Copyright 8 1995 by the Genetics Society of America
Meiotic Instability of 9thium
sylvaticum
as
Demonstrated by Inheritance of
Nuclear Markers and Karyotype Analysis
Frank Martin
Plant Pathology Department, University of Florida, Gainesville, Florida 3261 1-0680
Manuscript received June 30, 1994 Accepted for publication December 6, 1994
ABSTRACT
Progeny from a sexual outcross between opposite mating types of Pythium sylvaticum were analyzed for inheritance of RFLP and random amplified polymorphic DNA (RAF'D) markers. Although most were inherited in expected Mendelian frequencies, several were not. Pulsed field gel electrophoresis was employed to examine these unexpected patterns of marker inheritance at a karyotypic level. Parental oogonial and antheridial isolates had different electrophoretic karyotypes and minimum number of chromosome-sized DNAs (13 and 12, respectively), however, summation of the sizes of all chromosomal bands for each isolate was similar at -37 Mb. Progeny karyotypes differed significantly from each other and the parental isolates, ranging in estimated minimum number of chromosome-sized DNAs from 9 to 13 and the summation of band sizes within each isolate from 28.1 to 39.0 Mb. For the eight isolates most extensively analyzed, 80% of the progeny chromosome-sized DNAs were nonparental in size or hybridization grouping of cDNA clones and isolated RAPD markers. Based on the results of Southern analysis it appears that length mutations and perhaps aneuploidy and translocations have contributed to generation of karyotypic polymorphisms. Nineteen field isolates of P. sylvaticum collected from the same location also exhibited significantly different karyotypes, suggesting that the meiotic instability observed in the laboratory also is occurring in field populations.
T
HE oomycete genus Pythium (family Pythiaceae) is taxonomically and ecologically diverse with>
120described species occupying a range of terrestrial and aquatic habitats (VAN DER PLAATS-NITERINK 1981; DICK
1990). Although the primary economic importance of
members of this genus are as plant pathogens, one spe- cies also is a mammalian pathogen ( DECOCK et al. 1987)
and several species are under evaluation for their pro- duction of human dietary supplements ( GANDHI and WEETE 1991; STINSON et al. 1991). Although the mem- bers of the Pythiaceae are fungal in appearance, phylo- genetic comparisons place them more closely aligned to algae and plants than with the eumycota (KNOLL
1992; BALDAUF and PALMER 1993). Unlike most eumy-
cotan fungi, the genus is diploid throughout its entire life cycle with meiosis occurring in the gametangia be- fore fertilization.
In nature the primary means of survival is the sexual reproductive structure (the oospore), but with some species it also may be the asexual spore (the sporan- gia). Most of the species in the genus are homothallic and readily self with a total of seven species classified as heterothallic. Although it is generally considered that heterothallic species require the presence of an oppo- site mating type (oogonial as the maternal and anther- idial as the paternal parent) to sexually reproduce, ex- ceptions to this have been reported. With some isolates of heterothallic species low levels of homothallic behav-
ior have been observed (VAN DER PLAATS-NITERINK
Genetics 1 3 9 1233-1246 (March, 1995)
1981). The mechanisms responsible for this are not known, although variation in ploidy similar to what has been observed in the related genus Phytophthora (see
BUSIER 1992) or meiotic translocation events are possi- bilities.
and not the generation of chromosome-sized polymor- phism (MARTIN 1995).
The purpose of this investigation was to evaluate the effect of meiosis on the inheritance of specific genetic markers and karyotypic stability. Prior experimentation on the stability of karyotypes following selfing of two homothallic species provided conflicting results (MAR-
TIN 1995). For
P.
ohgundrum, almost all progeny were identical to the parental isolate. The only variation de- tected was a loss of a putative supernumerary chromo- some that encoded sequences duplicated on the largest chromosome. In contrast, although the karyotypes of all progeny from a selfed isolate ofP.
spinosum were nearly identical to the parental isolate, 61% were poly- morphic for at least one chromosome-sized DNA. One possible reason for this instability after meiosis may lie in the observation that the isolate appears to be hetero- zygous for rDNA (MARTIN 1990) and thus, may be ahybrid isolate generated by sexual outcrossing. Al-
though the ability of this species to outcross has not been experimentally tested, outcrossing between iso- lates of another homothallic species ( P . ultimum) has been reported in laboratory crosses (FRANCIS and ST.
To evaluate the effect of sexual outcrossing on the generation of chromosomal polymorphisms, opposite mating types of the heterothallic species P. sylvuticum were crossed and the progeny evaluated for inheritance of genetic markers and electrophoretic karyotype. To follow the inheritance of individual chromosomes, cDNA clones and several random amplified polymor- phic DNA ( RAPD ) markers were used as probes in Southern analysis. The results indicate that outcrossing between isolates with different karyotypes can generate significant levels of polymorphisms in the progeny. The analysis of field isolates collected from the same loca- tion suggests that meiotic instability contributes to intra- specific karyotypic polymorphisms under natural condi- tions.
CLAIR 1993).
MATERIALS AND METHODS
Fungal cultures: Opposite mating types of P. syluaticum were recovered from Iowa (1987-14, an oogonial isolate) and Kentucky ( 1987-73, an antheridial isolate)
.
Both isolates were used previously to investigate mitochondrial inheritance in this species (MARTIN 1989). Both isolates had a strict heter- othallic behavior and did not produce sexual reproductive structures when grown singly in agar or liquid medium (de- scribed below) or in grass blade culture ( MARTIN 1992). Nineteen field isolates of P . syluaticum collected from the Johnston Plant Breeding Station of Pioneer Hybred Interna-tional, Inc. in Johnston, Iowa by D. HAEFELE also were exam- ined by pulsed field electrophoresis. All isolates were recov- ered from infected germinating corn seed and were stored until use as previously described (MARTIN 1995)
.
Crosses and isolation of progeny: Agar transfers of the op- posite mating types were coinoculated into petri plates con- taining half strength V-8 broth (AYES and LUMSDEN 1975)
amended with 5 pg/ p1 cholesterol and incubated at 25" in the dark for 14 days. The contact zone between the two cultures containing the oospores was removed, homogenized in a blender at high speed for 20 sec and subjected to sonification treatment at 200 watts for 30 sec. A modification of the proce- dure of RUBEN et al. (1980) was used to selectively eliminate sporangia; the spore suspension was treated with 0.2% potas- sium permangenate for 20 min and rinsed three times in sterile deionized water prior to plating on oospore germina- tion medium that contained 1.5% corn meal agar (CMA) , 0.01 g soybean lecithin (Sigma Chemical Co., St. Louis, MO) , 0.3% Tween 20 and 100 p g / p l penicillin. Prior investigations demonstrated that this procedure was effective in eliminating the viability of all mycelial fragments and sporangia.
Plates were incubated in continuous light at room tempera- ture, with the initiation of oospore germination beginning after -48 hr. Germinated oospores were visualized under a stereo microscope and transferred to fresh-water agar plates using a fine pointed transfer needle. To eliminate the possibil- ity that multiple germinated oospores might be present in the transfer, single hyphal branches were recovered 12 hr later and transferred to potato dextrose agar slant tubes. After 5 days growth, the cultures were covered with sterile mineral oil and the slants were kept as long-term storage of reference isolates. Mating type of the progeny was determined by pair- ing individually with the parental isolates on CMA supple- mented with 5 pg/ml cholesterol.
