solving of the structure also indicated w hy the residues at positions 12 and 61 are such potent activators when mutated. The glycine residue at position 12 is extremely close to the arginine finger inserted by GAP and to the glutamine residue at position 61. Any m utation of Gly 12, even to alanine, results in displacement of Gln61 due to the more bulky side chain and loss of GAP activity. However, the m utations do not affect the ability of Ras to bind to GAP. Gln61, meanwhile has an essential role in stabilising the transition state and mutation to most residues disrupts the interaction w ith the arginine finger. Again, m utations do not affect binding of GAP, merely GAP function.
1.4.3. M echanism of GEF Activity.
complex after GDP has left the nucleotide binding site. Hence the A snl7 proteins bind to and sequester GEFs.
1.4.4. Effector Binding.
The residues im portant for the binding of effectors to Ras have been identified by several m utational analyses that searched for m utations that abolished Ras signalling w ithout affecting GTP binding. Most residues have been confirmed as im p o rtan t by the three dim ensional structure of Raf-1 complexed w ith R apl (Nassar et a l, 1995). The region termed the "Effector Domain" lies between resides 32-40 and is highly conserved in Ras proteins from all species. The Effector Domain has subsequently been the subject of large scale m utational analyses aim ed at determining the exact residues required for the binding and activation of the three best characterised Ras effector molecules - Raf, Ral and PI 3-kinase. Point substitutions have been identified that abolish the binding of two of the three know n Ras effectors whilst leaving the binding and activation of the third effector intact (White et a l, 1995; Rodriguez-Viciana et a l, 1997). These m utations have proved extremely useful tools for the study of Ras function since they allow the effects of activating Ras to be delineated into three distinct biochemical pathways - a pathway involving the activation of Raf, another involving PI 3-kinase and a third involving Ral-GDS. The signalling pathways downstream of Ras wül be discussed in detail below.
1.4.5. Post-translational M odification.
Ras proteins i are
post-translationally m odified by the addition of one or more lipids to the extreme carboxy terminus of the protein. The modifications allow association of the protein with membranes (in the case of Ras w ith the inner side of the plasma membrane) although some other small GTPases are localised to internal membranes. The im portance of the post-translational m odification can be dem onstrated by expressing m utant Ras proteins that cannot be modified. These proteins have no in vivo activity in locus formation assays for transforming ability and are localised to the cytosol (Wülumsen et a l, 1984a; Wülumsen et al, 1984b).
The lipid modifications are directed by an extreme carboxy terminal sequence. The sequence is CAAX (where G is cysteine, A is an aliphatic amino acid and X is usually methionine, serine or leucine) and it provides the commonly used named
for this type of m embrane targeting sequence - the CAAX-box. The sequence is recognised by a prenyl transferase that covalently adds the lipids to the cysteine residue of the sequence, after which the term inal three amino acid residues are proteolytically cleaved. CAAX farnesyl transferase was the first prenyltransferase to be identified and cloned. It recognises with high affinity CAAX sequences where the terminal X residue is methionine (found in the m ammalian Ki-Ras and N-Ras) and w ith ten fold less affinity those sequences which end in serine, for example Ha- Ras. The enzyme adds a farnesyl lipid to the cysteine residue (Seabra et a l, 1995a). CAAX g eran y lg eran y l tran sferase is responsible for the a d d itio n of a geranylgeranyl lipid to the CAAX-box of many small GTPases including Rho, Rac, R apl and (one presumes) the majority of the Dictyostelium discoideum Ras proteins. It recognises CAAX sequences ending in leucine (Seabra et a l , 1995b). Both prenyltransferases are thought to be cytosolic enzymes.
The loss of transform ing ability of Ras th at corresponds to a lack of lipid modification has suggested that the CAAX prenyl transferases are potential targets for therapeutic antitum our drugs. Much research is currently underw ay aimed at finding specific inhibitors of these enzymes, w ith m ost w ork concentrated on CAAX farnesyl transferase. M ost sm all GTPases oth er th an Ras are geranylgeranylated so inhibiting only the farnesyl transferase m ay reduce unw anted side effects (Gibbs et al, 1994).
