JOURNAL OFCLINICALMICROBIOLOGY, July 2005, p. 3324–3333 Vol. 43, No. 7 0095-1137/05/$08.00⫹0 doi:10.1128/JCM.43.7.3324–3333.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Multilocus DNA Sequence Comparisons Rapidly Identify
Pathogenic Molds
Jennifer L. Rakeman,
1Uyen Bui,
1Karen LaFe,
1Yi-Ching Chen,
1Rhonda J. Honeycutt,
2and Brad T. Cookson
1,3*
Departments of Laboratory Medicine1and Microbiology,3University of Washington, Seattle, Washington, and Clarity BioSciences, Inc., Carlsbad, California2
Received 10 December 2004/Returned for modification 26 January 2005/Accepted 24 March 2005
The increasing incidence of opportunistic fungal infections necessitates rapid and accurate identification of the associated fungi to facilitate optimal patient treatment. Traditional phenotype-based identification meth-ods utilized in clinical laboratories rely on the production and recognition of reproductive structures, making identification difficult or impossible when these structures are not observed. We hypothesized that DNA sequence analysis of multiple loci is useful for rapidly identifying medically important molds. Our study included the analysis of the D1/D2 hypervariable region of the 28S ribosomal gene and the internal transcribed spacer (ITS) regions 1 and 2 of the rRNA operon. Two hundred one strains, including 143 clinical isolates and 58 reference and type strains, representing 43 recognized species and one possible new species, were examined. We generated a phenotypically validated database of 118 diagnostic alleles. DNA length polymorphisms detected among ITS1 and ITS2 PCR products can differentiate 20 of 33 species of molds tested, and ITS DNA sequence analysis permits identification of all species tested. For 42 of 44 species tested, conspecific strains displayed >99% sequence identity at ITS1 and ITS2; sequevars were detected in two species. For all 44 species, identifications by genotypic and traditional phenotypic methods were 100% concordant. Because dendrograms based on ITS sequence analysis are similar in topology to 28S-based trees, we conclude that ITS sequences provide phylogenetically valid information and can be utilized to identify clinically important molds. Addi-tionally, this phenotypically validated database of ITS sequences will be useful for identifying new species of pathogenic molds.
Fungal infections have been increasing in prevalence among hospitalized patients, particularly immunocompromised indi-viduals. Invasive mold infections have very high mortality rates; successful treatment requires rapid and accurate identification of the pathogen. Aspergillus is the most prevalent infectious mold in immunocompromised patients; however, other molds, such asFusariumspp. and Zygomycetes, are increasingly fre-quent causes of infection (2, 18, 30).
The phenotype-based identification methods routinely used in clinical laboratories require expertise and can be time-con-suming and laborious. Isolates are identified by recognition of colonial morphology and microscopic reproductive structures; few biochemical tests are available to aid in identification. Phenotypic variants may not be identifiable or may be misi-dentified. Typically in the clinical laboratory, isolates that can-not be identified by reproductive structures are described as
Mycelia sterilia, a name indicating a filamentous fungus that displays no distinguishing phenotypes recognized by routine clinical laboratory analyses.
Phenotypic methods can, in some cases, take weeks, a time frame that is not clinically useful. Rapid identification of molds causing invasive disease could facilitate the timely administra-tion of effective therapy. For example,Aspergillus flavusand
Aspergillus terreus have been reported to be resistant to
am-photericin B (39, 45), and Aspergillus fumigatus can become resistant to itraconazole (9).
Molecular methods for identification of pathogenic fungi have been validated for use in clinical settings (6, 7, 10, 14, 20–22, 32, 38, 48). rRNA genes, including the 28S gene (26S gene in all yeasts), are conserved, accrue single nucleotide changes at a relatively low rate, and provide useful phyloge-netic information (51). In eukaryotes, the rRNA operon in-cludes internal transcribed spacer regions 1 and 2 (ITS1 and ITS2), which do not encode functional rRNAs or proteins. These loci have increased levels of DNA sequence diversity compared to other loci within the operon, including the 28S gene (19, 51). Thus, ITS sequences may provide accurate iden-tification of closely related isolates and species which cannot readily be distinguished using 26S or 28S rRNA gene se-quences. This hypothesis has been confirmed for yeasts iso-lated in clinical laboratories (6–8, 14, 43, 48). For example, ITS sequence analysis provides a more accurate taxonomic place-ment for a number of yeasts, includingCryptococcus humicolus,
Pichia veronae, andP. fabianii(6), which were not resolved by analysis of 26S sequences. Henry et al. (22) demonstrated sufficient sequence diversity at ITS1 and ITS2 to distinguish six medically important Aspergillus species from one another. Turenne et al. (48) and De Baere et al. (8) used capillary electrophoresis to examine the lengths of ITS2 PCR products for identifying fungi. This method is rapid but does not provide species-specific identification for all organisms tested, demon-strating a need for additional analyses.
We have developed a rapid molecular method for
identify-* Corresponding author. Mailing address: Departments of Labora-tory Medicine and Microbiology, University of Washington, Box 357110, Seattle, WA 98195. Phone: (206) 6131. Fax: (206) 598-6189. E-mail: [email protected].
3324
on May 16, 2020 by guest
http://jcm.asm.org/
ing pathogenic molds based on the lengths and sequences of ITS1 and ITS2. The combination of ITS1 and ITS2 length polymorphisms identifies 20 of 33 species of molds tested. Analysis of ITS1 and ITS2 DNA sequences unambiguously identified all molds tested to the species level; 44 species were represented in the analysis. Identification by ITS DNA se-quence analyses was concordant with, but more specific than, identification by 28S DNA sequence analysis and phenotypic analyses. ITS-based phylogenetic trees are similar in overall topology to those constructed with 28S sequences, demonstrat-ing that ITS loci are taxonomically informative. Therefore, phylograms constructed with ITS sequences will allow accurate taxonomic assignment of previously unidentified or uncharac-terized molds. We present here a database of ITS DNA se-quences, validated with 28S DNA sequence and phenotypic analyses, that will allow rapid identification of molds isolated in the clinical laboratory by ITS DNA sequence comparisons.
MATERIALS AND METHODS
Strains.Two hundred one strains of molds, representing 44 species, were characterized in this study. Our phenotypically validated database includes in-formation from type strains obtained from either the American Type Culture Collection (ATCC) (n⫽10) or the Centraalbureau voor Schimmelcultures (CBS) (n⫽17). Clinical strains (University of Washington Fungal Project
[UWFP]) (n⫽143) were isolated in the mycology laboratory at the University of Washington Medical Center. ITS1⫹2 (n⫽13), 28S (n⫽15), or ITS1⫹2 and 28S (n⫽3) DNA sequence information from 31 reference strains (including 22 type strains) available in public databases was included in the sequence analyses. (The ITS region referred to here as ITS1⫹2 is amplified using primer binding sites in the 3⬘end of the 18S rRNA gene and the 5⬘end of the 28S rRNA gene: the resulting PCR product includes the entirety of ITS1, the 5.8S rRNA gene, and ITS2.). Strains are listed in Table 2. An additional 42 clinical isolates (Aspergillus flavus[n⫽1],A. fumigatus[n⫽24],A. niger[n⫽9], andTrichophyton rubrum
[n⫽ 8]) were analyzed by phenotypic methods and ITS1 and ITS2 length polymorphism analysis only (Table 1).
