ASSEMBLY AND DISASSEMLY OF THE WNT BETA-CATENIN DESTRUCTION COMPLEX
Kristina Schaefer
A dissertation submitted to the faculty at the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Curriculum of
Genetics and Molecular biology in the School of Medicine.
Chapel Hill 2018
Approved by: Mark Peifer Robert Duronio Amy Shaub Maddox Kevin Slep
iii ABSTRACT
Kristina N. Schaefer: Assembling and Disassembling the Wnt Beta-catenin destruction complex (Under the direction of Mark Peifer)
Wnt signaling provides a key example for cell-cell signaling pathways that regulate embryonic development and stem cell homeostasis and then are frequently inappropriately activated in cancers. Wnt signaling acts by regulating levels of β-catenin (βcat), an essential transcriptional coactivator of Wnt target genes. The tumor suppressors APC and Axin, along with the kinases GSK3 and CK1, form the core of the multiprotein destruction complex (DC), which targets βcat for phosphorylation, ubiquitination and destruction. In the presence of Wnt ligands, DC function is down-regulated, allowing levels of βcat levels to rise, eventually entering the nucleus to activate transcription of Wnt target genes. Based on earlier work, we hypothesize that the DC is a supramolecular entity that self-assembles by Axin and APC polymerization, and that regulation of assembly and stability of the DC regulates its function. We tested this
iv
inactivation. We also found that endogenous Axin and APC2 proteins and their antagonist Dishevelled accumulate at roughly similar levels. Hetero-polymerization between Dishevelled and Axin via their DIX domains is essential for Wnt down regulation of the DC, yet the
v PREFACE
“One, remember to look up at the stars and not down at your feet. Two, never give up work. Work gives you meaning and purpose and life is empty without it. Three, if you are lucky enough
to find love, remember it is there and don't throw it away
.
”Stephan Hawking
We were invited by Developmental Cell to write a review on the current view of Wnt signaling. Therefore, instead of a typical introduction chapter, we wrote a review. This review centered around our hypothesis that the destruction complex is really a biomolecular
condensate. We discuss recent research in the field and describe how several recent reports support our hypothesis. This chapter was a collaboration between Mark and I. We have now submitted the manuscript.
vi
Clara Williams analyzed Engrailed expression, and helped with determining embryonic lethality, terminal cuticle phenotypes, and prepping embryos for imaging analysis. Dave Roberts imaged and analyzed Arm levels in APC2 mutants. Dan McKay analyzed RNAseq data. I helped organize the team of authors on this paper and performed all other experiments. The manuscript was written by me and Mark Peifer with input from the other authors.
Chapter 3 describes my work on how beta-catenin is transferred from the destruction complex to the E3 ligase. The groundwork of this project was started when I was a rotation student, and many tools utilized in this project were created by an undergraduate honors research student Lauren Bauer, who I supervised. At the time of this preface, we still finishing up a few experiments and plan to include this data as part of a manuscript we hope to submit later this year. Mira Pronobis conducted and analyzed the FRAP data. I performed the
vii
ACKNOWLEDGEMENTS
Before I even entered graduated school, I was given a series of article about how difficult it would be, and then I was questioned whether I really wanted to purse my Ph.D. Throughout my graduate school career I have regretted this decision and have been elated I went back to school. I want to thank those who helped me back on the path of science and discovery.
1) To Dr. Xin Zhang, without him I would have never found my love for research or decided to attend graduate school in the first place.
2) To Mark Peifer who pushed me to be a greater thinker, to make a better argument, and to be healthily skeptical about my own data.
3) To my other lab members and peers - thanks for being my sounding board, my late-night confidant, for helping me stay the course, and for getting me out the lab and doing something fun every once and a while.