DNA isolation: Agar transfers for each isolate were made individually into liquid B medium amended with 100 p g / p l penicillin (MARTIN and KISTLER 1990). After 3 days growth at 25", the contents of two plates were combined, the agar blocks removed and excess medium adsorbed onto paper towels. The mycelium was then placed into a 1.5-ml microfuge tube, frozen and lyophilized. The tissue was disrupted with a wooden stick, 1.0 ml of DNA extraction buffer (M ART IN and
KISTLER 1990) was added and the contents mixed with a pipet
tip before incubation at 65" for 15 min. After the addition of 0.5 ml of 5 M potassium acetate the tubes were mixed and incubated on ice for 20 min and then centrifuged at 4" for 15 min. One milliliter of the supernatant was removed and treated sequentially with RNAse (50 ug/ml) and pronase (200 ug/ml) at 37" for 60 min each. The tubes were extracted once with pheno1:chloroform:isoamyl alcohol (25:24:1) fol- lowed by ch1oroform:isoamyl alcohol (24:l) and the DNA precipitated by mixing 0.75 ml with 75 ul 3 M sodium acetate and 0.7 ml isopropanol. After incubation at room tempera- ture for several minutes, the DNA was pelleted by centrifuga- tion at room temperature. Residual supernatant was removed and the pellet washed with 70% EtOH before drying and resuspending in 50 pl TE buffer. Purified rDNA was separated from nuclear and mitochondrial DNA by cesium chloride- bisbenzimide ultracentrifugation of large scale DNA extrac- tions as previously described (MARTIN 1990) .
Meiotic Instability of Pythium 1235
(Madison, W I ) with cDNA synthesis performed using stan- dard techniques ( SAMBROOK et al. 1989). A Not1 primer-adap tor was used to prime first strand synthesis and all reactions were carried out in DEPGtreated water with RNAsin (Pro- mega). The cDNA was ligated into NotI-EcoRV digested pBluescript vector (Stratagene) and the bacterial host for transformation was DH5 alpha.
Determining inheritance of DNA markers: Polymorphisms in the rDNAwere useful for differentiating between the oppo- site mating types. A MspI polymorphism in the ITS 2 region was characteristic of the oogonial parent (1987-14) whereas a RsaI polymorphism in the IGS region was characteristic of the antheridial parent ( 1987-73). Restriction digests, elec- trophoretic separations, and Southern analysis of genomic DNA to identify these polymorphisms were performed as pre- viously described (MARTIN 1991 )
.
Hybridization probes were cloned fragments of the rDNA repeat unit of the oogonial parent spanning the variable regions (MARTIN 1990).
RAPD analysis was done using the procedure of WILLIAMS et al. ( 1990). Primers were obtained from Operon Technolo- gies (Alameda, CA) and were used at a final concentration of 0.4 p ~ . Amplifications were conducted in a total volume of 25 pl, which contained 2.5 pl 1 O X buffer; 100 p~ dATP, DCTP, dGTP, and dTTP; 3 mM MgClp; and 100 ng of fungal DNA. One unit of Tuq polymerase (Promega, Madison, W I ) was added in a volume of 5 p1, and the tubes were layered with mineral oil before placing in the thermocycler (Temp- cycler 11, Coy Laboratory Products, Ann Arbor, M I ) . The cy- cling parameters for amplification were 1 min denaturation at 95" followed by 35 cycles of 1 min annealing at 35", 2 min extension at 72" and 1 min denaturation at 94.5'. The final cycle had an extension time of 10 min at 72". Ramping times between all temperatures were set at the fastest settings for the machine. Amplified products were electrophoretically separated on 2-396 agarose for scoring of polymorphic bands. Parental isolates were included in all amplifications as positive controls and amplifications were repeated if spurious results were observed. Amplifications for progeny isolates that lacked specific bands were repeated to ensure that the results were valid. Only robust and consistently amplified bands were uti- lized for data analysis. The primers that were used, E-1
GACC) , Ell (GTAGACCCGT) , B-12 (CCTTGACGCA) and F-10 (GGAAGCTTGG), generated fragment sizes of 0.47; 0.65; 1.7 and 0.52; 0.52, 0.34, 0.30, 0.27, and 0.23; 0.65 and 0.62; and 0.56 kb, respectively.
Pulsed field gel electrophoresis of intact chromosomal DNA Sample preparation of intact chromosomal DNA and pulsed field gel electrophoresis (PFGE) were done as pre- viously described (MARTIN 1995) . A Bio Rad CHEF I1 electro- phoresis apparatus was maintained in a cold room at 4" and run with 0.5X TBE running buffer (0.045 M Tris-borate, 0.001
M EDTA, pH 8.0). Agarose gels were made with 1% FMC Fastlane agarose and were run with continuously ramped switching intervals for the length of time indicated in the figure legends. The data in Table 2 was collected from multi- ple gel runs where the switching intervals were varied to more accurately separate the smaller (30-60 min switching), me- dium sized (45-75 min switching) or larger chromosome sizes (50-80 min switching).
(CCCAAGGTC) , E-17 (CTACTGCCGT) , E-20 (AACGGT-
RESULTS
Recovery of progeny: A total of
56
single oospore colonies were recovered, however, some of these were not included in further analysis because of their failureto continue growth when transferred from the germina- tion medium. Because the transfer was done by remov- ing the agar surrounding the germinated oospore, lack of growth was not because of disruption of the fungus. At the time of transfer the germ tubes were
100-300
pm in length and tended to be irregular in growth morphology. After transfer their growth was limited and the hyphae lysed after 4-5 days incubation. These colo- nies are referred to as abortive germinants. Mating type segregated in the progeny in a3:l
ratio (antheridial to oogonial) with10%
of the progeny producing low amounts of oospores in single grass blade culture.Marker inheritance: Restriction fragment length polymorphisms (RFLPs) in the ITS
2
and IGS regions of the nuclear encoded rDNA from the oogonial and antheridial parent, respectively, could be used to differ- entiate between parents. All 51 of the F1 progeny exam- ined were heterozygous for both parental polymor- phisms, indicating that the progeny were the result of sexual outcrossing and not selfing (Table1).
A total of six RAF'D primers were used that generated 12 poly- morphic bands that were strongly and consistently am- plified. While many of the parental markers were pres- ent in the progeny, some were not (Table1).
For example, RAPDsE-17
and B-12-1 were inherited as if
they were homozygous in the parental isolates because there was no segregation in the progeny. Although markers from primersE-1, E-20-1, E-20-2
andE
1
1
-1 behaved in a similar fashion, there were from one to three isolates that did not have the marker present (different isolates for each marker)
.
Segregation ratios of1:l were observed for several markers; this is sugges-
tive that the parental isolate was heterozygous at these loci. Perhaps the most aberrant segregation ratios were obtained for B-11-2, which was inherited in an approxi- mate1:3 ratio. This segregation ratio is unexpected
regardless of whether the marker was homo- or hetero- zygous in the parent. Although a similar segregation ratio was observed for El
1-5,
a marker that was present in both parental isolates, this could be obtained if both of the parents were heterozygous.Electrophoretic karyotype: The oogonial and an- theridial parents contained a minimum of
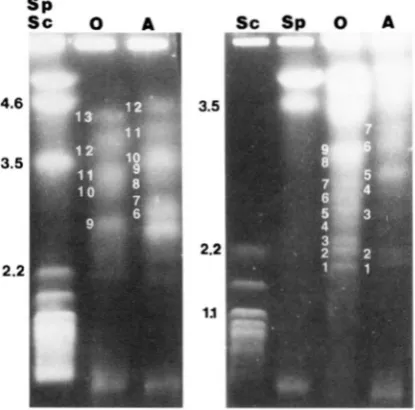
13
and12
chromosomal-sized DNAs, respectively; however it is possible that some bands may have been doublets be- cause their staining intensity varied (Figurel).