Morphological and biochemical analyses.All molds were identified with stan-dard phenotype-based algorithms used in clinical laboratories. Except where noted, all media and stains were obtained from Remel, Inc. (Lenexa, KS). Initial identification was based on colonial morphology of isolates grown on Sabouraud dextrose agar (as modified by Emmons) (SDA) plates, inhibitory mold agar, mycobiotic agar, and/or brain heart infusion agar with sheep blood with or without chloramphenicol (50g/ml), gentamicin (40g/ml), and cycloheximide (500g/ml) (media used depended on the collection site of the specimen) and microscopic morphology of lactophenol aniline blue-stained preparations (4, 13, 26). Molds other than presumptiveAspergillusspp. were grown on potato dex-trose agar for microscopic analyses, and potato flake agar was used for slide culture analysis of presumptiveAcremonium,Fusarium,Exophalia,Hortea, and
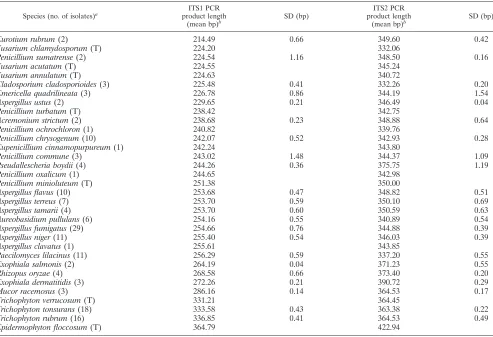
[image:2.585.47.540.80.417.2]Cladosporiumspp. (42). Based on these results, the following additional tests were performed as noted.Aspergillusspp. were identified by morphology on malt extract agar made with 15 g of agar per liter (37) and Czapek-Dox agars (Difco, Detroit, MI) (27, 37).A. fumigatuswas differentiated by its ability to grow at elevated temperatures (42°C) on SDA slants (BBL Microbiology Systems, Inc., TABLE 1. ITS1 and ITS2 length polymorphisms distinguish mold species
Species (no. of isolates)a
ITS1 PCR product length
(mean bp)b
SD (bp)
ITS2 PCR product length
(mean bp)b
SD (bp)
Eurotium rubrum(2) 214.49 0.66 349.60 0.42
Fusarium chlamydosporum(T) 224.20 332.06
Penicillium sumatrense(2) 224.54 1.16 348.50 0.16
Fusarium acutatum(T) 224.55 345.24
Fusarium annulatum(T) 224.63 340.72
Cladosporium cladosporioides(3) 225.48 0.41 332.26 0.20
Emericella quadrilineata(3) 226.78 0.86 344.19 1.54
Aspergillus ustus(2) 229.65 0.21 346.49 0.04
Penicillium turbatum(T) 238.42 342.75
Acremonium strictum(2) 238.68 0.23 348.88 0.64
Penicillium ochrochloron(1) 240.82 339.76
Penicillium chrysogenum(10) 242.07 0.52 342.93 0.28
Eupenicillium cinnamopurpureum(1) 242.24 343.80
Penicillium commune(3) 243.02 1.48 344.37 1.09
Pseudallescheria boydii(4) 244.26 0.36 375.75 1.19
Penicillium oxalicum(1) 244.65 342.98
Penicillium minioluteum(T) 251.38 350.00
Aspergillus flavus(10) 253.68 0.47 348.82 0.51
Aspergillus terreus(7) 253.70 0.59 350.10 0.69
Aspergillus tamarii(4) 253.70 0.60 350.59 0.63
Aureobasidium pullulans(6) 254.16 0.55 340.89 0.54
Aspergillus fumigatus(29) 254.66 0.76 344.88 0.39
Aspergillus niger(11) 255.40 0.54 346.03 0.39
Aspergillus clavatus(1) 255.61 343.85
Paecilomyces lilacinus(11) 256.29 0.59 337.20 0.55
Exophiala salmonis(2) 264.19 0.04 371.23 0.55
Rhizopus oryzae(4) 268.58 0.66 373.40 0.20
Exophiala dermatitidis(3) 272.26 0.21 390.72 0.29
Mucor racemosus(3) 286.16 0.14 364.53 0.17
Trichophyton verrucosum(T) 331.21 364.45
Trichophyton tonsurans(18) 333.58 0.43 363.38 0.22
Trichophyton rubrum(16) 336.85 0.41 364.53 0.49
Epidermophyton floccosum(T) 364.79 422.94
a
T indicates that the PCR product from the type strain only was analyzed.
b
ITS1 PCR products were generated using primers ITS1 and ITS2, ITS2 PCR products were generated using primers ITS3 and ITS4, and PCR product lengths were determined by denaturing capillary electrophoresis as described in Materials and Methods.
VOL. 43, 2005 RAPID IDENTIFICATION OF PATHOGENIC MOLDS 3325
on May 16, 2020 by guest
http://jcm.asm.org/
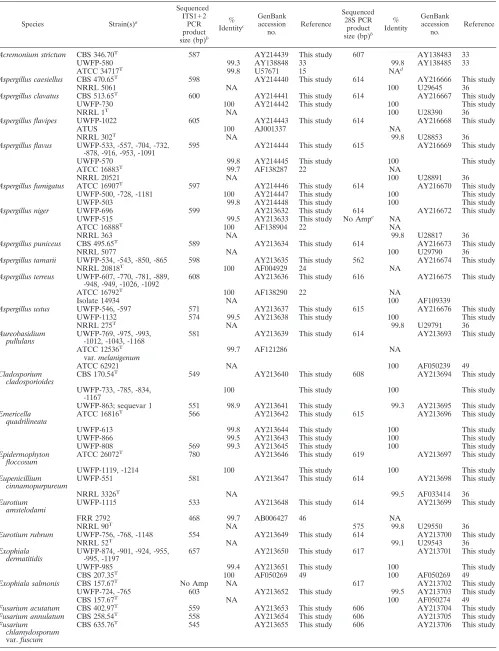
TABLE 2. ITS and 28S sequences are⬎99% identical between conspecific type strains and clinical isolates
Species Strain(s)a
Sequenced ITS1⫹2
PCR product size (bp)b
% Identityc
GenBank accession
no.
Reference
Sequenced 28S PCR
product size (bp)b
% Identity
GenBank accession
no.