The addition of a farnesyl or geranylgeranyl lipid to Ras is required bu t is not sufficient to target the protein to the plasma membrane. A second targeting motif is required in the carboxy terminus of the protein (Hancock et a l , 1991). In both H um an Ha-Ras and N-Ras proteins two cysteine residue at positions 181 and 184 are palmitoylated but these lipids are only added if the farnesyl lipid is first added to the term inal cysteine. The palmitoylated lipids are responsible for membrane anchoring of the protein although the mechanism of anchoring is uncertain - the lipids may interact w ith a protein component of the membrane or simply insert into the phospholipid bilayer.
Successively replacing the lysines w ith uncharged residues reduces the affinity of Ki-Ras for the plasma membrane and alters the cellular localisation of the protein to w ard s cytosolic. At least two lysine residues are required for m em brane association.
1.5. The Biological Functions of Ras.
The ras genes were first discovered as viral oncogenes following the study of acutely transforming Harvey and Kirsten m urine retroviruses. The initial studies were followed by the recognition in the early 1980s that the viral ras genes are m utated forms of normal cellular genes (DeFeo et a l, 1981; Ellis et a l, 1981; Tabin et a l , 1982; Taparowsky et a l, 1982; Capon et a l, 1983). Biochemical analysis of the products of the viral and cellular ras genes dem onstrated the guanine nucleotide binding capabilities of the Ras proteins and the failure of the m utated viral form to hydrolyse GTP. This observation led directly to the elucidation of the mechanism behind the ability of the viral Thr-24-mutated Ras proteins to induce tum ours and cellular transformation (Sweet et a l, 1984).
The majority of studies over nearly twenty years following the identification of the ras genes has been aimed at an understanding the growth control function of the proteins. H ow ever, the m orphological changes associated w ith Ras transform ation also implicate Ras in the control of cell motility, polarity and m orphology. In addition, Ras signalling pathw ays are ubiquitously found in eukaryotes and have been shown to control process as seemingly diverse as cell fate decisions and differentiation in worm s and flies (Kayne and Sternberg, 1995; W assarman et a l, 1995); apoptosis (Kauffmann-Zeh et a l, 1997), cell cycle arrest and senescence in m am m alian cells (Lloyd, 1998); cytoskeletal rearrangem ents and pinocytosis (Bar-Sagi and Feramisco, 1986); cytokinesis (Ng et a l, 1992); chemotaxis in Dictyostelium (Insall et a l, 1996) and pheromone signalling in yeast (Fukui et al, 1986). Recently, a role for Ras signalling has even been described for synaptic transmission and long-term memory (Brambilla et a l, 1997). This description of the biological properties of Ras will describe the evidence that suggests such functions for Ras before describing the signalling pathways and molecules that mediate the effects of Ras in these processes.
1.5.1. C ellu lar T ran sfo rm atio n - th e C o n tro l of C ell P ro life ra tio n an d Morphology.
Cellular transform ation requires several process to be corrupted. Cells are w idely categorised as transform ed w hen capable of serum free and anchorage independent growth, when they lose contact inhibition of growth and w hen they gain the ability to form tum ours in nude mice, v-ras genes are capable of transform ing im m ortalised cell lines directly and produce aggressive highly m etastatic tum ours (P a rtin et a l, 1988). In contrast, v - s r c is also capable of transforming cells but tends to produce tum ours of much less metastatic potential (Tatsuka et a l, 1996). These effects are obvious indications that Ras has a role in normal cell proliferation and motility.
The ability of Ras to transform cell lines was dem onstrated directly by microinjection studies using purified recom binant Ras proteins in the m id 1980s (Feram isco et a l, 1984; Stacey and Kung, 1984). Microinjection of the viral Ras protein (Thr24) caused rapid morphological changes including rounding of the cells, membrane ruffling and blebbing and also stimulated proliferation (Feramisco et a l, 1984). Normal Ras protein failed to produce these effects. The v-Ras protein was also shown to be able to transform only im m ortalised cell lines and not prim ary cells, an observation that supported the idea of oncogene co-operation in transformation (Land et a l, 1983). Indeed, activated Ras proteins induce senescence in primary cell cultures and not proliferation (Ridley et a l, 1988; Lloyd, 1998).
A role for the norm al, non-m utated form of Ras in the control of cell proliferation was demonstrated by injection of a blocking antibody to Ras, Y13-259, into non-transform ed cells. The antibody prevented serum induced grow th and dem onstrated the essential nature of Ras in this process (Mulcahy et a l, 1985). The pathways leading to Ras from growth factor receptors and downstream from Ras to the transcriptional apparatus that m ediate its effects on proliferation have been heavily studied and will be described below.