At- tempts to clarify the actual chromosome numbers by probing genomic Southerns with cloned telomeric se- quences from another fungus (Fusarium oxysporum,POWELL and KISTLER
1990)
was unsuccessful due to the4.t
3.5
2.2
TABLE 1
Inheritance of molecular markers in the progeny of a cross between opposite mating types of Pythium sylvatimm
Parents" Parental F, segregation'
nuclear
Marker 0 A condition" Observed Expected
rDNA ITS
+
- Ho .5l:O 1 :0rDNA IGS -
+
H o 51:O 1 :0RAPD Primer E- 1
E-1 7
+
E-20-1 -
+
Ho 42: 1 1:on
E-20-2
+
E 1 1-1 -
+
Ho 46: 1 1:on
-
+
Ho 43: 1 19'- Ho 48:O 1
:o
- Ho 4.53 1:W'
Rl l-2
-
+
He 11:36 1:l"Ho 1:Od
E 1 1-3
+
He 26:21 1:lR11-4 -
+
He 23:24 1:lB-11-5
+
+
Ho 36: 1 1 1 :OnHe (both) 3: 1
R12-1
+
- Ho 48:O 1:oR12-2 -
+
He 27:21 1:lF-l 0
+
- He 26:2 1 1:l-
" 0, oogonial, A, antheridial.
I, Suspected nuclear condition in parental isolates based on segregation ratios. Ho, homozygous; He, heterozy-
' Presence:absence of the marker.
'Chi-square analysis indicated that observed values were significantly different from expected ratios ( P
<
gous.
0.001).
S
S
I
FIGURE 1 .-Electrophoretic karyotype for parental oogo- nial ( 0 ) and antheridial ( A ) isolates separated with a ramped switching interval of 5O-80 min and run time of 345 hr (left) or 30-60 min and run time of 348 hr (right). Molecular size markers in the left lanes are Scllizosaccllarom?cps pomfw and Saccharomyces cmmisinr and represent Mb.
parent, respectively. Whether these values reflect the actual genome size of the isolates cannot be conclu- sively stated due to the inability to accurately differenti- ate doublet hands or homologous chromosomes dif- fering in size.
Meiotic Instability of F'ythium 1237
TABLE 2
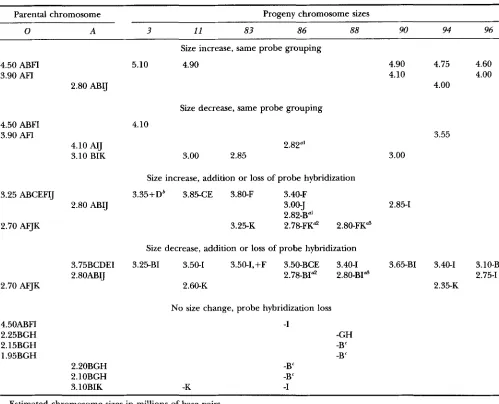
Molecular sizes of chromosome sized bands from CHEF electrophoresis of parental and FI progeny isolates of 4thium sylvaticum
Parental F, Progeny
0 A 3 11 8 3 86 88 90 94 96
4.50 ABFI" 4.90 BF 5.10 ABF 4.90 ABFI 4.50 ABFI 4.50 ABF 4.50 ABFl 4.90 ABFl 4.75 ABFI 4.60 ABFI 3.90 AFI 4.10 AIJ 4.10 ABF 4.20 3.80 ABCEIJ 3.90 AFI 3.90 AFI 4.10 AFI 4.00 AB11 4.00 AFI 3.40 DEF 3.75 BCDEI 3.35 ABCDEF 3.85 ABFIJ 3.50 BCDEF 3.50 DI 3.55 AFIJ 3.65 CDE X.59 AFI 3.75 AH 3.25 ABCEFIJ 3.50 3.25 CDE 3.50 BCDE 3.25 AFJ 3.40 ABCEIJ 3.40 BCDE 3.40 CEI 3.40 BCDE 3.40 ADEj 3.00 B1 3.35 K 3.10 A 3.35 CE 3.00 BIK 3.10 BK 3.10 BIK 3.10 BI 3.10 B 3.10 CDEI 2.80 K 3.10 BIK 2.85 B 3.10 BI 2.85 B I K 3.00 ABI 2.85 BK 3.00 B I K 3.00 I K 3.00 BK 2.70 AFJK 3.00 BI 2.80 B 3.00 BIK 2.55 2.82 AIJ 2.80 A1 2.85 AI31 2.80 K 2.80 IK
2.48 2.55 2.45 2.65 R 2.20 BGH 2.55 B 2.50 2.50 B 2.39 AFJ 2.70
2.35 2.45 2.25 2.60 AFJ 2.45 J 2.45 2.35 2.15 BGH 2.46
2.25 BGH 2.20 BGH 2.15 BGH 2.40 2.35 2.2.5 B 2.10 BGH 2.43
2.15 BGH 2.10 BGH 1.95 BGH 2.15 BGH 2.20 GH 2.15 GH 1.85 GH 2.10 RGH
1.95 BGH 2.10 GH 1.95 GH 1.90 GH
37.25 Mb 37.80 Mb 35.85 Mh 38.50 Mb 28.10 Mb 38.65 Mb 38.10 Mb 36.55 Mh 31.65 Mh 39.00 Mb
2.52 B 2.80 ABIJ 2.50 B 2.80 BK 2.45 2.78 A 1 2.70 2.75 ARFIJK 2.55 B 2.75 AI31
Sizes are in millions of base pairs and were estimated based on comparative migration with Schizosncchnrom~lrrs p o m b and
Saccharomyces cerevisiae.
" Letters represent the hybridization of different cDNA clones or isolated RAPDs band to specific chromosome sized DNAs. A, rDNA; B, pC1-34; C, pC2-10; D, pC2-19; E, pC2-24; F, RAPDs E17-1; G, pC1-17; H , pC1-61; I, pC1-18; J, pC1-1; K, RAPDs B-11-1 (location on isolate 3 not known for probes I, J and K).
lar in size to the parental chromosomes, however, some were unique (Table 2 )
.
Regression analysis of chromo- some numbers and summation of their sizes in progeny isolates yielded a linear relationship ( R2 = 0.95, P = 0.00004), indicating that differences in chromosomal numbers rather than significant changes in chromo- some size were predominantly responsible for the ob- served variation.Southern analysis of pulsed field gels: To determine if similar-sized chromosome bands in the parental and progeny isolates were homologous and to clarify the events associated with generating the polymorphisms,
VJ
+
+
4.6-
1.6-
i
'
FIGURE 2.-Electrophoretic karyotype for parental oogo- nial ( 0 ) , antheridial ( A ) , and FI progeny isolates of Fythium sylvaticum (numbered lanes) separated with a ramped switch- ing interval of 45-80 min and run time of 440 hr. Far left are molecular-sized markers Schizosaccharomyces pombe and Sac- charomyces c e r e v i s i a e and represent Mb.
Southern transfers of the PFGE gels were probed with coding regions (rDNA and cDNA clones) and two
RAPD bands. Additional RAPD bands were evaluated for use as probes, however, they were not included in the results as they represented repetitive DNA and were present on every chromosome. Genes for rDNA were present on four and two chromosomes for the oogonial and antheridial parents, respectively. The FI progeny had mixtures of these parental patterns as well as non- parental chromosomal sizes (Figure 3A, Table 2 )
.