Reference
Acremonium strictum CBS 346.70T 587 AY214439 This study 607 AY138483 33
UWFP-580 99.3 AY138848 33 99.8 AY138485 33
ATCC 34717T 99.8 U57671 15 NAd
Aspergillus caesiellus CBS 470.65T 598 AY214440 This study 614 AY216666 This study
NRRL 5061 NA 100 U29645 36
Aspergillus clavatus CBS 513.65T 600 AY214441 This study 614 AY216667 This study
UWFP-730 100 AY214442 This study 100 This study
NRRL 1T
NA 100 U28390 36
Aspergillus flavipes UWFP-1022 605 AY214443 This study 614 AY216668 This study
ATUS 100 AJ001337 NA
NRRL 302T NA 99.8 U28853 36
Aspergillus flavus UWFP-533, -557, -704, -732,
-878, -916, -953, -1091 595 AY214444 This study 615 AY216669 This study
UWFP-570 99.8 AY214445 This study 100 This study
ATCC 16883T 99.7 AF138287 22 NA
NRRL 20521 NA 100 U28891 36
Aspergillus fumigatus ATCC 16907T 597 AY214446 This study 614 AY216670 This study
UWFP-500, -728, -1181 100 AY214447 This study 100 This study
UWFP-503 99.8 AY214448 This study 100 This study
Aspergillus niger UWFP-696 599 AY213632 This study 614 AY216672 This study
UWFP-515 99.5 AY213633 This study No Ampe NA
ATCC 16888T 100 AF138904 22 NA
NRRL 363 NA 99.8 U28817 36
Aspergillus puniceus CBS 495.65T
589 AY213634 This study 614 AY216673 This study
NRRL 5077 NA 100 U29790 36
Aspergillus tamarii UWFP-534, -543, -850, -865 598 AY213635 This study 562 AY216674 This study
NRRL 20818T
100 AF004929 24 NA
Aspergillus terreus UWFP-607, -770, -781, -889,
-948, -949, -1026, -1092 608 AY213636 This study 616 AY216675 This study ATCC 16792T
100 AF138290 22 NA
Isolate 14934 NA 100 AF109339
Aspergillus ustus UWFP-546, -597 571 AY213637 This study 615 AY216676 This study
UWFP-1132 574 99.5 AY213638 This study 100 This study
NRRL 275T NA 99.8 U29791 36
Aureobasidium
pullulans UWFP-769, -975, -993,-1012, -1043, -1168 581 AY213639 This study 614 AY213693 This study
ATCC 12536T 99.7 AF121286 NA
var.melanigenum
ATCC 62921 NA 100 AF050239 49
Cladosporium cladosporioides
CBS 170.54T 549 AY213640 This study 608 AY213694 This study
UWFP-733, -785, -834,
-1167 100 This study 100 This study
UWFP-863; sequevar 1 551 98.9 AY213641 This study 99.3 AY213695 This study
Emericella
quadrilineata ATCC 16816
T 566 AY213642 This study 615 AY213696 This study
UWFP-613 99.8 AY213644 This study 100 This study
UWFP-866 99.5 AY213643 This study 100 This study
UWFP-808 569 99.3 AY213645 This study 100 This study
Epidermophyton
floccosum ATCC 26072
T
780 AY213646 This study 619 AY213697 This study
UWFP-1119, -1214 100 This study 100 This study
Eupenicillium
cinnamopurpureum UWFP-551 581 AY213647 This study 614 AY213698 This study
NRRL 3326T NA 99.5 AF033414 36
Eurotium
amstelodami UWFP-1115 533 AY213648 This study 614 AY213699 This study
FRR 2792 468 99.7 AB006427 46 NA
NRRL 90T
NA 575 99.8 U29550 36
Eurotium rubrum UWFP-756, -768, -1148 554 AY213649 This study 614 AY213700 This study
NRRL 52T NA 99.1 U29543 36
Exophiala
dermatitidis UWFP-874, -901, -924, -955,-995, -1197 657 AY213650 This study 617 AY213701 This study
UWFP-985 99.4 AY213651 This study 100 This study
CBS 207.35T 100 AF050269 49 100 AF050269 49
Exophiala salmonis CBS 157.67T No Amp NA 617 AY213702 This study
UWFP-724, -765 603 AY213652 This study 99.5 AY213703 This study
CBS 157.67T NA 100 AF050274 49
Fusarium acutatum CBS 402.97T 559 AY213653 This study 606 AY213704 This study
Fusarium annulatum CBS 258.54T
558 AY213654 This study 606 AY213705 This study
Fusarium chlamydosporum
var.fuscum
CBS 635.76T 545 AY213655 This study 606 AY213706 This study
Continued on facing page
3326
on May 16, 2020 by guest
http://jcm.asm.org/
TABLE 2—Continued
Species Strain(s)a
Sequenced ITS1⫹2
PCR product size (bp)b
% Identityc
GenBank accession
no.
Reference
Sequenced 28S PCR
product size (bp)b
% Identity
GenBank accession
no.
Reference
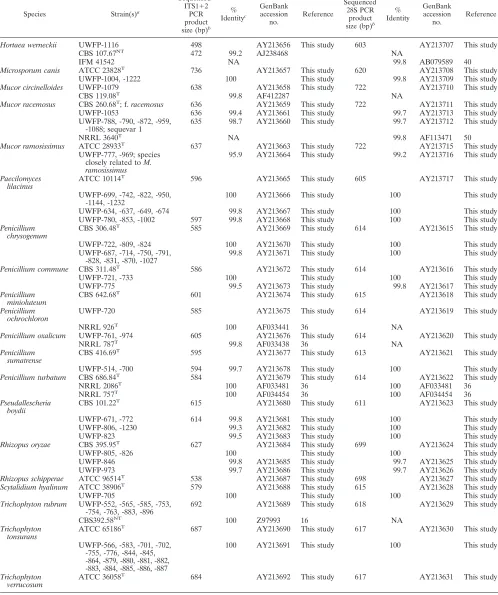
Hortaea werneckii UWFP-1116 498 AY213656 This study 603 AY213707 This study
CBS 107.67NT
472 99.2 AJ238468 NA
IFM 41542 NA 99.8 AB079589 40
Microsporum canis ATCC 23828T 736 AY213657 This study 620 AY213708 This study
UWFP-1004, -1222 100 This study 99.8 AY213709 This study
Mucor circinelloides UWFP-1079 638 AY213658 This study 722 AY213710 This study
CBS 119.08T 99.8 AF412287 NA
Mucor racemosus CBS 260.68T; f.racemosus 636 AY213659 This study 722 AY213711 This study
UWFP-1053 636 99.4 AY213661 This study 99.7 AY213713 This study UWFP-788, -790, -872, -959,
-1088; sequevar 1 635 98.7 AY213660 This study 99.7 AY213712 This study
NRRL 3640T NA 99.8 AF113471 50
Mucor ramosissimus ATCC 28933T 637 AY213663 This study 722 AY213715 This study
UWFP-777, -969; species closely related toM. ramosissimus
95.9 AY213664 This study 99.2 AY213716 This study
Paecilomyces
lilacinus ATCC 10114
T
596 AY213665 This study 605 AY213717 This study
UWFP-699, -742, -822, -950, -1144, -1232
100 AY213666 This study 100 This study
UWFP-634, -637, -649, -674 99.8 AY213667 This study 100 This study UWFP-780, -853, -1002 597 99.8 AY213668 This study 100 This study
Penicillium
chrysogenum CBS 306.48
T 585 AY213669 This study 614 AY213615 This study
UWFP-722, -809, -824 100 AY213670 This study 100 This study UWFP-687, -714, -750, -791,
-828, -831, -870, -1027
99.8 AY213671 This study 100 This study
Penicillium commune CBS 311.48T 586 AY213672 This study 614 AY213616 This study
UWFP-721, -733 100 This study 100 This study
UWFP-775 99.5 AY213673 This study 99.8 AY213617 This study
Penicillium
minioluteum CBS 642.68
T
601 AY213674 This study 615 AY213618 This study
Penicillium ochrochloron
UWFP-720 585 AY213675 This study 614 AY213619 This study
NRRL 926T 100 AF033441 36 NA
Penicillium oxalicum UWFP-761, -974 605 AY213676 This study 614 AY213620 This study
NRRL 787T 99.8 AF033438 36 NA
Penicillium
sumatrense CBS 416.69
T
595 AY213677 This study 613 AY213621 This study
UWFP-514, -700 594 99.7 AY213678 This study 100 This study
Penicillium turbatum CBS 686.84T
584 AY213679 This study 614 AY213622 This study
NRRL 2086T 100 AF033481 36 100 AF033481 36
NRRL 757T 100 AF034454 36 100 AF034454 36
Pseudallescheria boydii
CBS 101.22T 615 AY213680 This study 611 AY213623 This study
UWFP-671, -772 614 99.8 AY213681 This study 100 This study UWFP-806, -1230 99.3 AY213682 This study 100 This study
UWFP-823 99.5 AY213683 This study 100 This study
Rhizopus oryzae CBS 395.95T
627 AY213684 This study 699 AY213624 This study
UWFP-805, -826 100 This study 100 This study
UWFP-846 99.8 AY213685 This study 99.7 AY213625 This study UWFP-973 99.7 AY213686 This study 99.7 AY213626 This study
Rhizopus schipperae ATCC 96514T
538 AY213687 This study 698 AY213627 This study
Scytalidium hyalinum ATCC 38906T 579 AY213688 This study 615 AY213628 This study
UWFP-705 100 This study 100 This study
Trichophyton rubrum UWFP-552, -565, -585, -753,
-754, -763, -883, -896 692 AY213689 This study 618 AY213629 This study
CBS392.58NT 100 Z97993 16 NA
Trichophyton tonsurans
ATCC 65186T 687 AY213690 This study 617 AY213630 This study
UWFP-566, -583, -701, -702, -755, -776, -844, -845, -864, -879, -880, -881, -882, -883, -884, -885, -886, -887
100 AY213691 This study 100 This study
Trichophyton verrucosum
ATCC 36058T 684 AY213692 This study 617 AY213631 This study
a
Type strains are denoted by superscript “T”; neotype strains are denoted by superscript “NT.” Type strains listed first for an organism and all UWFP strains were analyzed by DNA sequence and phenotypic methods in our laboratory. UWFP isolates grouped together are 100% identitical at ITS1⫹2 and 28S. NRRL, Northern Regional Research Laboratory.