4) Thanks to my family who have always supported me in any they could. I would not have made it through without you guys.
viii
TABLE OF CONTENTS
LIST OF FIGURES ... xii
LIST OF TABLES... xiv
CHAPTER 1: EMERGING IDEAS IN REGULATING WNT SIGNALING: REGULATION BY POLYMERIZATION AND PARALLEL BEHAVIOR WITH BIOMOLECUAR CONDENSATES .. 1
OVERVIEW: ... 1
The textbook model of Wnt signaling ... 6
The Wnt-regulatory destruction complex—is it a biomolecular condensate? ... 8
The destruction complex is an internally ordered structure that assembles by polymerization ... 10
A functional destruction complex contains many more than four proteins ... 13
destruction ... 15
... 17
Other conserved sequences in APC’s intrinsically disordered region also play key functions... 18
APC may play additional positive and negative roles in Wnt signaling ... 19
Regulating a biomolecular condensate: Wnt signaling changes destruction complex localization and assembly ... 20
Wnt signaling causes a switch in the destruction complex mix, destabilizing it ... 21
One consequence of supermolecular assembly: the kinase GSK3 plays both positive and negative roles in the destruction complex via its access to many targets ... 23
Axin post-translational regulation plays complex roles ... 24
APC mutations in colorectal cancer target specific aspects of destruction complex function ... 27
The destruction complex is a multifunctional machine with other targets including the cytoskeleton ... 28
REFERENCES ... 33
ix
OVERVIEW: ... 46
Author Summary ... 46
INTRODUCTION ... 47
RESULTS ... 52
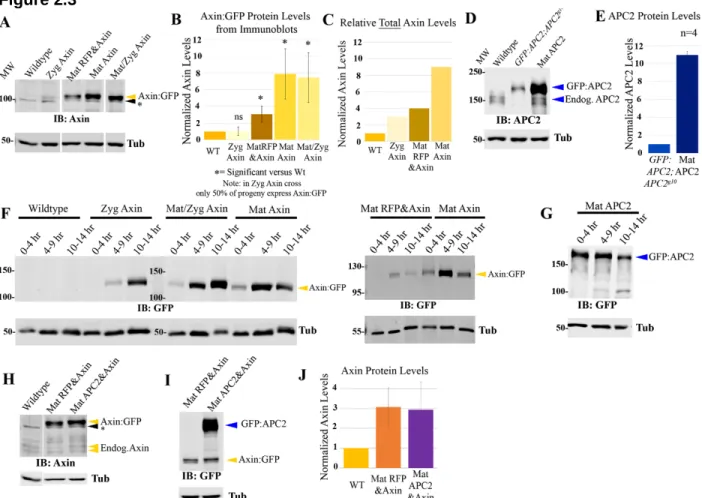
axinandAPC1/APC2are transcribed at similar levels ... 52
Axin and APC2 proteins accumulate at similar levels during early-mid embryogenesis ... 54
-fold, it inhibits Wg-regulated cell fate choice during embryogenesis ... 59
Elevating Axin levels has no effect on Arm levels in cells not receiving Wg signals, but does render the destruction complex more resistant to inactivation by physiological levels of Wg signaling ... 61
Levels of APC2 can be substantially elevated without significantly affecting viability or Wg-regulated cell fates ... 68
Elevating levels of APC2 strongly promotes downregulation of the destruction complex in response to physiological levels of Wg signaling ... 68
Simultaneously elevating levels of both APC2 and Axin inhibits Wg signaling more than elevating levels of Axin alone ... 71
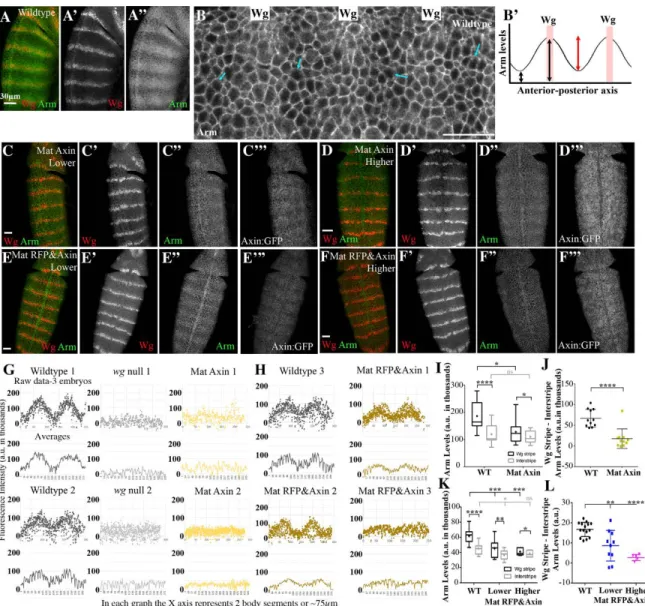
The relative ratio of APC2:Axin levels determines the effectiveness of Arm destruction ... 73
Axin assembles into cytoplasmic multiprotein destruction complexes, and Wnt/Wg signaling leads to their membrane-recruitment and elevates levels of cytoplasmic Axin ... 77
Wg signaling and GSK3 activity are each required for membrane recruitment of Axin puncta ... 86