For instance, isolate 11 had two rDNA encoding bands that were similar in size to the parents as well as a lower sized band that was not present in either parent. Similar results also were observed when probing with cDNA clones (Figure 3, B-D, Table2 ) .
All probes that have been examined hybridized to the smallest diffuse band at the bottom of each lane, this is believed to reflect the presence of degraded DNA generated during the preparation of the protoplast insert plugs.The chromosomal locations of a total of nine coding regions and three RAPD bands have been placed (Ta- ble 2 ) . RAF'D marker E 1 1 -2 had the same hybridiza- tion grouping as E1 1
-
1 ( data not shown ).
From com- parisons of chromosome sizes and location of these clones, it is apparent that the progeny vary from the parental isolates in several respects. One readily appar- ent difference is that similar sized chromosomes do not always encode the same grouping of clones. For example, in the antheridial isolate the 3.50"b band does not encode any of the clones evaluated, yet inv) +
s
3.5
3.5
2.2
8
22-
1 .
-
FIGURE 3.-Southern transfers of PFGE-separated chromosomes probed with rDNA (A; 40- to 70-min switching interval, 390
hr) or cDNA clones pC2-19 (B; 40- to 70-min switching interval, 390 h r ) , pC2-24 (C; 45- to 75-min switching interval, 400 hr) and pC1-34 ( D ; 45- to 75min switching interval, 408 hr)
.
Molecular-sized markers in the left lane for panel ( A ) are from Schizosaccharomyces pomhe and Snccharomym cprpoisinr and represent Mb.Mb chromosome in the antheridial isolate encoded markers ABIJ whereas in isolate 11 it encoded only BK. Similar comparisons may be observed among other chromosomal bands as well. Additional differences among progeny and parental isolates were associated with length mutations (either with or without differ- ences in probe hybridization grouping) or changes in probe grouping without detectable length mutations
(Table 3 ) .
In addition to size differences, some progeny con- tained chromosome bands with nonparental combina- tions of coding regions. In isolate 83 the 3.50-Mb chro- mosome encodes clones BCDEF and in isolate 96 the 3.40-Mb chromosome encodes A D E J , combinations that are not found in any parental chromosome. Al- though this is suggestive of translocations, chromosome mapping needs to be conducted to ensure that it does not represent comigration of nonhomologous chromo- somes.
Analysis of the progeny also suggests that aneuploids are present. After meiosis, each gametangium should contribute a haploid complement of chromosomes to the oospore, which would then pair with its homolog to form a diploid nucleus. If these homologs were the same size, only a single band encoding sequences from
Meiotic Instability of Pythium 1239
TABLE 3
Changes in size or groupings of probe hybridization on chromosome sued DNAs in parental isolates of qthium sylvaticum
compared with progeny isolates
Parental chromosome Progeny chromosome sizes
0 A 3 11 83 86 88 90 94 96
4.50 ABFI 3.90 AFI
4.50 ABFI 3.90 AFI
3.25 ABCEFIJ
2.70 AFJK
2.70 AFJK
4.50ABFI 2.25BGH 2.15BGH 1.95BGH
2.80
ABIJ
4.10 AIJ 3.10 BIK
2.80 ABIJ
3.75BCDEI 2.80ABIJ
2.20BGH 2.10BGH 3.1OBIK
Size increase, same probe grouping
5.10 4.90
Size decrease, same probe grouping
4.10
2.82"'
3.00 2.85
Size increase, addition or loss of probe hybridization
3.35+Db 3.85-CE 3.80-F 3.40-F
3.00-J 2.82-B"' 3.25-K 2.78-FKd 2.80-FKa
Size decrease, addition or loss of probe hybridization
3.25-BI 3.50-1 3.50-I,+F 3.50-BCE 3.40-1 2.78-BId 2.80-BI" 2.60-K
No size change, probe hybridization loss
-I
-GH -B" -B" -B"
-B"
-K -I
4.90 4.10
3.00
2.85-1
3.65-BI
4.75 4.60 4.00 4.00
3.55
3.40-1 3.10-B
2.75-1 2.35-K
Estimated chromosome sizes in millions of base pairs.
a Alternative explanations for generation of this chromosome are marked with a similar superscript notation.
bThis culture was lost prior to placement of clones H, I and J.
"This might also represent a deletion of a larger chromosome from the parental isolates with a size range of 2.15-2.25 Mb.
size and probe hybridization as the oogonial parent and the closest possible homolog was 0.70 Mb smaller.
An alternative explanation to aneuploidy for these results is that the different bands represented different sized homologs in which translocation events altered the probe grouping. Different-sized homologs have been observed in the diploid Candida albicans, although the size differences between homologs were smaller than noted above. Comparison of the results of THRASH-
BINGHAM and GORMAN (1992) and IWAGUCHI et al.
( 1992) indicate that homologs in the size range of 1.2
Mb differ by -100-200 kb; similar results also were observed in comparisons between larger dissimilar sized homologs as well. However, greater size differences
were observed for the diploid protozoan Trypanosoma
brucei;
in which homologous pairs differed in size by asmuch as 20% ( GOTTESDIENER et ai. 1990)
.
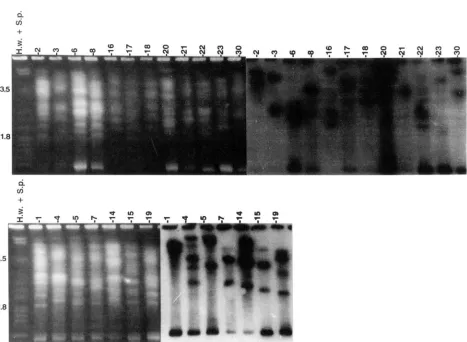
Karyotype of field isolates: Seven oogonial and
12
antheridial mating type isolates collected from the same research farm exhibited different electrophoretickaryotypes; no two isolates were the same (Figure 4 ) .
v)
+
4
v)
+
Q
FIGURE 4.-Electrophoretic karyotype of field isolates of Qthium sylunticum collected from the same farm in Johnston, Iowa, separated with a ramped switching interval of 55-90 min and run time of 425 hr. Antheridial mating types are in the top panel and oogonial in the bottom, Southern transfer on the right is for rDNA. Molecular-sized markers in the left lane are Schizo.tacchnro- mvws p o m k and Hansmula wingk and represent Mb.
bands depending on the field isolate, none of which were similar for any of the isolates. Two isolates ( 4 and
7 )
contained a chromosome < 1 Mb, an unusual size for this species ( M A R T I N 1995).DISCUSSION
Marker inheritance: Inheritance of both parental rDNA RFLPs in all F, isolates confirmed the biparental nature of the progeny. With a few exceptions, similar results also were observed for inheritance of
RAPD
markers that were believed to be homozygous in the parent. Nonmendelian inheritance was observed with four of these markers, however, the frequency was low and represented 2.2-6.3% of the progeny isolates. The greatest deviation from expected patterns of inheri- tance was for marker B-11-2; the observed 1:3 ratio does not correspond to ratios expected for homo- orheterozygosity in the parent. One interesting feature for this and the other four B-11 markers is that in South- ern analysis of PstI- or XhoIdigested genomic DNA from either parent, all marker hands hybridized to a
single 1.7-kb band. When used as a probe of PFGE gels,
B-1 1-2 also had the same chromosomal location as E1 1-1. One interpretation of these results is that the amplified sequences with this primer are part of a repet- itive element and that the DNA fragment for B-11-2
may be more prone to recombination than the other markers.