b
PCR product length was determined by sequencing as described in Materials and Methods. ITS1⫹2 PCR products were generated using ITS1 and ITS4 primers; the 28S D1/D2 region was amplified using primers NL-1 and NL-4.
c
Percent identity with the first strain listed for each organism. Strains listed together exhibit 100% sequence identity at ITS1⫹2 and 28S.
d
NA, not available.
e
No Amp, product did not amplify in several attempts with independently extracted DNA samples.
3327
on May 16, 2020 by guest
http://jcm.asm.org/
Sparks, MD) (44). To aid in the differentiation ofAcremoniumspp. andFusarium
spp., growth rate on SDA plates was observed (41, 42).Aureobasidiumspp. were identified by phenotypic analysis of Dalmau preparations from isolates grown on cornmeal agar with Tween 80 (44).Exophaliaspp. andHorteaspp. were tested for growth at 37°C and 42°C on SDA slants and were assayed for KNO3 utili-zation using potassium nitrate agar (42) and casein and tyrosine hydrolysis using
Nocardiaquad plates (29).Rhizopusspp. were tested for growth at 30°C and elevated temperature (42°C) on SDA slants (29). To differentiateCladosporium
spp. fromCladophialophora spp., gelatin liquefaction using nutrient gelatin (12%) media and growth at 37°C and 42°C on SDA slants was performed (29). The dermatophytesEpidermophytonspp.,Microsporumspp., andTrichophyton
spp. were tested for urease activity using Christiansen urea agar, reactions on bromcresol purple-milk solids-glucose agar, and morphology and pigment pro-duction on potato dextrose agar (25).Trichophytonspp. were further differenti-ated using vitamin test agars (T1 to T7) (25).
DNA extraction.Mold isolates grown on SDA plates at 30°C for 72 h were resuspended in 1 ml lysis buffer (100 mM Tris, 0.5% [wt/vol] sodium dodecyl sulfate, 3 mM EDTA, pH 7.5) containing approximately one-third volume 0.5-mm glass beads in 2-ml Bead-Beater tubes (BioSpec Products, Inc., Bartles-ville, OK). The vials were placed into a Mini-BeadBeater-8 (BioSpec Products, Inc., Bartlesville, OK) at high speed (3,200 rpm) for 3 min and then placed on ice. After centrifugation at 20,000⫻gfor 5 min, 800l of supernatant was trans-ferred to a new tube. Twenty microliters of proteinase K (20 mg/ml) was added, the tubes were incubated for 1 h at 37°C, and the proteinase K was inactivated by incubation at 65°C for 20 min. The DNA was extracted with 700l Tris-saturated phenol chloroform (1:1) two times followed by a chloroform extraction and ethanol precipitation. The DNA pellet was dried and resuspended in 150l of sterile nuclease-free water and treated with 12l of RNase (10 mg/ml) for 1 h at 37°C.
PCR and DNA sequencing.The ITS1⫹2 regions were PCR amplified using primers ITS1 and ITS4 (6, 7), and both strands of the PCR products were directly sequenced (7). In separate experiments, the ITS1 locus was amplified using primers ITS1 and ITS2 (6), and similarly, the ITS2 locus was amplified using primers ITS3 and ITS4 (7). The lengths of the resultant amplicons were deter-mined with single-nucleotide precision by automated capillary electrophoresis under denaturing conditions using an ABI310 genetic analyzer and GeneScan software (PE Applied Biosystems, Foster City, CA) as described previously (7). PCR amplification and sequencing of the D1/D2 hypervariable region of the 28S rRNA gene was performed using primers NL-1 and NL-4 as described previously (28).
Sequence and phylogenetic analysis.DNA sequences were aligned, edited, and analyzed as described previously (7); in addition, GeneStream align (35; http://xylian.igh.cnrs.fr/) was used. Manual editing was performed in Jalview version 1.3b (M. Clamp, European Bioinformatics Institute [http://circinus.ebi .ac.uk:6543/jalview/]) and with standard word processing software. CLUSTAL_X version 1.81 (47) was used for phylogenetic analysis. Dendrograms constructed with the neighbor-joining treeing algorithm were evaluated with 1,000 bootstrap analyses using CLUSTAL_X and visualized in Treeview version 1.5.3 (34).
RESULTS
ITS DNA sequence polymorphisms identify pathogenic molds to the species level.To test the hypothesis that genetic information in the ITS1 and ITS2 loci of pathogenic molds is diagnostically useful, we analyzed ITS length (Table 1) and sequence polymorphisms (Table 2). One hundred sixty-six strains representing 33 species of molds were analyzed to de-termine whether ITS1 and ITS2 PCR product length polymor-phisms (Table 1) are sufficient to identify clinically relevant molds to the species level. All isolates were identified by tra-ditional colonial and morphological analyses and appropriate biochemical tests (see Materials and Methods). Single PCR products amplified from purified mold genomic DNA were analyzed by capillary electrophoresis under denaturing condi-tions, which allows rapid determination of DNA fragment length with single-base-pair accuracy (7). ITS1 (3⬘end of the 18S rRNA gene, all of ITS1, and the 5⬘end of the 5.8S gene) and ITS2 (the 3⬘end of the 5.8S gene, all of ITS2, and the 5⬘
end of the 28S gene) PCR product length polymorphisms dis-tinguish 20 of 33 mold species tested (Table 1). Intraspecies ITS1 and ITS2 length variations obscure the distinction of
Penicillium sumatrensefromEmericella quadralineataand Pen-icillium communefromPenicillium oxalicum(Table 1). In con-trast, isolates ofAspergillus fumigatusandPseudallescheria boy-dii also displayed intraspecies variation in ITS PCR product lengths. However, they are identified by characteristic length polymorphisms because the distribution of ITS lengths for each group is sufficiently different to distinguish them from other molds. All PCR product length measurements were con-firmed by direct sequencing (see below), and a close correla-tion between the electrophoresis results (Table 1) and the actual number of nucleotides that comprise the ITS1 and ITS2 amplicons was observed.