Simultaneously elevating Axin and APC2 makes destruction complex puncta more resistant to disassembly by Wg signaling ... 89
Each destruction complex punctum includes tens to hundreds of APC2 or Axin proteins ... 90
Dsh accumulates at levels similar to those of APC2 and Axin and localizes to Axin puncta in cells that receive Wg signals ... 94
DISCUSSION ... 98
In vivo levels of APC2 and Axin are similar rather than orders of magnitude different ... 98
x
Wg signaling triggers membrane recruitment of Axin and may destabilize destruction
complex assembly ... 100
Elevating Axin levels renders the destruction complex less sensitive to inactivation by Wg signaling ... 102
APC2 is not rate-limiting for destruction complex activity but elevating its levels facilitates destruction complex inactivation ... 103
Effects of altering the Axin:APC2 ratio suggest APC2 can play both positive and negative roles in Wnt regulation ... 103
A proposed model of how Wnt signaling regulates destruction complex assembly and function ... 106
MATERIALS AND METHODS ... 107
Fly stocks, embryonic lethality, and cuticles ... 107
Immunostaining and antibodies ... 108
Assessing effects on Engrailed expression ... 109
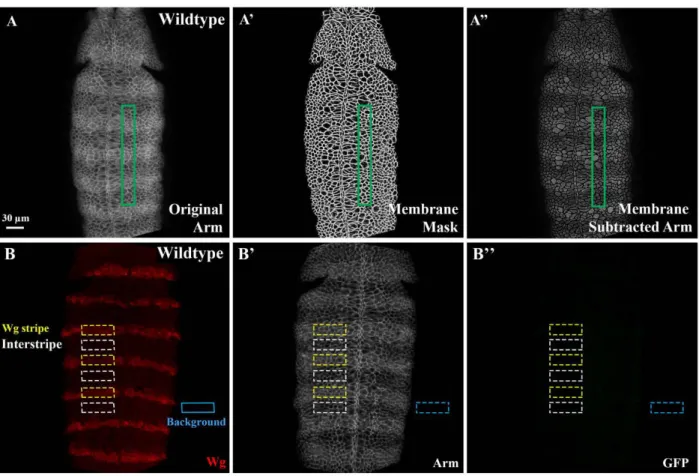
Quantitative analysis of Arm accumulation ... 109
Statistics ... 111
Immunoblotting ... 111
RNA-Seq ... 112
Cell culture and transfections ... 112
Yeast fluorescence comparison ... 112
REFERENCES: ... 122
CHAPTER 3: THE DANCE BETWEEN THE DESTRUCTION COMPLEX AND THE E3 LIGASE ... 128
OVERVIEW: ... 128
INTRODUCTION: ... 128
1. Recruitment ... 129
2.Phosphorylation ... 130
3. Ubiquitination ... 130
4.Degradation ... 131
RESULTS ... 133
A system to examine whether the destruction complex and the E3 ligase co-localize ... 133
Axin and not APC2 can recruit Slimb into the destruction complex... 134
The RGS domain of Axin is required for efficient Slimb recruitment ... 135
xi
Slimb localizes along Axin cables ... 141
DISCUSSION ... 141
Defining the DC and E3 ligase interaction... 141
The APC2:Axin complex recruits the F-box E3 adaptor Slimb ... 143
APC2 and SCFSlimb don’t mix, but Axin does ... 145
The RGS domain of Axin is necessary to efficiently recruit Slimb ... 145
occurs in a supra-molecular factory ... 147
METHODS: ... 149
Cell Culture and transfection ... 149
Immunofluorescence and Microscopy ... 149
Immunoprecipitation and Western blotting ... 149
REFERENCES ... 151
CHAPTER 4: DISCUSSION ... 155
And the debate goes on: Wnt regulation of the destruction complex ... 155
The basics: Wnt signaling pathway simplified ... 155
Size matters: Why is polymerization necessary for complex function? ... 157
Condensates: the more the merrier ... 157
Condensates: A way to bring multiple complexes together? ... 160
Multiple mechanisms of turing down of the destruction complex ... 161
Axin degradation or disassembly: control the scaffold, control the complex? ... 162