1986). The effects of Ras on cell morphology are now thought to be m ediated by small GTPases of the Rho family, as will be described below.
1.5.2. D ifferentiation and Cell Fate Choice.
A third function for Ras was also identified through the microinjection studies initiated in the mid 1980s. This function is the regulation of differentiation. PC12 cells differentiate in culture w hen neural grow th factor (NGF) is added to the grow th m edium and adopt a morphology reminiscent of sympathetic neurones, w ith long branching neurites. Injection of v-Ras leads to differentiation and w ithdraw al from the cell cycle in the absence of NGF, whereas non-m utated Ras has no affect (Bar-Sagi and Feramisco, 1985). More recently, the small GTPase, R apl, has been implicated in the differentiation of PC12 cells and it now appears that Rap and Ras regulate different but complementary aspects of the response (York et a l, 1998).
Ras was independently implicated in differentiation and in the control of cell fate choice in two different genetic systems - the differentiation of photoreceptors in the com pound eye of the fruit fly. Drosophila, and in the differentiation of the vulva in the nematode, C. elegans. The Drosophila eye consists of several repeated units known as orrunatidia, each of which is made of eight photoreceptor cells that differentiate in a stereotyped order. Of these cells the differentiation of the R7 cell is dependant on a Ras signalling pathway that leads from a cell surface receptor to transcription factors and which is homologous to the Ras signalling pathways that regulate proliferation and cell cycle entry in m ammalian cells (W assarman et a l, 1995). An activating Ras mutation leads to all photoreceptors differentiating as R7 cells, whereas a loss of function Ras m utation prevents differentiation of the R7 cell (Fortini et al, 1992). Vulval differentiation is similarly regulated by a Ras pathw ay that is homologous to the MAP kinase pathw ays of mammalian cells (Kayne and Sternberg, 1995). Again, an activated Ras leads to excess differentiation so that multiple vulvae form, whereas loss of function Ras alleles result in a vulvaless phenotype (Beitel et al, 1990).
Drosophila and C. elegans have proved invaluable systems for studying Ras signalling pathw ays because the affects of activating and inhibitory mutations can be studied in isolation from effects on proliferation. In addition the ability to order com ponents of the signalling pathw ay by genetic epistasis experim ents has provided a com plem entary in vivo approach to the biochemical analysis of Ras signalling in mammalian cells.
D ifferentiation is often associated w ith w ith d raw al from the cell cycle. Interestingly, recent studies have begun to investigate an often overlooked phenom enon - th at Ras produces a grow th arrest in p rim ary cells, no t transformation (Ridley et a l, 1988). Activated Ras proteins co-operate w ith cell cycle inhibitor proteins such as the cyclin-dependant kinase inhibitors to w ithdraw prim ary cells from the cell cycle perhaps reflecting a mechanism by which cells can initiate differentiation in response to extracellular signals (Lloyd et a l, 1997; Serrano et a l, 1997).
1.5.3. Ras in the Control of Normal Cell Motility.
The changes in cell m otility that accom pany transform ation are extreme examples of Ras activity. Transform ation of cells by transfection w ith v-ras correlates w ith increased chemotactic and chemokinetic responses (Varani et a l, 1986; Ochieng et a l, 1991) but one problem in using transformed cells is that the changes in motility can only be correlated w ith v-ras expression. The increased motility could be indirectly due to the cellular transform ation process or due to instability in the phenotype of the v-ras transform ed cells. For example, stable embryonic cell lines expressing v-ras rapidly become heterogeneous in their characteristics, including their metastatic potential. To understand the effects of Ras on normal cell motility a more appropriate approach is to study the effects of activating or inhibiting Ras over short term periods, for example by microinjection.