The entire ITS locus (ITS1⫹2 from the 3⬘ end of the 18S gene to the 5⬘end of the 28S gene (ITS1 plus the entire 5.8S gene plus ITS2), and the D1/D2 hypervariable region of the 28S rRNA gene were analyzed by direct sequencing of both the forward and reverse strands of the respective PCR products (Table 2). Forty-four species were represented in the analysis, and type strain data for all 43 recognized species was included and validated by phenotypic analyses; one probable new spe-cies was also identified (see below). One hundred forty-three clinical isolates and 27 type strains were analyzed phenotypi-cally and genotypiphenotypi-cally in our laboratory. Inclusion of sequence information for 31 reference strains from GenBank was justi-fied by identity to DNA sequences from fungal isolates phe-notypically and gephe-notypically identified in our laboratory (Ta-ble 2). ITS1 and ITS2 DNA sequence analyses specifically identify all 44 species tested. Conspecific strains (including type strains) demonstrate⬎99% sequence identity at the ITS1 and ITS2 loci for all but two species tested (Cladosporium cladosporioides and Mucor racemosus) (Table 2 and below). Further, phenotypic designations were 100% concordant with ITS1, ITS2, and 28S DNA sequence-based identifications.
ITS sequence analyses refine identification: sequevars and probable new species defined.Our data and those of others have demonstrated that conspecific fungi are⬎99% identical at the 28S/26S and ITS loci (6, 7, 28, 43). Genetic variants among isolates of a species are designated sequevars (7) when an isolate is (i) phenotypically consistent with the type strain, (ii) genotypicallyⱖ99% identical to the type strain at multiple loci, and (iii)⬍99% identical to the type strain at one diag-nostic locus (6). We identified isolates of several species of molds displaying increased sequence diversity (⬍99% identity) at one of the ITS loci, together with high levels of conservation (ⱖ99% identity) at the 28S gene and the other ITS locus and phenotypic concordance (Table 3). These data (i) indicate that sequence diversity independently accumulates in each locus (ITS1, ITS2, and 28S) and (ii) are consistent with previous observations of clinically important yeasts (6). On this basis, we submit that the analyses summarized in Table 2 represent multilocus sequence-based identifications.
Analyses of multiple DNA loci defines a sequevar within the species Cladosporium cladosporioides. In the clinical labora-tory, phenotypic characteristics differentiateCladosporiumspp. from Cladophialophora spp. These analyses include colonial and microscopic analysis, gelatin liquefaction, and tests for the ability to grow at 37°C and 42°C (29), yet these data do not
3328 RAKEMAN ET AL. J. CLIN. MICROBIOL.
on May 16, 2020 by guest
http://jcm.asm.org/
provide further classification ofCladosporium isolates to the species level. Sequence analyses allow speciation of these iso-lates, including isolates ofCladosporium cladosporioides, and 28S- and ITS-based identifications are 100% concordant (Ta-ble 3). We identified a clinical isolate, UWFP-863, that is phenotypically consistent with Cladosporium spp., and is ⬎99% identical to the type strain at the 28S and ITS2 loci. UWFP-863 is only 98.3% identical to the type strain at the ITS1 locus (Table 3), which further refines the classification of this strain as a sequevar distinguishable from other isolates of
C. cladosporioidesand the type strain. We have designated this strainC. cladosporioidessequevar 1.
Similarly, traditional phenotypic analyses in the clinical lab-oratory readily identify Mucorspp. to the genus level, while genetic analyses can differentiateMucor racemosusfrom other
Mucorspecies. A group of five clinical isolates (UWFP-788, -790, -827, -959, and -1088) that exhibit phenotypes consistent withMucorspp. are 100% identical to each other at the 28S, ITS1, and ITS2 loci and show⬎99% identity to theM. race-mosus type strain (CBS 260.68) at 28S and ITS1 (Table 3). However, identity between the clinical isolates and the type strain at ITS2 is 98.6%. Therefore, the clinical isolates are designatedM. racemosussequevar 1.
DNA sequence analyses of multiple loci can also identify species that are not differentiated by phenotypic or single-locus sequence analyses. Two of the 143 clinical isolates we analyzed, strains UWFP-777 and UWFP-969, potentially represent a new species that is closely related toMucor ramosissimus. The strains, isolated from two different patients, have identical ITS1, ITS2, and 28S DNA sequences and exhibit phenotypes consistent withMucorspp. These strains display 99.2% identity to the M. ramosissimus type strain ATCC 28933 at the 28S locus but only 95.1% and 96.0% identity at ITS1 and ITS2, respectively (Table 3). Sequence diversity of this magnitude at two loci compared to the type strain sequences suggests that UWFP-777 and -969 most likely represent members of another species ofMucorclosely related toM. ramosissimus. These data show that analysis of multiple loci extends the specificity of identification of Mucorspecies beyond the capabilities of ei-ther traditional methods routinely used in the clinical labora-tory or 28S DNA sequence analysis alone.
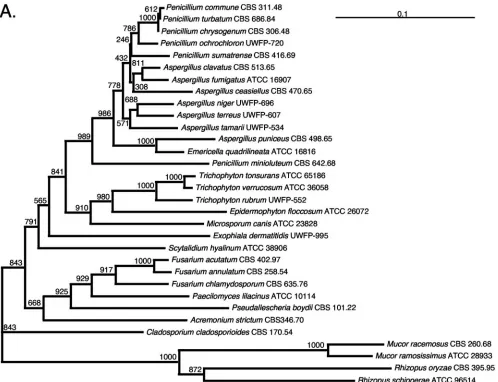
Phylogenetic analysis.rRNA gene sequences, including 28S sequences, have been shown to provide taxonomically useful information (51). To determine whether ITS sequences could accurately identify relationships among diverse mold taxa, we compared phylogenetic trees constructed with ITS sequences to those constructed with 28S sequences. The topologies of the ITS and 28S trees are highly similar (Fig. 1). In addition, high bootstrap values are observed at the deeply branching nodes, and similarly high values are observed at branches separating more closely related genera. Thus, the ITS tree demonstrates that there are species-specific, phylogenetically informative, taxonomically useful sequences within the ITS loci of patho-genic molds. The high level of intraspecies sequence identity (Table 2) demonstrate the stability of the loci within species, and the high bootstrap values (Fig. 1) provide confidence in the species-specific designations obtained from our sequence analyses. The data presented in Fig. 1 and Table 2 indicate that previously uncharacterized molds could be identified in the clinical laboratory by placing their ITS sequences into an es-tablished tree constructed with DNA sequence information derived from phenotypically validated mold isolates.
DISCUSSION
[image:6.585.44.542.80.206.2]DNA-based identification has been utilized successfully to identify pathogenic fungi (1, 3, 5–7, 10–12, 14, 20–23, 31, 32, 38, 48, 52, 53). In order for molecular identification methods to be successful, it is imperative that phenotypically well-charac-terized mold isolates from clinical samples are used for devel-oping the method and building the sequence database. In this work, we demonstrate that ITS1 and ITS2 DNA sequences are useful for identification of 44 species of pathogenic molds; our findings are validated with 28S DNA sequence and phenotypic information from 143 clinical isolates, 27 type strains, and genetic information from an additional 31 strains. One hun-dred eighteen diagnostic alleles are represented in our pheno-typically validated database. ITS1 and ITS2 PCR product lengths as determined by capillary electrophoresis distinguish 20 of 33 species of molds tested. However, analysis of ITS DNA sequences provides definitive identification to the spe-cies level for all mold isolates tested. ITS-based identification
TABLE 3. ITS sequence analysis identifies sequevars within species and defines possible new species
Strain(s) Identity by
traditional analysesa Identity by genetic analysesb
Sequence similarity to type strain (%)c
28S ITS1 ITS2
CBS 170.54 Cladosporium Cladosporium cladosporioides Type strain Type strain Type strain
UWFP-773, -785, -834 Cladosporium Cladosporium cladosporioides 100 100 100
UWFP-863 Cladosporium Cladosporium cladosporioides
sequevar 1
99.3 98.3 99.4
CBS 260.68 Mucor Mucor racemosusf.racemosus Type strain Type strain Type strain
UWFP-788, -790, -827, -959, -1088
Mucor Mucor racemosussequevar 1 99.7 99.3 98.6
ATCC 28933 Mucor Mucor ramosissimus Type strain Type strain Type strain
UWFP-777, -969 Mucor Species closely related to
Mucor ramosissimus
99.2 95.1 96.0
a
Isolates were identified by phenotypic analyses as described in Materials and Methods.
b
Isolates were further identified by ITS1 and -2 and 28S sequence analysis as described in Materials and Methods. Strains listed together exhibit 100% sequence identity at all three loci.
c
Sequence identity was determined by pairwise analysis of the indicated sequences.