Does a change in scenery result in a change in function? ... 163
The destruction complex loses a friend... 165
Dishevelled: The regulator of Axin puncta ... 167
Revised Model: It’s all about who your friends are at the time ... 171
Putting all the pieces together ... 171
Shifting friends ... 172
xii
LIST OF FIGURES
Figure 1.1 - Properties of a biomolecular condensate…...………...…4
Figure 1.2 - The Wnt signaling pathway and APC, Axin and Dvl proteins
contain properties of proteins found in condensates………...………..………...7
Figure 1.3 - Axin and Axin plus APC accumulate in non-membrane
bound puncta…...…….…...….11
Figure 1.4 - In vivo recruitment of APC2 into Axin:GFP puncta……….………....…..22
Figure 1.5 - A revised model of the destruction complex...30
Figure 2.1 - Endogenous APC2 and Axin proteins accumulate at similar
levels…….…...……...53
Figure 2.2 - Crosses used to achieve different level and timing of Axin
elevation...56
Figure 2.3 - Developing tools to differentially elevate levels of Axin:GFP………...…...…....58
Figure 2.4 - Elevating Axin produces dose-sensitive inhibition of Wg
signaling, while increasing APC2 levels does not……….……….60
Figure 2.5 - Increasing Axin levels reduces the ability of endogenous Wg signaling to turn down the destruction complex but has little
or no effect in Wg-Off cells………...………...…63
Figure 2.6 - Assessing gradation of Arm levels across the segment
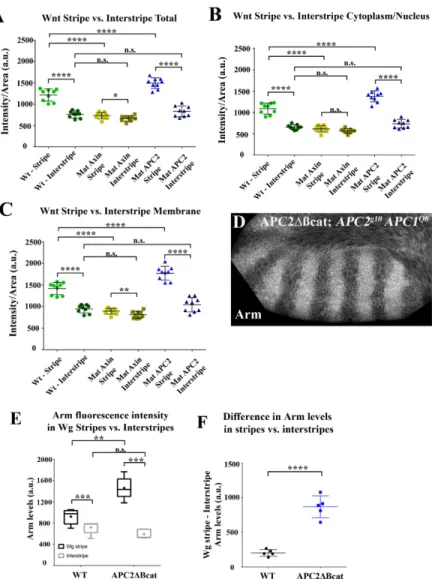
and absolute levels of Arm in Wg stripes and interstripes...66
Figure 2.7 - The opposite effects of Axin versus APC2 overexpression on Arm levels in Wg-ON cells are observed in both the cytoplasmic
and membrane-associated pools. ...67
Figure 2.8 - Elevating APC2 levels increases the ability of endogenous Wg signaling to turn down the destruction complex, thus increasing
Arm levels in cells receiving Wg…………...……...…….70
Figure 2.9 - The relative ratios of APC2 to Axin levels determine effects on
embryonic viability and Wg-regulated cell fates………...72
Figure 2.10 - Illustration of how embryos were sorted as to inferred genotype...75
Figure 2.11 - The relative ratios of APC2 to Axin levels determine effects on
Arm destruction……...………...……….…..76
Figure 2.12 - Axin:GFP can largely restore normal Wnt signaling after
xiii
Figure 2.13: Flag-tagged Axin assembles into puncta indistinguishable
from those assembled by Axin:GFP...80
Figure 2.14 - Axin assembles into cytoplasmic multiprotein destruction complexes together with APC2, and Wg signaling leads to their membrane-recruitment and elevates levels of cytoplasmic Axin………..….…………..…82
Figure 2.15: When Axin is localized using an antibody to the GFP epitope-tag, it emphasizes the elevation in cytoplasmic Axin in Wg-ON cells and de-emphasizes Axin puncta in Wg-OFF cells...85