Based on segregation ratios and the observation that mitochondrial DNA is maternally inherited (MARTIN 1989), most RAF'D markers appear to be nuclear in origin. The only possible exception is for E-17 and
B-12-1, both of which are present in the oogonial par- ent and did not segregate in the F,. Evaluation of nu- clear or cytoplasmic location of these markers will have to be determined by analysis of backcross progeny.
Similar variation in inheritance of nuclear markers has been reported in crosses of other oomycetes. FOR-
STER and COFFEY (1990) identified nuclear encoded RFLP markers that were homozygous in parental strains of Phytophthuru purusiticu, but segregated in a nonmen- dellian fashion in 41 % of the progeny isolates. Because
Meiotic Instability of Pythium 1241
from sexual outcrossing rather than induced selfing. The explanation that was proposed for the aberrant patterns of marker inheritance was meiotic nondisjunc- tion leading to the creation of aneuploid progeny. How- ever, because this could not be confirmed by linkage analysis, translocations or deletion events as observed with P . sylvuticum in this study also are possible explana- tions. Similar aberrant patterns of inheritance were ob- served for isozyme loci in backcross and sibling crosses for Phytophthoru infestans ( SPIELMAN et al. 1990) and for some hybridization bands for the repetitive element
RG57 in genomic Southerns of F1 progeny ( GOODWIN
et al. 1992). In contrast, HULBERT et al. ( 1988) observed that with a few exceptions, most RFLP marker loci were inherited in the expected Mendelian ratios for crosses with Bremia lactucae, another oomycete but in a different family. In one cross two of 44 loci segregated as 11:27 and 28:lO instead of 1:l and in another cross a total of six loci behaved in a similar fashion. One individual from these crosses was homozygous for two loci when heterozygosity was expected. Due to the inheritance of other markers the mechanism responsible for this was thought to be a translocation event rather than aneu- ploidy. Using different markers, FRANCIS and MICHEG
MORE (1993) observed that loci present on the smaller chromosome size ranging from 0.3 to 1.6 Mb were the only ones that did not segregate in a Mendelian fashion, which was consistent with the inheritance of the chro- mosomes as observed by PFGE.
It should be noted that the parental and progeny isolates in this investigation were the same that were used to examine mitochondrial inheritance in a sexual outcross for this species (MARTIN 1989). The inheri-
tance of nuclear markers confirms that these isolates were the result of outcrosses and not selfings and sup- port the conclusions presented that, within the limits of detection used, mitochondria were uniparentally in- herited and contributed by the maternal parent. These results are in contrast to what has been observed with Phytophthora spp., where opposite mating types are bisexual (capable of maternal or paternal behavior) as well as capable of induced selfing (reviewed in BRASIER
1992).
Electrophoretic karyotype: Significant polymorphisms were observed in the electrophoretic karyotypes of the two parental isolates and their progenies. Although the oogonial and antheridial isolates had similar values for summation of chromosomal band sizes of -37 Mb, the numbers of chromosome-sized DNAs (13 and 12, re- spectively) and karyotypes were different. These differ- ences also were reflected in the karyotype of progeny isolates, where numbers of chromosomal-sized DNAs ranged from 9 to 13 with many of their sizes not only different from the parents, but also from each other. The polymorphisms were so extensive that no two of the 15 progeny isolates examined (data for 4 not shown
because of incomplete Southern analysis data) had the same karyotype. Summation of estimated chromosome band sizes for the progeny also exhibited significant variation and ranged from 28.1 to 39.0 Mb. Due to the linear relationship between chromosome number and summation of band sizes, it appears that differences in numbers of chromosomal bands are responsible for this difference rather than chromosomal recombination generating significantly different-sized chromosomes.
Comparison of staining intensities of chromosomal bands revealed that different amounts of DNA are pres- ent, therefore summation of calculated sizes of individ- ual chromosome-sized DNAs has several limitations for determining genome sizes and may represent an under- estimation of the actual values. This approach does not take into consideration the possibility that heterologous chromosomes may migrate with the same apparent mo- lecular size generating a doublet band. Conversely, be- cause Pythium spp. are diploid, it is possible that differ- ent chromosome bands may actually be size variants of a homologous pair. This has been observed in other diploids such as T. brucei ( GOTTESDIENER 1990) and C.
albicans (MAGEE et al. 1988; IWAGUCHI et al. 1990;
THRASH-BINNGHAM and GORMAN 1992)
,
as well as in the haploid fungus A . bisporus when the mycelium is dikaryotic ( LODDER et al. 1993). Although differences in band-staining intensity may be an indication that one of the above events has occurred, accurate quantifica- tion may be difficult because of the diffuse banding patterns and similar rates of migration in the pulsed field gels of the medium to large chromosome-sized DNAs. For the above reasons, scanning densitometry of gel negatives to obtain a comparative estimate of DNA amounts in each band was not attempted in this study. Therefore, correlating counts of chromosomal-sizedDNAs with chromosome numbers, as well as attempting to estimate genome size by summation of band sizes should be approached with caution. To obtain a more accurate genome size estimate, genome reconstruction should be utilized ( HUDSPETH et al. 1977; FRANCIS et al.
1990;
hlAo
and TYL,ER 1991).possibility that the bands represent similar-sized heter- ologous chromosomes are needed. Although not con- tributing to polymorphisms with parental isolates, some progeny chromosomes appear to be the result of aneu- ploidy. The recovery of homothallic progeny from strictly heterothallic parental isolates also is suggestive that translocations or variation in progeny ploidy has occurred.
The extent of polymorphisms of chromosomal-sized DNAs generated by meiosis in this species is quite strik- ing. With a total of 94 chromosomal-sized DNAs identi- fied in the eight progeny isolates examined,
73
( 80.2% ) were polymorphic when compared with the parental isolates for size or clustering of coding regions. In view of the polymorphisms that were associated with chro- mosomes encoding rDNA in other fungi (Neurospora crassa, BUTLER and METZENBERG 1989; Coprinus cinereus, PUKKILA and SKRZYNIA 1993; Kluveromyces lactis, MALES ZKA and CLARK-WALKER 1989; Candida albicans, IWA- GUCHI et al. 1992; RUSTCHENKO-BULGAC et al. 1993; S. cerevisiae and S. pombe, PASERO and MARILLY 1993) the chromosomal locations of rDNA in relation to polymor- phisms in P. sylvaticum progeny was investigated. Al- though rDNA was encoded on four and two chromo- somal bands in the oogonial and antheridial parent, respectively, it is located on from two to five chromo- somes in the progeny for a total of37
chromosomal bands for all eight isolates; 89% of which were nonpa- rental. In total, 35% of all chromosomal bands in the progeny were represented by rDNA encoding chromo- somes that were nonparental. To gauge the significance of this compared wtih other repetitive coding regions, the chromosomal location of cDNA clone pC1-34(clone B in Table 2 ) was evaluated in all isolates. The parental isolates each contained seven chromosomal bands encoding this clone whereas the progeny con- tained from five to eight. Of the total of 65 chromo- somal bands in the progeny that should have had the clone present, 83% were nonparental. Because some of the polymorphic chromosomes encoded both rDNA and pC1-34 ( 3 total in parental isolates), progeny chromosomal bands with both coding regions were eliminated from the analysis and only those encoding pC1-34 were examined;
78%
were nonparental. Simi- lar results were obtained with probe pC2-19 (clone D )that was present on a single chromosomal band in each parent. For the eight progeny examined, the cDNA probe hybridized to 10 chromosomal bands, all of which were nonparental for size or probe grouping. Although the accuracy of these comparisons is limited by the inability to separate nonhomologous comigrat- ing bands and include them in the analysis, the results suggest that for P. sylvaticum, encoding rDNA does not predispose a chromosome to higher rates of meiotic instability than other chromosomes.