VOL. 43, 2005 RAPID IDENTIFICATION OF PATHOGENIC MOLDS 3329
on May 16, 2020 by guest
http://jcm.asm.org/
is 100% concordant with identification provided by 28S DNA sequence analysis and phenotypic methods. In addition, anal-ysis of multiple genetic loci differentiates sequevars within spe-cies (i.e.,CladosporiumandMucor). Finally, phylogenetic re-lationships are accurately predicted by ITS sequences, as ITS-based trees are topologically similar to trees ITS-based on 28S sequence analysis.
The integration of molecular analyses with traditional phe-notypic methods of fungal identification can significantly in-crease the specificity and dein-crease the turnaround time for the identification of clinically important molds. Production of re-productive structures or expression of specific biochemical phenotypes by fungi grown in culture is required for pheno-typic analyses. Previously uncharacterized molds are difficult to identify by such methods, yet accurate taxonomic placement of the uncharacterized molds, and hence their identity, can be
inferred from their phylogenetic relationships with well-char-acterized mold isolates. Molecular analyses of multiple DNA loci can provide a more rapid (24 h versus several weeks) and sometimes more specific identification of an organism than traditional culture-based methods: identification is refined by including ITS and 28S sequence analyses with phenotypic char-acterization. The increased level of sequence variation at ITS loci compared to the 28S locus will facilitate the future devel-opment of rapid, hybridization-based identification methods (38).
Using phenotypic methods alone, an isolate ofCladosporium
[image:7.585.46.544.69.451.2]spp. can be differentiated fromCladophialophoraspp. in a time frame of 1 to 4 weeks. In addition to colonial and microscopic morphology, two tests are performed to identify the organism in our laboratory: gelatin liquefaction and temperature toler-ance (growth at 37°C and 42°C) (29). Molecular analysis of the
FIG. 1. 28S and ITS1/ITS2 DNA sequence-based phylogenetic trees of clinically significant molds. Trees were drawn using the neighbor-joining method with 1,000 bootstrap analyses. Numbers at the nodes indicate the bootstrap values, and the bars indicate relative genetic distance. The source of the organism or sequence is indicated in Table 2. The zygomycetesMucorspp. andRhizopusspp. were used as outgroups in both trees. The trees include organisms for which DNA from the type strain was sequenced in our laboratory or for which type strain sequence data were verified by identity to a DNA sequence generated in our laboratory from a conspecific isolate. (A) ITS-based tree of 32 molds. Analysis included partial 18S and 28S sequences and complete ITS1, 5.8S, and ITS2 sequences. (B) 28S-based tree of 27 molds. Analysis included the D1/D2 hypervariable region of the 28S rRNA gene.
3330 RAKEMAN ET AL. J. CLIN. MICROBIOL.
on May 16, 2020 by guest
http://jcm.asm.org/
ITS1, ITS2, and 28S DNA sequences of the isolate can identify
C. cladosporioides to the species and sequevar levels. This analysis can be completed in 24 h once the organism is isolated from a patient specimen.
Multilocus analysis allows the separation of possible new species that are not differentiated by standard clinical labora-tory phenotypic analyses or single-locus analyses alone. Voigt et al. (50) used 18S and 28S sequences to describe the phylog-eny of medically important zygomycetes, including six species ofMucor.Mucor ramosissimus (NRRL 3042) andMucor cir-cinelloides(NRRL 3631) show 99.8% identity at the 18S locus and 99.2% identity at the 28S locus. The strain ofM. ramosis-simusused in the Voigt et al. study, NRRL 3042, shows 100% identity at the 28S locus to the M. ramosissimus type strain included in our study, ATCC 28933. Consistent with the ap-parent monophylogeny of these species based on 18S and 28S analysis alone, we identified two clinical isolates (UWFP-777 and -969) that display⬎99% 28S DNA sequence identity to the type strain ofM. ramosissimus (ATCC 28933); however, these isolates show only 98.9% identity at the 28S locus to that ofM. circinelloides NRRL 3631, demonstrating that
[image:8.585.46.538.70.458.2]UWFP-777 and -969 are not M. circinelloides. ITS DNA sequence analyses clearly differentiate UWFP-777 and -969 from theM. ramosissimus type strain because they show significant se-quence divergence at ITS1 and ITS2 (95.1 and 96.0%, respec-tively) compared with ATCC 28933. These strains may repre-sent another species closely related to M. ramosissimus that would not be recognized by 28S or phenotypic analysis alone. TheTrichophyton rubrum complex has recently been char-acterized at the molecular level (17). A group of 11 species and varieties were reclassified asT. rubrum based on analysis of ITS1, ITS2, and 5.8S DNA sequence analyses, PCR finger-printing, and amplified fragment length polymorphism analy-sis. ITS-based phylograms of this group revealed a tree in which 11 species and varieties grouped together in two distinct clades. The ITS DNA sequences from different species within the formerT. rubrumgroup are not identical to each other, as evidenced by the arrangement of the ITS-based phylogenetic tree (17). Our analysis supports the polyphyletic nature of the newly designatedT. rubrum group. The eight clinical isolates that we analyzed are 100% identical to each other at ITS1, ITS2, and 28S and are 100% identical at ITS1 and ITS2 to the
FIG. 1—Continued.
VOL. 43, 2005 RAPID IDENTIFICATION OF PATHOGENIC MOLDS 3331
on May 16, 2020 by guest
http://jcm.asm.org/
strain designated the neotype strain for T. rubrum (CBS 392.68). Included in theT. rubrumgroup by Graser et al. (17) is the strain originally designated the type strain for Trichophy-ton fluviomuniense, CBS 592.68, which shows only 98.3% iden-tity to the UWFP clinical isolates and to theT. rubrumneotype strain (CBS 392.68) at ITS1 and ITS2. Demonstration of ⬍99% identity at two diagnostic loci suggests to us that this expandedT. rubrum group comprises at least two species,T. rubrumandT.fluviomuniense.