Figure 2.16 - Wg signal and GSK3/Zw3 activity are important for destruction complex membrane recruitment and GSK3/Zw3 regulates release of Arm from the destruction complex...……...……...87
Figure 2.17 - Ubiquitous expression of Wg increases embryonic lethality and induces a loss of denticle belts, whereas Dsh overexpression has little effect on viability and cuticle phenotype ...88
Figure 2.18 - The destruction complex contains thousands of APC2 or Axin molecules after over-expression in SW480 cells, and 10-100s of Axin molecules in vivo in embryos...92
Figure 2.19 - Dsh accumulates at similar levels to Axin and APC2, and co-localizes with Axin puncta in Wg-ON but not Wg-OFF cells………...95
Figure 3.1 - Axin recruits Slimb into cytoplasmic puncta…………...….136
Figure 3.2 - Axin is unable to recruit Cul1 or SkpA into the complex………...……138
Figure 3.3 - Slimb is not recruited by mito-APC2………...…..140
Figure 3.4 - The RGS domain of Axin is necessary for Slimb recruitment into Axin puncta…...…142
Figure 3.5 - Slimb turnover in the destruction complex is unaffected by co-localization with Axin or Axin and APC2………...144
xiv
LIST OF TABLES
Table 2.1 - Normalized densitometry values...114
Table 2.2 - Embryonic viability...115
Table 2.3 - Embryonic and first instar larva cuticle phenotype...116
Table 2.4 - Rows of En-expressing cells per segment...117
Table 2.5 - Effects on Arm levels of elevating Axin and/or APC2 levels...118
Table 2.6 - Quantification of the differences in Arm levels in Wg-stripe versus interstripe cells...119
Table 2.7 - Quantification of the different pools of Arm levels in Wg-stripe versus interstripe-cells...120
1
CHAPTER 1: EMERGING IDEAS IN REGULATING WNT SIGNALING: REGULATION BY POLYMERIZATION AND PARALLEL BEHAVIOR WITH BIOMOLECUAR CONDENSATES
OVERVIEW:
Wnt/ βCatenin signaling plays key roles in cell fate decisions in embryonic and post
embryonic development across the animal kingdom, and also helps maintain homeostasis in
many tissues. As a result, loss- and gain-of-function mutations in the pathway are found in both
developmental disorders and in many human cancers. In the absence of Wnt ligands, signaling
is kept off by the multiprotein destruction complex, while pathway activation requires the
destruction complex to be downregulated. Here we describe recent advances in the field that
have provided new insights into the activity of the destruction complex and the mechanisms of
its downregulation and point out parallels to other cell biological processes carried out by
biomolecular condensates that form by phase separation.
The transformation of a fertilized egg into the body of an animal is among the most
remarkable events in biology. Individual cells must choose fates based on their position, and
then maintain those fates for a lifetime through tissue homeostasis. Cell-cell communication is
critical for this, and a handful of cell-cell signaling pathways play especially important roles.