W ound healing assays are commonly used as an in vitro model of the complex interactions that occur during the healing of an actual w ound. One im portant aspect of the healing response is an increase in m otility of fibroblasts and endothelial cells th at w ill enable the w o u n d to be closed rap id ly and for angiogenesis to occur. Dominant active Ras was shown to increase the motility of prim ary endothelial cells in a w ound assay and also to increase the random ness of movement, as if the controls that allow the cells to m igrate in a co-ordinated manner in the w ound had been disrupted (Sosnowski et a l, 1993). Conversely, cells injected w ith a blocking anti-Ras antibody w ere alm ost completely immobile. Fibroblast grow th factor (FGF) is a potent activator of endothelial cell motility in these assays, bu t the blocking anti-Ras antibody inhibited the FGF effects implying that FGF acts via Ras. Activation of Ras following binding of grow th factors to their receptors is a well characterised phenomenon which is described below.
directly in the control of chemotaxis of non-transform ed fibroblasts tow ards a source of PDGF (Kundra et a l, 1995). The assay used (migration w ithin a Boyden chamber) makes the distinction between chemotaxis and chemokinesis difficult, but transient transfection of dom inant negative ras constructs inhibited the response without inhibiting general cell motUity,
Of importance to this thesis, a RasGEF was identified in Dictyostelium that is req u ired for the chem otactic response to cAMP d u rin g aggregation, again implicating a Ras pathw ay in the control of chemotaxis (Insall et a l, 1996). As w ith the inhibition of fibroblast movement towards PDGF by dominant negative Ras, the general motility of the aimless null cells was not affected by the loss of the RasGEF, merely the ability of the cells to migrate towards the source of the chemoattractant. Combined w ith the loss of directionality of the endothelial cells injected w ith activated Ras in a w ound healing assay, the discovery of requirem ent for Ras for chemotaxis would appear to an im portant indicator that Ras is involved in the regulation of normal cell motility.
1.5.4. Ras Function in Saccharomyces cerevisiae.
The budding yeast Saccharomyces cerevisiae has two RAS genes identified by low stringency hybridisation in the early 1980s (DeFeo-Jones et a l, 1983; Powers et a l, 1984). The S. cerevisiae RAS genes share extensive homology w ith their mammalian counterparts, especially in the N-termini, but have an extended C-terminus. They also appear to regulate ostensibly comparable processes - cell proliferation and differentiation. Indeed, hum an Ha-ras is capable of substituting for the yeast genes (Kataoka et a l, 1985). However, the effector pathw ay controlled by the yeast Ras proteins is unique and has no obvious homologue in higher eukaryotes.
The two yeast RAS genes, RA Sl and RAS2, are redundant and either can be disrupted without effect. The rasl-ras2- double m utant is non-viable and indicates that Ras function is required for germ ination of spores and vegetative grow th (Kataoka et a l, 1984; Tatchell et al, 1984). An indication of a role for Ras in the yeast differentiation response was provided by expression of an activated m utant which blocked sporulation in response to starvation.
The growth arrest associated w ith loss of both RAS genes is phenotypically similar to that associated w ith loss of adenylate cyclase activity (Toda et a l, 1985). This similarity led to the demonstration that adenylate cyclase is a direct target of the Raslp and Ras2p proteins and can also be activated by hum an Ha-Ras (Broek et a l, 1985). Activation of adenylate cyclase leads to cAMP synthesis that is required
for progression past START and entry in to the cell cycle (M atsumoto et a l, 1982). Adenylate cyclase is the only target for the yeast Ras proteins that has been identified to date but it is not a Ras effector in mammalian cells. The mammalian adenylate cyclase shares no structural homology w ith the yeast protein although Ha-Ras can substitute for the yeast Rases in the activation of yeast adenylate cyclase. The function of Ras in yeast is conserved in that it is also essential for the entry of mammalian cells into the cell cycle in response to growth factors (Mulcahy et a l, 1985).
More recent analysis of Ras function in yeast has dem onstrated a second function, in addition to the activation of adenylate cyclase, that may reflect a novel function in mammalian cells. In rasl'ras2- cells growth abnormalities occur even if the failure to activate adenylate cyclase is bypassed, indicating that a second function for Ras probably exists. Moreover, a triple m utant where the ras-related gene, RSRl, is also disrupted fails to grow at any tem perature even w ith the adenylate cyclase block bypassed. These cells enter the cell cycle correctly but arrest late in M-phase, suggesting that Ras is required for M-phase completion (Morishita et a l, 1995). The block can be rescued by several proteins know n to function at this stage of the cell cycle but also by expression of mammalian Ha-Ras. This function of Ha-Ras may be conserved in mammalian cells, but Ha-Ras can also activate yeast adenylate cyclase despite the fact th at this do not occur in mammalian cells. The actions of Ras late in the cell cycle are not easy to study in mammalian cells because loss of function leads to growth arrest in G1 before the restriction point.