The methods we describe are rapid. After a mold is cultured on agar medium (72 h), identification can be achieved within 24 h. The 43 recognized species that we analyzed represent 19 genera; 80.3% of all mold isolates (n ⫽ 1,164, isolated in a 12-month period) identified by phenotypic methods in our laboratory belong to the 19 genera tested in this study. These 1,164 strains include 146 Penicillium isolates (12.5% of the total) that were not identified to the species level. An addi-tional 112 isolates of the 1,164 (9.62%) were identified pheno-typically as a mold but were not identified to the genus level. In the first 12 months in which DNA sequence analysis was inte-grated into our clinical laboratory’s identification algorithm, 89 molds were identified by DNA sequence analyses. Isolates were analyzed genotypically if they did not produce reproduc-tive structures within 10 days of isolation or if structures were produced but if phenotypic evaluation did not result in defin-itive identification. Fifty-seven of the 89 isolates produced only sterile hyphae after 10 days and, without DNA sequence anal-yses, would have been reported as M. sterilia or, if possible, more specifically identified after further incubation. Of these 57, 16 were identified to the species level, including three atypical A. fumigatus isolates, and 16 were identified to the genus level. The remaining 25 isolates were assigned a taxo-nomic orientation based on the relationship of their DNA sequences to those of related organisms in the database. Thus, DNA sequence-based methods can identify isolates that oth-erwise defy identification by traditional phenotypic analyses. The integration of multilocus sequence analyses with pheno-type-based identification algorithms in a clinical laboratory provides a rapid and definitive identification that, in some cases, surpasses the specificity of identification by 28S se-quence analyses and phenotypic methods alone (Table 2). We conclude that the phenotypically validated ITS DNA sequence database will be useful for identification of routinely isolated molds, previously unidentified molds, and molds that do not show specific or expected morphological and biochemical phe-notypes.
ACKNOWLEDGMENTS
We thank Jennifer Prentice for her technical assistance and exper-tise and Elaine Brooks for assistance preparing the manuscript.
This publication was made possible in part by grant number 7R44GM57669-04 from the National Institute of General Medical Sciences to Clarity BioSciences, Inc.
The contents of this paper are solely the responsibility of the authors and do not necessarily represent the official views of the NIGMS.
REFERENCES
1.Abliz, P., K. Fukushima, K. Takizawa, and K. Nishimura.2004. Identifica-tion of pathogenic dermatiaceous fungi and related taxa based on large subunit ribosomal DNA D1/D2 domain sequence analysis. FEMS Immunol. Med. Microbiol.40:41–49.
2.Baddley, J. W., T. P. Stroud, D. Salzman, and P. G. Pappas.2001. Invasive
mold infections in allogeneic bone marrow transplant recipients. Clin. Infect. Dis.32:1319–1324.
3.Boysen, M. E., K. G. Jacobsson, and J. Schnurer.2000. Molecular identifi-cation of species from the Penicillium roqueforti group associated with spoiled animal feed. Appl. Environ. Microbiol.66:1523–1526.
4.Campbell, M. C., and J. L. Stewart.1980. The medical mycology handbook. Wiley Medical Publication, New York, N.Y.
5.Chen, X., C. P. Romaine, Q. Tan, B. Schlagnhaufer, M. D. Ospina-Giraldo, D. J. Royse, and D. R. Huff.1999. PCR-based genotyping of epidemic and preepidemicTrichodermaisolates associated with green mold ofAgaricus bisporus. Appl. Environ. Microbiol.65:2674–2678.
6.Chen, Y. C., J. D. Eisner, M. M. Kattar, S. L. Rassoulian-Barrett, K. Lafe, U. Bui, A. P. Limaye, and B. T. Cookson.2001. Polymorphic internal tran-scribed spacer region 1 DNA sequences identify medically important yeasts. J. Clin. Microbiol.39:4042–4051.
7.Chen, Y. C., J. D. Eisner, M. M. Kattar, S. L. Rassoulian-Barrett, K. LaFe, S. L. Yarfitz, A. P. Limaye, and B. T. Cookson. 2000. Identification of medically important yeasts using PCR-based detection of DNA sequence polymorphisms in the internal transcribed spacer 2 region of the rRNA genes. J. Clin. Microbiol.38:2302–2310.
8.De Baere, T., G. Claeys, D. Swinne, G. Verschraegen, A. Muylaert, C. Mas-sonet, and M. Vaneechoutte. 2002. Identification of cultured isolates of clinically important yeast species using fluorescent fragment length analysis of the amplified internally transcribed rRNA spacer 2 region (ITS2). BMC Microbiol.2:21.
9.Denning, D. W., S. A. Radford, K. L. Oakley, L. Hall, E. M. Johnson, and D. W. Warnock.1997. Correlation between in-vitro susceptibility testing to itraconazole and in-vivo outcome ofAspergillus fumigatusinfection. J. Anti-microb. Chemother.40:401–414.
10.Einsele, H., H. Hebart, G. Roller, J. Loffler, I. Rothenhofer, C. Muller, R. Bowden, J. Van Burik, D. Engelhard, L. Kanz, and U. Schumacher.1997. Detection and identification of fungal pathogens in blood by using molecular probes. J. Clin. Microbiol.35:1353–1360.
11.Esteve-Zarzoso, B., C. Belloch, F. Uruburu, and A. Querol.1999. Identifi-cation of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Bacteriol.49:329–337. 12.Ferrer, C., F. Colom, S. Frases, E. Mulet, J. L. Abad, and J. L. Alio.2001.
Detection and identification of fungal pathogens by PCR and by ITS2 and 5.8S ribosomal DNA typing in ocular infections. J. Clin. Microbiol.39:2873– 2879.
13.Fisher, F., and N. A. Cook.1998. Fundamentals of diagnostic mycology. W. B. Saunders, Philadelphia, Pa.
14.Fujita, S. I., Y. Senda, S. Nakaguchi, and T. Hashimoto.2001. Multiplex PCR using internal transcribed spacer 1 and 2 regions for rapid detection and identification of yeast strains. J. Clin. Microbiol.39:3617–3622. 15.Glenn, A. E., C. W. Bacon, R. Price, and R. T. Hanlin.1996. Molecular
phylogeny ofAcremoniumand its taxonomic implications. Mycologia88:369– 383.
16.Graser, Y., M. El Fari, R. Vilgalys, A. F. Kuijpers, G. S. de Hoog, W. Presber, and H. Teitz.1999. Phylogeny and taxonomy of the family Arthrodermata-ceae (dermatophytes) using sequence analysis of the ribosomal ITS region. Med. Mycol.37:105–114.
17.Graser, Y., A. F. Kuijpers, W. Presber, and G. S. de Hoog.2000. Molecular taxonomy of theTrichophyton rubrumcomplex. J. Clin. Microbiol.38:3329– 3336.
18.Groll, A. H., P. M. Shah, C. Mentzel, M. Schneider, G. Just-Nuebling, and K. Huebner.1996. Trends in the postmortem epidemiology of invasive fungal infections at a university hospital. J. Infect.33:23–32.
19.Guarro, J., J. Gene, and A. Stchigel.1999. Developments in fungal taxon-omy. Clin. Microbiol. Rev.12:454–500.
20.Hall, L., S. Wohlfiel, and G. D. Roberts.2003. Experience with the MicroSeq D2 large-subunit ribosomal DNA sequencing kit for identification of com-monly encountered, clinically important yeast species. J. Clin. Microbiol.
41:5099–5102.
21.Hall, L., S. Wohlfiel, and G. D. Roberts.2004. Experience with the MicroSeq D2 large-subunit ribosomal DNA sequencing kit for identification of fila-mentous fungi encountered in the clinical laboratory. J. Clin. Microbiol.
42:622–626.
22.Henry, T., P. C. Iwen, and S. H. Hinrichs.2000. Identification ofAspergillus
species using internal transcribed spacer regions 1 and 2. J. Clin. Microbiol.
38:1510–1515.
23.Hermosa, M. R., I. Grondona, E. A. Iturriaga, J. M. Diaz-Minguez, C. Castro, E. Monte, and I. Garcia-Acha.2000. Molecular characterization and identification of biocontrol isolates ofTrichodermaspp. Appl. Environ. Mi-crobiol.66:1890–1898.