These findings do not rule out the possibility that
alteration in the copy number of rDNA repeat units may contribute to variation in rDNA encoding chromo- somal sizes after meiosis. This has been observed in a
number of eumycotan fungi (BUTLER and METZENBERG
1989; IWAGUCHI et al. 1992; MALESZKA and CLARK-
WALKER 1993; PASERO and MARILLY 1993; PUKKILA and
SKFUYNIA 1993; RUSTCHENKO-BULGAC et al. 1993) and may be occurring in P. sylvaticum as well. However, due to the larger sizes of the chromosomal bands in ques- tion this event may not be apparent unless the deletions were significant. In C. cinerea the numbers of rDNA cistrons also was observed to be mitotically unstable
(
P U K K I ~
and SKRZWIA 1993). Although mitotic insta- bility was not extensively examined in the current inves- tigation, size variation of the rDNA encoding chromo- somes was not observed in any of the insert sample preparations made over a 3-yr period. In addition, mi- totic instability of karyotypes was not observed by PFGE when another species, P. myriotylum, was transferred on a weekly basis for one year (MARTIN 1995).
Similar to what has been observed with P. sylvaticum,
ZOLAN et al. (1994) also observed that sexual outcross- ing can occur between strains of C. cinereus with differ- ent karyotypes and that chromosomal polymorphisms are common in the progeny. This generation of poly- morphisms was not observed when strains with the same karyotype were crossed. Chromosome polymorphisms following meiosis also have been identified in S. cerevis- iae ( ONO and ISHINARO 1988)
,
Ustilago maydis ( KINSCH- ERF and LEONG 1988), Lqbtosphan'a maculans ( PLUMMER and HOWLETI 1993) and the diploid protozoan malar- ial parasite Plasmodium falciparum ( WALLIKER et al., 1987). The contribution of aneuploidy to chromo- somal polymorphisms following meiosis has been dis- cussed for the haploid Tilletia caries/controversa by RUSSELL and MILLS ( 1993) and Yawowia lipolytica by
NAUMOVA et al. ( 1993)
.
The reasons underlying the high frequency of mei- otic rearrangements in P. syluaticum have yet to be con- clusively determined, although one possibility is the presence of repetitive DNA. Although the frequency of repetitive sequences in the genome of P. sylvaticum has not been determined, hybridization of several RAPD markers and cDNA clones to multiple bands following Southern analysis of pulsed field gels as well as genomic digests is suggestive that they are not uncommon. There is precedence for repetitive DNA representing a sig- nificant portion of the genome in other oomycetes. For Phytophthora megasperma fsp. glycinea, which also is in the Pythiaceae, MAO and TYLER (1991 ) observed that 53% of the genome was comprised of repetitive DNA. Similar results (65% repetitive sequences) also were obtained for Bremia lactucae (FRANCIS et al. 1990) al- though the oomycete AchZya bisexualis had only 20% repetitive sequences ( HUDSPETH et al. 1977)
.
Meiotic Instability of Pythium 1243
gested to be responsible for the high translocation fre- quencies observed in the diploid yeast,
C.
albicans( THRASH-BINGHAM and GORMAN 1993). Several differ- ent families of repetitive sequences have been identified in this fungus that are either dispersed throughout the genome ( SOLL et al. 1987; CUTLER et al. 1988; SCHERER and STEVENS 1988; LASKER et al. 1991; SADHU et al. 1991 ) or are localized on a specific chromosome ( IWAGUCHI et al. 1992). There appears to be a positive correlation between the chromosomal location of some of these repeated sequences and propensity for undergoing re- arrangements. For example, the absence of these se- quences on chromosome 5 has been suggested as the reason for its conservation in size among different iso- lates ( IWAGUCHI et al. 1992; THRASH-BINGHAM and GOR-
MAN 1993). Because this organism has no known sexual
stage, it is believed that mitotic translocations between dispersed repetitive elements are responsible for chro-
mosomal polymorphisms ( THRASH-BINGHAM and GOR-
MAN 1992). Dispersed repetitive elements also have
been identified on different chromosomes in
C.
cinereus( ZOLAN et al. 1992) , and it has been suggested that they may contribute to chromosome length polymorphisms obtained when parental isolates differing in karyotypes are crossed ( ZOLAN et al. 1994). Recombination be- tween dispersed repeated elements also has been impli- cated in generating polymorphisms in Drosophila melano- gaster (MONTGOMERY et al. 1991) and Plasmodium falciparum ( CORCORAN et al. 1988)
.
When examining a diploid organism such as Pythium spp. or
C.
albicans, different sized chromosomal bands on pulsed field gels may not necessarily represent heter- ologous chromosomes. In C. albicans, the clustering of coding regions on specific chromosomes stays constant, so homologs can be identified even if the chromosomes have different sizes. Based on Southern analysis this species has been determined to have eight chromo- somes, however, additional bands have been identified in some isolates ( MAGEE 1988; IWAGUCHI et nl. 1990; THRASH-BINGHAM and GORMAN 1992).
These bands represent different sized homologs that migrate as dis- tinctly different bands but tend to be similar in size to each other. For example, the two smallest sized chromo- somes in isolate SC5314 migrated as separate bands, yet contained similar hybridization grouping of probes andwere considered homologous by THRASH-BINGHAM and
GORMAN ( 1992)
.
Although molecular size data were not presented, comparison with isolates also examined by IWAGUCHI et nl. ( 1992) revealed their sizes to be in the range of 1.05 to 1.2 Mb and differ by -100-200 kb. This difference was similar to that observed for other small (third and fourth smallest bands) as well as larger bands that were different sized yet considered homologs. Therefore, although homologs in this genus may migrate as distinct bands, their sizes tend to be relatively similar and appear to differ by only several hundred kilobase.In contrast to the results observed with
C.
albicans, significant variations in the clustering of specific coding regions in the parental as well as progeny isolates of P. sylvaticum were observed, thereby making it difficult to identify different sized homologs. Although some prog- eny chromosomes shared probe groupings similar to each parental isolate, suggesting that they may be differ- ent-sized homologs, the size differences compared to parental bands were greater than those observed for C. albicans. One exception to this may be the smallest bands encoding cDNA clones BGH. In the oogonial and antheridial parents, these coding regions were clus- tered on three and two chromosomes, respectively, that differed by 100-300 kb, a size range also observed inC.
albicans.Species behavior in nature: The progeny evaluated in this investigation are the result of a laboratory cross, which leaves open a question concerning the involve- ment of the observed meiotic instability on genome evolution under natural conditions in the field. This may be particularly important for those progeny that have reduced growth rates or alteration in virulence, factors that are observed in many progeny (F. MARTIN, unpublished data) and may have an influence on the ecological competence of the isolate, thereby contribut- ing to its ability to survive in the field. Comparison of karyotypes for isolates collected from the same research farm revealed that no two isolates had the same karyo- type or chromosomal location encoding rDNA or a cDNA clone. Although this is suggestive that the above described meiotic instability has been occurring in a natural field population by sexual outcrossing, it is not conclusive evidence. An alternative explanation may be that the isolates are a reflection of the natural variation present in the species and that multiple introductions reflecting this range in variation has occurred. How- ever, the results observed with the laboratory cross and the extensive polymorphisms observed in these field isolates compared with other isolates of P. sylvaticum collected from geographically diverse areas (MARTIN
1995) strongly suggest that sexual outcrossing is respon- sible.