1.5.5. Ras Function in Schizosaccharomyces pombe.
The single ras gene in the fission yeast Schizosaccharomyces pombe shares greater homology w ith its counterparts in higher eukaryotes than do the two S. cerevisiae genes in that they do not have an extended C-terminus (Fukui and Kaziro, 1985). R aslp is not required for growth in S. pombe but is required for the mating response (Fukui et a l, 1986). It is an im portant com ponent of the signalling pathw ay that directs the pheromone response and sexual differentiation, rasl- cells are sterile and fail to agglutinate, sporulate or induce the gene encoding the pherom one receptor {m a m 2 ;X u et a l, 1994). They also exhibit altered m orphology, being rounded instead of the normal elongated shape. However, Rasl is not required for growth, nor for the regulation of adenylate cyclase as the two Ras proteins are in S.
Haploid S, pombe cells secrete a mating pheromone upon starvation that induces growth arrest and morphological changes in haploid cells of the other mating type. In contrast, the S. cerevisiae pheromones are secreted constantly. The pheromone binds to the G-protein linked seven-pass transm em brane receptor encoded by either the m aml or map3 genes (depending on mating type), causing the release of (3y-subunits. The Ga- subunit is responsible for activation of a MAP kinase cascade com prising the products of the byr2, hyrl, and s p k l genes (hom ologues of Raf/MEKK, MEK and MAP kinase respectively), that leads eventually to induction of gene expression (Herskowitz, 1995). R aslp lies at the head of the MAP kinase pathw ay th at results in the induction of gene expression bu t despite a direct interaction, R aslp is not sufficient to fully activate the MAP kinase cascade. Full activation, as determ ined by the induction of the mam2 gene, requires a second co operative signal from the a-subunit of an heterotrimeric G-protein, encoded by the g p a l gene, itself activated by the binding of the sexual pherom one of the other mating type to the seven-pass transmembrane receptor on the cell surface (Xu et a l, 1994). The phenotype caused by the loss of R aslp is more extensive than the defects caused by the loss of Byr2p, the Raf homologue. The byr2- cells show identical defects in agglutination, sporulation and lack of gene induction as the rasl-cells but exhibit normal, elongated morphology (Wang et a l, 1991). This difference suggests that a second signalling pathw ay is activated by R aslp th at directs changes in cell morphology, and indeed a second pathw ay was discovered in a genetic screen for cells unable to mate successfully but which were also rounded (Chang et a l, 1994). One of the genes identified in this screen encodes a homologue of the S. cerevisiae CDC24p, an exchange factor for the small GTPase CDC42p, which is involved in bud site selection in yeast and in the regulation of actin polymerisation and motility in mammalian cells (Tapon and Hall, 1997). A second gene product discovered in the same screen. Sc d ip , does not interacts directly w ith R aslp nor w ith the S. pombe Cdc42p, bu t will interact w hen a third protein identified in the screen, Scd2p, is added to the interaction assay. scd2 encodes a homologue of the S. cerevisiae protein BEMlp, and may act as a scaffold that will bring together the signalling molecules of the pathw ay -R aslp, Scdlp, and Cdc42p (Herskowitz, 1995).
the outw ard similarities are matched by the similarities in the signaUing pathways that control the processes. The differentiation of Dictyostelium is described below.
1.5.6. ras Gene Targeting in Mice.
The functions attributed to mammalian Ras proteins are in the main the result of the num erous studies that have expressed dom inant active or inhibitory forms and observed the effects. A more subtle method of investigating the functions is to remove the protein genetically by gene targeting. By this m ethod it is possible to develop a picture of the functions for which Ras is genetically required. Mice deficient for Ki-Ras (Johnson et a l, 1997; Koera et a l, 1997) and N-Ras (Umanoff et a l, 1995) have been generated but did not produce the expected phenotypes. The evidence that Ras is essential for proliferation emanating from the studies w ith dom inant proteins were not confirmed in the Ras-deficient mice. The N-ras knock out mice develop entirely norm al and are fully fertile, thus dem onstrating no requirement for N-ras at all. Ki-ras is an essential gene, but the defects apparent in the transgenic mice were not immediately due to a failure in cell proliferation. One Ki-ras knock-out mouse line suffers from anaemia during embryogenesis and dies between embryonic days 12.5 and 15 (Johnson et a l, 1997). In these embryos Ki-ras appears to be essential only for the generation of haemopoietic cell lines. The second Ki-Ras knock-out m ouse line has different defects - m yocardial development is aberrant and widespread neural apoptosis occurs - but the mice die at about the same stage of embryogenesis (Koera et a l, 1997). The differences between the mice lines may be resolved by closer examination of the defects but both lines dem onstrate that Ki-Ras is not required for general cell proliferation either.