24.Ito, Y., S. W. Peterson, D. T. Wicklow, and T. Goto. 2001.Aspergillus pseudotamarii, a new aflatoxin producing species inAspergillussectionFlavi. Mycol. Res.105:233–239.
25.Kane, J., R. C. Summerbell, L. Sigler, S. Krajden, and G. Land.1997. Laboratory handbook of dermatophytes: a clinical guide and laboratory manual of dermatophytes and other filamentous fungi from skin, hair and nails. Star Publishing Co., Belmont, Calif.
3332 RAKEMAN ET AL. J. CLIN. MICROBIOL.
on May 16, 2020 by guest
http://jcm.asm.org/
26.Kern, M. E.1985. Medical mycology. F. A. Davis Company, Philadelphia, Pa.
27.Klich, M. A., and J. I. Pitt.1988. A laboratory guide to commonAspergillus
species and their teleomorphs. Commonwealth Scientific and Industrial Re-search Organization, Division of Food Processing, North Ryde, New South Wales, Australia.
28.Kurtzman, C. P., and C. J. Robnett.1997. Identification of clinically impor-tant ascomycetous yeasts based on nucleotide divergence in the 5⬘end of the large-subunit (26S) ribosomal DNA gene. J. Clin. Microbiol.35:1216–1223. 29.Larone, D.1995. Medically important fungi: a guide to identification, 3rd ed.
ASM Press, Washington, D.C.
30.Marr, K. A., R. A. Carter, F. Crippa, A. Wald, and L. Corey.2002. Epide-miology and outcome of mould infections in hematopoietic stem cell trans-plant recipients. Clin. Infect. Dis.34:909–917.
31.Motoyama, A. B., E. J. Venancio, G. O. Brandao, S. Petrofeza-Silva, I. S. Pereira, C. M. Soares, and M. S. Felipe.2000. Molecular identification of
Paracoccidioides brasiliensisby PCR amplification of ribosomal DNA. J. Clin. Microbiol.38:3106–3109.
32.Ninet, B., I. Jan, O. Bontems, B. Lechenne, O. Jousson, R. Panizzon, D. Lew, and M. Monod.2003. Identification of dermatophyte species by 28S ribo-somal DNA sequencing with a commercial kit. J. Clin. Microbiol.41:826– 830.
33.Novicki, T. J., R. Geise, A. P. Limaye, K. Lafe, L. Bui, U. Bui, and B. T. Cookson.2003. Genetic diversity among clinical isolates ofAcremonium strictumdetermined during an investigation of a fatal mycosis. J. Clin. Mi-crobiol.41:2623–2628.
34.Page, R. D. M.1996. TREEVIEW: an application to display phylogenetic trees on personal computers. Comput. Appl. Biosci.12:357–358. 35.Person, W. R., T. Wood, Z. Zhang, and W. Miller.1997. Comparison of DNA
sequences with protein sequences. Genomics46:24–36.
36.Peterson, S.2000. Phylogenetic relationships inAspergillusbased on rDNA sequence analysis, p. 323–355.InR. Samson and J. Pitt (ed.), Integration of modern taxonomic methods forPenicillium andAspergillusclassification. Harwood Academic Publishers, Amsterdam, The Netherlands.
37.Raper, K. B., and D. I. Fennell.1965. The genusAspergillus. Robert E. Kreiger Publishing Company, Huntington, N.Y.
38.Selvarangan, R., A. P. Limaye, and B. T. Cookson.2002. Rapid identification and differentiation ofCandida albicansandCandida dubliniensisby capillary-based amplification and fluorescent probe hybridization. J. Clin. Microbiol.
40:4308–4312.
39.Seo, K., H. Akiyoshi, and Y. Ohnishi.1999. Alteration of cell wall compo-sition leads to amphotericin B resistance inAspergillus flavus. Microbiol. Immunol.43:1017–1025.
40.Sharmin, S., K. Haritani, R. Tanaka, P. Abliz, K. Takizawa, A. Sano, K. Fukushima, K. Nishimura, and M. Miyaji.2002. The first isolation of Hor-taea werneckiifrom a household guinea pig. Jpn. J. Med. Mycol.43:175–180.
41.Sigler, L., and M. J. Kennedy.1999.Aspergillus,Fusarium, and other oppor-tunistic moniliaceous fungi, p. 1212–1241.InP. R. Murray, E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (ed.), Manual of clinical microbiology, 7th ed. ASM Press, Washington, D.C.
42.St. Germain, G., and R. C. Summerbell.1996. Identifying filamentous fungi: a clinical laboratory handbook. Star Publishing Company, Belmont, Calif. 43.Sugita, T., A. Nishikawa, R. Ikeda, and T. Shinoda.1999. Identification of
medically relevantTrichosporonspecies based on sequences of internal tran-scribed spacer regions and construction of a database forTrichosporon iden-tification. J. Clin. Microbiol.37:1985–1993.
44.Sutton, D. A., A. W. Fothergill, and M. G. Rinaldi.1998. Guide to clinically significant fungi. Williams and Wilkins, Baltimore, Md.
45.Sutton, D. A., S. E. Sanche, S. G. Revankar, A. W. Fothergill, and M. G. Rinaldi.1999. In vitro amphotericin B resistance in clinical isolates of As-pergillus terreus, with a head-to-head comparison to voriconazole. J. Clin. Microbiol.37:2343–2345.
46.Tamura, M., H. Kawasaki, and J. Sugiyama.1999. Identity of the xerophilic speciesAspergillus penicillioides: integrated analysis of the genotypic and phenotypic characters. J. Gen. Appl. Microbiol.45:29–37.
47.Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G. Higgins.1997. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res.25:4876–4882.
48.Turenne, C. Y., S. E. Sanche, D. J. Hoban, J. A. Karlowsky, and A. M. Kabani.1999. Rapid identification of fungi by using the ITS2 genetic region and an automated fluorescent capillary electrophoresis system. J. Clin. Mi-crobiol.37:1846–1851.
49.Untereiner, W. A., and F. A. Naveau.1999. Molecular systematics of the Herpotrichiellaceae with an assessment of the phylogenetic positions of
Exophiala dermatitidisandPhialophora americana. Mycologia91:67–83. 50.Voigt, K., E. Cigelnik, and K. O’Donnell.1999. Phylogeny and PCR
identi-fication of clinically important zygomycetes based on nuclear ribosomal-DNA sequence data. J. Clin. Microbiol.37:3957–3964.
51.White, T. J., T. Bruns, S. Lee, and J. Taylor.1990. Amplification and direct sequencing of fungal ribosomal RNA sequences for phylogenetics, p. 315– 322.InM. A. Innis, D. H. Gefland, J. J. Sninsky, and T. J. White (ed.), PCR protocols: a guide to methods and applications. Academic Press, Inc., New York, N.Y.
52.Yokoyama, K., L. Wang, M. Miyaji, and K. Nishimura.2001. Identification, classification and phylogeny of theAspergillussection Nigri inferred from mitochondrial cytochrome b gene. FEMS Microbiol. Lett.200:241–246. 53.Zhao, J., F. Kong, R. Li, X. Wang, Z. Wan, and D. Wang.2001. Identification
ofAspergillus fumigatusand related species by nested PCR targeting ribo-somal DNA internal transcribed spacer regions. J. Clin. Microbiol.39:2261– 2266.
VOL. 43, 2005 RAPID IDENTIFICATION OF PATHOGENIC MOLDS 3333