Biological significance: The high rates of meiotic in- stability and suspected aneuploidy observed in this cross may be an adaptive process allowing for sexual outcross- ing to occur between isolates with significantly different karyotypes or ploidy. Biologically speaking, it may have a significant impact on the success of sexual outcrossing in this heterothallic species since the opposite mating types examined thus far all have different karyotypes
needed for meiosis, the ability to outcross would in- crease the genetic variability that could be obtained. The ability of a homothallic isolate to outcross in nature is not known, however, this phenomenon has been re- ported in laboratory crosses for the homothallic species
P. ultimum (FRANCIS and ST. CLAIR 1993)
.
Although the potential for outcrossing to occur be- tween isolates of P. sylvaticum with such different karyo- types has advantages, it also may have a detrimental effect on some progeny. As previously noted, 9% of the germinated oospores were abortive germinants, mean- ing that the germ tubes grew with abnormal morphol-
ogy for several hundred microns and stopped. This has been investigated for crosses between other isolates as well, with some of them having higher or lower frequen- cies (F. MARTIN, unpublished data)
.
With the observed frequencies of recombination and suspected aneu- ploidy, it is possible that the genomes of these particular oospores lacked necessary coding regions either through loss of specific regions of chromosomes or en- tire chromosomes. Although endogenous nutrient re- serves present in the oospore may have allowed for ger- mination, once depleted the hyphae may have been no longer able to grow. A similar observation concerning abortive germinants and a hypothesis for their occur- rence also has been suggested for Ph. parusiticu ( FOR-STER and COFFEY 1990). Chromosomal recombination and aneuploidy also may contribute to the variation observed in the numbers of mature oospores produced and the frequency of oospore germination when differ- ent isolates of P. sylvaticum are crossed. Depending on the parental isolates, significant variation in these char- acteristics may be observed (F. MARTIN, unpublished data). PUKKILA (1992) also observed that outcrosses between isolates of C. cinereus with polymorphic karyo- types yielded tetrads with low viability, as did NAUMOVA
et al. ( 1993 ) for Y . lipolytica.
Although P. sylvuticum is a heterothallic species that normally requires opposite mating types to sexually re- produce ( CAMPBELL and HENDRIX 1967), there are sev- eral reports to the contrary. PFWIT and GREEN (1971) observed that a number of the antheridial field isolates they examined were capable of selfing. VAN DER PLATTS
NITERINK (1968) reported a similar finding with the ex- ception that selfing was observed at a lower frequency and that predominantly oogonial isolates were homo- thallic. Although the parental isolates used in this investi- gation were strictly heterothallic, -10% of the progeny exhibited low levels of homothallic behavior. This was a fairly stable trait because most selfed progeny from a homothallic isolate also were homothallic, and they also were capable of outcrossing with heterothallic isolates
(P. GAVINO and F. MARTIN, unpublished data). With the observed frequency of recombination and aneuploidy these may have been generated by recombination events or aneuploidy placing the mating type genes together
in the same nucleus and may be an explanation for the generation of the above noted examples of homothal- lism in field isolates. This would be an adaptive advan- tage for this heterothallic species by allowing meiosis to occur in the absence of opposite mating types.
The influence of meiosis on chromosomal karyotype in pythium: The effects of meiosis on progeny karyo- type were different depending on the species. For the homothallic species P. oligundmm, which tends to repro- duce parthenogenically without antheridial attach- ments, the SI progeny were identical to the parent in all but one isolate (which lacked a putative supernumerary chromosome; F. MARTIN 1995). In contrast, selfed
progeny of the homothallic species P. spinosum were variable and -61% of the progeny had at least one
nonparental chromosome (MARTIN 1995). Although
these polymorphisms were not nearly as extensive as was observed for progeny from sexual outcrossing of P. sylvaticum, similarities in the types of karyotypic differ- ences were observed. One possible explanation for the differences in results between P. oligandrum and P. spino- sum may be that the isolate of P. spinosum arose from a sexual outcross. This supposition is based on the pres- ence of approximately equal amounts of polymorphic rDNA repeat units in a single oospore culture of the isolate that was selfed, suggesting that the isolate was heterozygous for rDNA (MARTIN 1990). Although the ability of P. spinosum to sexual outcross has not been examined experimentally, this phenomenon has been observed for another homothallic species in the genus (FRANCIS and ST. CLAIR 1993)
.
If this assumption is true for P. spinosum, then polymorphisms generated by selfing may have arisen by a similar process as observed in this investigation for outcrossing of P. sylvaticum. The different amounts of polymorphisms observed in the progeny of these crosses may be a reflection of the ability of P. spinosum to readily self and subsequently reduce duplicated sequences or aneuploid chromo- somes with successive generations.The contribution of outcrossing on generation of nonparental chromosomes in subsequent selfed prog- eny was observed in some isolates of P. sylvaticum. As
noted above, 10% of the progeny from the P. sylvaticum
outcross examined in this manuscript were homothallic (similar results also were observed when one of the progeny isolates were backcrossed to a parental iso- late)
.
Karyotype analysis of one of these homothallic isolates and 15 of its selfed progeny revealed significant levels of polymorphisms ( P . GAVINO and F. MARTIN,unpublished data)
.
With the level of meiotic instability observed for outcrossed and selfed progeny of P. sylvati- cum, if sexual outcrossing occurs among homothallic species even on an infrequent basis it may be enough to generate the levels of intraspecific karyotypic varia- tion that have been observed in the genus (MARTIN,Meiotic Instability of F'ythium 1245
tion of the potential for homothallic species to outcross under natural conditions in the field.
The author would like to thank M. ZOLAN for providing a preprint of a manuscript before publication. This work was supported by United States Department of Agriculture grant 90-37151-5428. This is Florida Agricultural Experiment Station Journal Series Number R-03927.
LITERATURE CITED
ADAM, R. D., 1992 Chromosome size variation in Giardia lamblia: the role of rDNA repeats. Nucleic Acids Res. 20: 3057-3061. AYERS, W. A., and R. D. LUMSDEN, 1975 Factors affecting production
and germination of oospores of three F'ythium species. Phytopa-
BALDAUF, S. L., and J. D. PALMER, 1993 Animals and fungi are each other's closest relatives: congruent evidence from multiple p r e teins. Proc. Natl. Acad. Sci. USA 90: 11558-11562.
BRASIER, C. M., 1992 Evolutionary biology of phytophthora. I. Genetic system, sexuality and the generation of variation. Annu. Rev. Phytopathol. 30: 153-171.
BUTLER, D. K., 1992 Ribosomal DNA is a site of chromosome break- age in aneuploid strains of neurospora. Genetics 131: 581-592. BUTLER, D. K., and R. L. METZENBERG, 1989 Premeiotic change of
nucleolus organizer size in Neurospora. Genetics 1 2 2 783-791. BUTLER, D. K., and R. L. METZENBERG, 1990 Expansion and contrac-
tion of the nucleolus organizer region of Neurospora: changes originate in both proximal and distal segments. Genetics 126:
BUTLER, D. K , and R. L. METZENBERG, 1993 Amplification of the nucleolus organizer region during the sexual phase of Neurospora crassa. Chromosoma 1 0 2 519-525.
CAMPBELL, W. A., and F. F. HENDRIX, 1967 A new heterothallic F'yth- ium from Southern United States. Mycologia 59: 274-278. CHOMCZWSKI, P., and N. SACCHI, 1987 Single step method of RNA
isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162: 156-159.