Among these is the Wnt pathway (Nusse and Clevers, 2017), which directs cell fates from the
initial establishment of the vertebrate body axes to the detailed architecture of the kidney or
nervous system. The key developmental roles of these pathways mean that mutations in
2
bone density and growth disorders (e.g. Robinow Disease) and progressive vision loss (Familial
exudative vitreoretinopathy).
The same signaling pathways play critical roles in tissue homeostasis, maintaining proper
cell numbers by regulating tissue stem cell proliferation. To ensure signaling occurs only at the
right time and place, dedicated negative regulatory machinery has evolved to keep signaling
completely off in the absence of ligand. In the Wnt pathway this is accomplished by the
destruction complex, a multiprotein machine that targets the key Wnt-effector beta-catenin
(βCat) for phosphorylation, and ultimate ubiquitination and destruction. Mutations in destruction
complex proteins like Adenomatous polyposis coli (APC) occur in a wide variety of cancers and
play the initiating role in virtually all colorectal tumors. As a result, mechanisms by which Wnt
signaling is regulated are the subject of intensive research, potentially providing new cancer
therapies. Here we summarize current knowledge about Wnt signaling, framing it in the context
of the emerging idea that many key cellular processes are carried out in large non-membrane
bound cellular compartments, an idea we think provides new insights into destruction complex
function and regulation. Due to space limitations, we focus on canonical Wnt signaling, not its
variants, and on its core conserved components; other proteins with tissue- or animal
phyla-specific roles will be neglected, though they are important for a full picture of Wnt signaling and
its regulation (e.g. (Adler and Wallingford, 2017; Green et al., 2014; Malinauskas and Jones,
2014).
Centralized cellular boutiques: biomolecular condensates create cellular signaling and regulatory compartments
The cell is a complex place. Like a city, within its boundaries hundreds of different activities
occur simultaneously at different places, from transcription to translation to metabolism to
transport to cell signaling. To organize this complexity, cells dedicate particular locations to
3
compartments, ranging from the ER or Golgi to the smallest exocytic vesicle. Within them
contents are segregated from the bulk cytoplasm and interchange occurs via specialized
transport systems. However, relying on specialized transport is insufficient to organize the vast
volume of cytoplasm and nucleoplasm not encompassed within a membrane-bound organelle.
To solve this problem, cells evolved an additional mechanism of organizing cellular
compartments that does not require membrane enclosure. Some of these structures were large
enough to merit recognition by cell biology’s pioneers (Gall, 2000)—e.g., nucleoli or Cajal
bodies, locations of ribosome or spliceosome assembly within nuclei, or the germplasm of
animal eggs, containing determinants specifying germ cell fate.
In the past decade scientists recognized that these entities are examples of a much broader
group of non-membrane bound cellular compartments that organize specific proteins and RNAs.
They are key to diverse cellular processes including transcription, the DNA damage response,
and cellular signaling (Banani et al., 2017; Holehouse and Pappu, 2018). Pioneering work on
the C. elegans germline P granules and on signaling centers organized by SH3 domain proteins
led to the idea that these structures assemble by “liquid-liquid phase separation” (Brangwynne
et al., 2009; Li et al., 2012a). Multivalent interactions among their protein and/or RNA
constituents lead to self-assembly, creating compartments separated from the bulk cytoplasm
where the concentration of key players is exceptionally high, significantly speeding intricate
reactions and/or processes (reviewed in Banani et al., 2017). The field emerged from concepts
from soft-matter physics and polymer chemistry, which provide a biophysical basis and
theoretical framework for this behavior. Critically, molecules can freely diffuse within, into and
out of these structures, as they are not enclosed in a lipid bilayer and are often liquid-like in
nature. This allows them to serve as centralized functional hubs for particular cellular processes,
4
Figure 1.1
5
leave. They also serve as storage depots for key players to be deployed at later times.
Structures like these recently were given the broad name “biomolecular condensates”,
reflecting the broad range of cellular and molecular processes that occur within them.