should show embryonic defects associated w ith failure to proliferate, but this is not the case. The reason is probably redundancy between the three mouse Ras proteins w hereby one can substitute for the loss of the other. Similarly, the lack of w idespread defects in proliferation or differentiation - phenotypes that could be expected from previous studies - may be due to redundancy. The N-ras knock-out mice and heterozygote Ki-ras mice were crossed w ith some resulting increase in the severity of the defects (Johnson et a l , 1997). This observation argues strongly for redundancy between the ras genes and suggests that all three genes may need to be targeted in order to determine the true requirem ents for Ras protein function in mice.
1.6. Ras Signalling Pathways.
The majority of work on Ras signalling pathw ays has concentrated on the role of Ras in the control of cellular grow th and proliferation, ras genes are widely m utated in hum an cancers (Bos, 1989) and the concentration of work over almost tw enty years reflects the im portance of Ras as a regulator of grow th. This description of Ras signalling pathways will concentrate initially on the pathw ays that lead to the activation of Ras, from receptor tyrosine kinases and serpentine receptors, and then will describe the immediate effectors of active Ras and the downstream pathways from these effectors.
1.6.1. Activation of Ras.
The description of the common pathw ays that leads to Ras activation will proceed in reverse order, from the proteins that perform the actual activation process - the RasGEFs - to the mechanism by which they are themselves activated, and eventually to the growth factors and other extracellular signals that initiate Ras activation.
I.6.I.I. RasGEFs
instance to be a component of the R as/adenylate cyclase pathway, and likely to act as an activator of Ras (Broek et a l, 1987; Robinson et a l, 1987). Subsequently by in vitro studies it was dem onstrated to be an exchange factor for Ras (Jones et a l, 1991). CDC25p has since become the prototypical member of the RasGEF family and the tem perature sensitive cdc25-l strain, which suffers a grow th arrest at the restrictive temperature, has been used for the cloning of novel RasGEFs from other species (Martegani et a l, 1992; Goldberg et a l, 1993). The cdc25- line is also used commonly to determine if a putative GEF has RasGEF activity (Liu et a l , 1993a; Coccetti et a l, 1995). CDC25p is a large protein (1589 amino acids) but the minimal dom ain required for exchange factor activity is much smaller (approximately 450 amino acids; Lai et a l, 1993). This domain is highly conserved in RasGEFs from all species.
The best characterised RasGEFs in higher eukaryotic cells belong to members of the Son of Sevenless (Sos) Family. The original member of this family was identified, along w ith a Drosophila Ras protein, as a com ponent of the signalling pathw ay that controls the differentiation of the R7 photoreceptor cell in the Drosophila com pound eye and which lies dow nstream of the Sevenless receptor (Sev; (Rogge et a l, 1991)). The R7 photoreceptor cell differentiation is induced by the neighbouring R8 cell in each ommatidium by the action of the transmembrane Bride of Sevenless protein (Boss) th at is expressed in the region of plasm a membrane adjoining the R7 cell. The receptor for Boss is the Sevenless receptor (Sev), a transmembrane tyrosine kinase receptor that shares homology w ith growth factor receptors (Hafen et a l, 1987).
Loss of function mutations in sev prevent the R7 cell in each om m atidium from differentiating into a photoreceptor whereas gain of function m utants cause aU Sev- expressing cells to adopt the R7 fate (Basler et a l, 1991). Sos and Ras were both shown to suppress loss of function sev mutations, placing them downstream of Sev in the genetic pathw ay (Rogge et a l, 1991; Simon et a l, 1991). The cloning of Sos then revealed strong homology w ith CDC25p from S. cerevisiae and suggested a link between Ras and tyrosine kinase receptors (Bonfini et a l, 1992). Multiple Sos homologues have since been identified in mammalian species.
increase in the am ount of Sos found associated w ith membranes after treatm ent of cells w ith grow th factors, a treatm ent th a t leads to Ras activation. M ost convincingly, Sos or CDC25 proteins targeted to the plasma membrane by fusing a Ras CAAX-box to the C-terminus constitutively activated Ras in mammalian cells in the absence of growth factors (Aronheim et al., 1994) or in yeast cells (Quilliam et al., 1994). Membrane recruitment is now considered to be a universal mechanism for the activation of RasGEFs.
