Characterization of Group C and G Streptococcal Strains That Cause Streptococcal Toxic Shock Syndrome
Full text
Figure



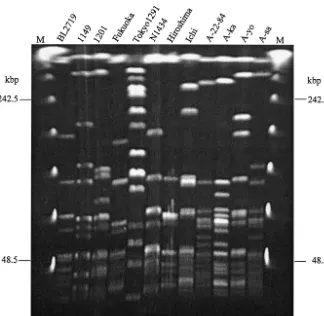
Related documents
It is reasonable, because Indonesian government has allocated at least 20 percent for education and health from the total Indonesian budget, while according to Economic
multi-varied MDF-SAR model with four descriptors it can be said that antiallergic activities of substituted N 4-methoxyphenyl benzamides strongly depend on the geometry of
The analysis included five factors that showed a premise that they could have an impact on the efficiency of the leadership style of managers: sex, the level of management,
In this analysis, Spearman and Kendall tau b correlation were selected because the data for currency exchange rate and crude oil price are non-normally distributed. Spearman
Hal tersebut terjadi karena Sumber Daya Manusia (SDM) pada kelurahan Ujung Menteng kurang potensial, hal ini dapat tergambar dari tingkat pendidkan masyarakat yang
Ultimately, the study of suicide among gay and bisexual youth may shed new light on the unifying characteristics of adolescent victims, unraveling the common threads of risk
However, this case portrays the dra- matic electrocardiographic appearance of acute myocardial ischemia in a boy with biopsy-proven myocarditis who had only mild chest pain.
Effect of pH on decolourization of Direct Blue 15 in wastewater medium containing glucose (0.1% w/v) under partial anaerobic condition at 45 o C.. Klebsiella