CORCORAN, L. M., J. K. THOMPSON and D. J. KEMP, 1988 Homologous recombination within subtelomeric repeat sequences generates chromosome size polymorphisms in P. falciparum. Cell 53: 807-
813.
CUTLER, J. E., P. M. GLEE and H. L. HORN, 1988 Candida albicans and Candida stellatoidemspecific DNAfragment. J. Clin. Microbiol.
26: 1720-1724.
DECOCK, A. W. A. M., MENDOZA, L., PADHYE, A. A., AJELLO, L., and L. K A U W , 1987 Pythium insidiosumsp. nov., the etiologic agent of pythiosis. J. Clin. Micro. 2 5 344-349.
DICK, M. W., 1990 k2y to Pythium. University of Reading Press, Read- ing, U.K.
FORSTER, H., and M. D. COFFEY, 1990 Mating behavior of Phytophthura parasitica: evidence for sexual recombination in oospores using DNA restriction fragment length polymorphisms as genetic markers. Exp. Mycol. 14: 351-359.
FRANCIS, D. M., and R. W. MICHELMORE, 1993 Two classes of chromo- some-sized molecules are present in Bremia lectucae. Exp. Mycol.
FRANCIS, D. M., and D. A. ST. CLAIR, 1993 Outcrossing in the homo- thallic oomycete, Pythium ultimum, detectedwith molecular mark- ers. Curr. Genet. 24: 100-106.
FRANCIS, D. M., S. H. HULBERT and R W. MICHELMORE, 1990 Genome size and complexity of the obligate fungal pathogen, Bremia lactu- cue. Exp. Mycol. 14: 299-309.
GANDHI, S. R., and J. D. WEETE, 1991 Production of the polyunsatu- rated fatty acids arachidonic acid and eicosapentaenoic acid by the fungus Pythium ultimum. J. Gen. Microbiol. 137: 1825-1830. GOODWIN, S. B., A. DRENTH and W. E. FRY, 1992 Cloning and genetic
analyses of two highly polymorphic, moderately repetitive nu- clear DNAs from Phytophthora infestam. Cum. Genet. 2 2 107-
115.
~ T T E S D I E N E R , IC, J. GARCIA-ANOVEROS, M. (2.5. LEE and L. H. T. VAN
DER PLOEG, 1990 Chromosome organization of the protozoan TryPanosomu brucei. Mol. Cell. Biol. 10: 6079-6083.
thology 6 5 1094-1100.
325-333.
17: 284-300.
HUDSPETH, M. E. S., W. E. TIMBERLAKE and R. B. GOLDBERG, 1977 DNA sequence organization in the water mold Achlya. Proc. Natl. Acad. Sci. USA 74: 4332-4336.
HULBERT, S. H., T. W. I L O ~ , E. J. LEGG, S. E. LINCOLN, E. S. LANDER
et al., 1988 Genetic analysis of the fungus, Bremia lactucae, using restriction fragment length polymorphisms. Genetics 120: 947- 958.
IWAGUCHI, S.-I., M. HOMMA and K. TANAKA, 1990 Variation in the electrophoretic karyotype analyzed by the assignment of DNA probes in Candida albicans. J. Gen. Microbiol. 136: 2433-2442. IWAGUCHI, S. I., M. HOMMA, H. CHIBANA and K. TANAKA, 1992 Isola-
tion and characterization of a repeated sequence ( RPSl ) of Can- dida albicans. J. Gen. Microbiol. 138: 1893-1900.
KINSCHEW, T. G., and S. A. LEONG, 1988 Molecular analysis of the karyotype of Ustilago muydis. Chromosoma 96: 427-433. KISTLER, H. C., and V. P. W. MIAO, 1992 New modes of genetic
change in filamentous fungi. Annu. Rev. Phytopathol. 30: 131- 152.
KNOLL, H. A,, 1992 The early evolution of eukaryotes: a geological perspective. Science 256: 622-627.
LASKER, B. A., L. S. PAGE, T. J. L o n , G . S. KOBAYASHI and G. MEDOFF, 1991 Characterization of CARE-1: Candida albicans repetitive element-1. Gene 1 0 2 45-50.
LODDER, S., K. GULL and D. WOOD, 1993 An electrophoretic karyo- type of the cultivated mushroom- Agaricus bispurus. Curr. Genet.
MAGEE, B. B., Y. KOLTON, J. A. GORMAN and P. T. MAGEE, 1988 Assignment of cloned genes to the seven electrophoretically sepa- rated Candida albicans chromosomes. Mol. Cell. Biol. 8: 4721- 4726.
IMALESZKA, R., and G. D. CLARK-WALKER, 1989 A petite positive strain of Huyveromyces Zuctis has a 300 kb deletion in the rDNA cluster. Curr. Genet. 1 6 429-432.
IMALESZKA, R., and G . D. CLARK-WALKER, 1993 Yeasts have a four- fold variation in ribosomal DNA copy number. Yeast 9 53-58.
MAO, Y., and B. M. TYLER, 1991 Genome organization of Phytophthora megaspenna fsp. glycinea. Exp.Myco1. 15: 283-291.
MARTIN, F. N., 1989 Maternal inheritance of mitochondrial DNA in sexual crosses of Pythium sylvaticum. Curr. Genet. 16: 373-374. MARTIN, F. N., 1990 Variation in the ribosomal DNA repeat unit within single oospore isolates of the genus Pythium. Genome 3 3
MARTIN, F. N., 1991 Linear mitochondrial molecules and intraspe- cific mitochondrial genome stability in a species of Qthium. Ge- nome 34: 156-162.
MARTIN, F. N., 1992 Pythium, pp. 39-49 in Methods for Research on Soilborne Phytopathogenic Fungi, edited by L. L. SINGELTON, J. D. MIHAIL and C. M. RUSH. APS Press, St. Paul, MN.
MARTIN, F. N., 1995 Electrophoretic karyotype polymorphisms in the genus Pythium. ( i n press) .
MARTIN, F. N., and H. C. KISTLER, 1990 Species-specific banding patterns of restriction endonucleasedigested mitochondrial DNA form the genus Pythium. Exp. Mycol. 14: 32-46.
MILLS, D., and K. MCCLUSKY, 1990 Electrophoretic karyotypes of fungi: the new cytology. Mol. Plant-Microbe Interact. 3 351- 357.
MONTGOMERY, E. A., S.-M. HUANG, C. H. LANGLEY and B. H. JUDD, 1991 Chromosome rearrangements by ectopic recombination in Drosophila melanagaster: genome structure and evolution. Ge- netics 129: 1085-1098.
NAUMOVA, E., G. NAUMOV, P. FOURNIER, H.-U. NGUYEN and C. GAIL LARDIN, 1993 Chromosomal polymorphism of the yeast Yarrowia
lipolytica and related species: electrophoretic karyotyping and hybridization with cloned genes. Curr. Genet. 2 3 450-454.
ONO, B., and Y. ISHINO-ARAO, 1988 Inheritance of chromosome length polymorphisms in Saccharomyces cermisiae. Curr. Genet. 14:
PASERO, P., and M. MARILLEY, 1993 Size variation of rDNA clusters in the yeasts Saccharomyces cermisiae and Schizosaccharomyces pombe. Mol. Gen. Genet. 236: 448-452.
PETES, T. D., 1979 Yeast ribosomal DNAgenes are located on chromo- some XII. Proc. Natl. Acad. Sci. USA 76: 410-414.
24: 496-499.
585-591.