I.6.I.2. Adapter Proteins.
Sos is usually found in the cytosol in association w ith a second protein, an adapter protein, which facilitates the membrane recruitm ent of the GEF. The first adapter protein to be identified was the product of the sem-5 gene of C. elegans as a result of its involvem ent in the differentiation of the vulva (Clark et a l, 1992). Vulval development, like the differentiation of the Drosophila photoreceptors is a process dependent on an induction event. Three vulval precursor cells are induced to differentiate into vulval cells by an epidermal growth factor - like (EGF) signal from the anchor cell in the somatic gonad. Failure to differentiate into vulval cells, for example in the absence of the inductive signal, results in the adoption of an epiderm al fate (Alberts et al, 1994). The C. elegans ras homologue (encoded by the let-60 gene) regulates this induction pathway. Activated forms of Let-60 lead to a m ultivulval phenotype where excess differentiation into the vulval cell fate occurs. In contrast, inhibitory Let-60 forms produce a vulvaless phenotype (Beitel et ah, 1990; H an and Sternberg, 1990). Sem-5 was placed dow nstream of the Let-23 receptor but upstream of Ras in this pathway (Clark et a l, 1992).
The highly homologous drk (downstream of receptor kinase) and grb l (growth factor receptor binding) genes were cloned from Drosophila and hum an expression libraries respectively by screening w ith the tyrosine phosphorylated intracellular taü of the hum an EGF receptor (Lowenstein et a l, 1992; Olivier et a l, 1993; Simon et a l, 1993). The high degree of homology of the Grb2 and Drk proteins to Sem-5 immediately suggested a link between the activation of tyrosine kinase receptors and the activation of Ras, and indeed the conservation between the proteins is such that both Grb2 and Drk can functionally substitute for Sem-5 (Stem et a l, 1993).
The adapter proteins are simple m odular proteins w ith no apparent catalytic activity consisting of two Src-Homology 2 (SH2) dom ains separated by a Src- Homology 3 (SH3) domain. SH2 domains are independently folding motifs of approxim ately 100 amino acids th at are w idely found in signal transduction
m olecules and which bind w ith high affinity to phospho-tyrosine containing sequences. Tyrosine residues in the intracellular regions of growth factor receptors are commonly autophosphorylated following ligand binding. SH3 dom ains are smaller (approximately 50-60 amino acids in length) and bind to proline rich sequences found in some cytoskeletal proteins and significantly, in the C-terminus of Sos (Pawson and Gish, 1992). It is through the C-terminal dom ain that Sos associates with Grb2.
Grb2 and Sos can be co-immunoprecipitated from the cytosol of quiescent cells and both become associated w ith grow th factor receptors upon stimulation of the cells w ith mitogens (Buday and Downward, 1993). By association w ith an adapter protein, Sos is able to translocate to the membrane in response to the appropriate stimulation where it can activate Ras. More recent studies have indicated that the Grb2-Sos interaction is not a perm anent one and can be regulated as p art of a feedback loop that will limit the extent of Ras activation. Activation of MAP kinase by Ras (see below) leads to the phosphorylation of Sos and the reduction in Sos- m ediated Ras activation. Reports differ on the direct consequences of Sos phosphorylation. Sos either remains associated w ith Grb2 but the complex can no longer bind to phospho-tyrosine sequences in the growth factor receptors, or Sos dissociates from Grb2 (Buday et a l , 1995; Rozakis-Adcock et a l, 1995)[Corbalan- Garcia, 1996 #120].
I.6.I.3. Receptors.
The Let-23 and Sev receptors that bind the inducing signals proteins in C. elegans vulval developm ent and Drosophila eye developm ent respectively are both members of the EGF family of tyrosine kinase receptors (RTKs) (Hafen et a l , 1987; Aroian et a l, 1990). Ras is activated in response to many growth factors in addition to EGF, including platelet-derived grow th factor (PDGF), insulin, and neural grow th factor (NGF). The receptors become activated upon ligand binding w ith resultant dim érisation and cross phosphorylation of tyrosine residues in the intracellular domains. The phospho-tyrosine act as targets for the binding of SH2- containing proteins such as the Grb2 adapter family.
1.6.2. Alternative Pathways for Ras Activation.