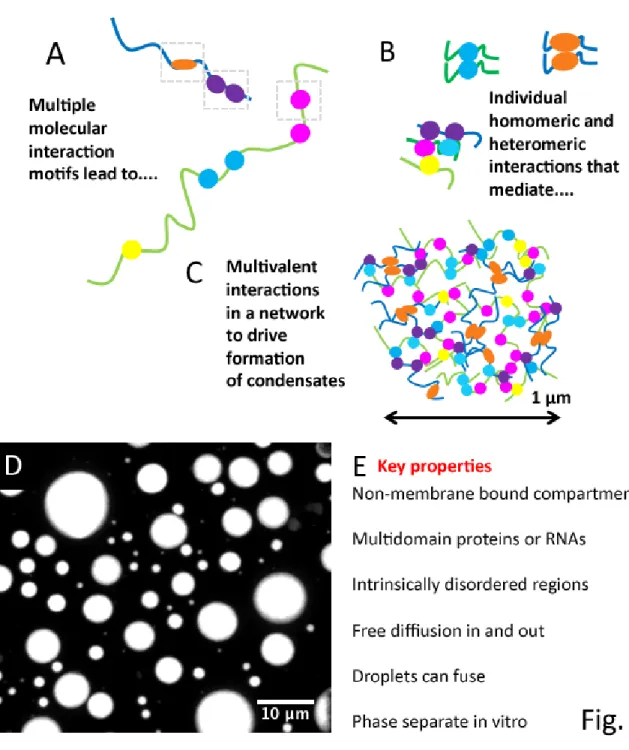
Condensates have a number of defining properties (Banani et al., 2017; Fig. 1.1), though
precise definitions are still being established. Each is a non-membrane bounded structure
ranging up to micron scale that concentrates proteins and/or RNAs at a particular cellular
site. They assemble by multivalent interactions mediated by multidomain proteinsand/or
RNAs with multiple protein or RNA interaction sites (Fig. 1.1). Many of the proteins involved
contain “intrinsically disordered regions (IDRs)” -- these lack tertiary structure, are often not
highly conserved in sequence, and self-interact or include within them interaction sites for other
proteins (Fig. 1.1A-B). IDRs are often tethered to folded domains (Mittal et al., 2018). Even after
phase separation, protein components freely diffuse into and out of the structures. Some
condensates can transition to a more gel-like state (Wang et al., 2018), with reduced exchange
with the bulk cytosol, a process that can contribute both to function and to pathogenesis. One
key to understanding assembly of condensates is the ability to reconstitute phase separation
behavior in vitro, with minimal components (Fig. 1.1D). Both in vitro and in vivo, liquid
condensates can fuse and relax to minimize surface tension. The rapidly expanding universe
of biological processes and structures encompassed under the biomolecular condensate
umbrella and the challenge of defining the rules governing their assembly, disassembly, and
function have made this one of the fastest growing areas of cell biology. As we’ll see below,
structures that regulate and transduce Wnt signals share many features with biomolecular
6
The textbook model of Wnt signaling
Like most key signaling pathways regulating development, the primary output of the
canonical Wnt pathway is a change in the cell’s transcriptional program. This occurs by
regulating levels of βCat, a co-activator of transcription (reviewed in Gammons and Bienz, 2017;
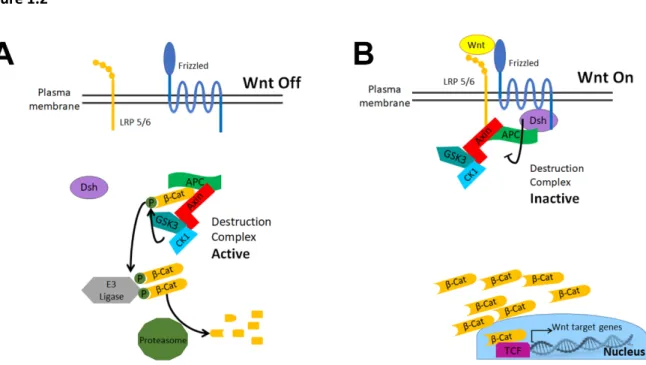
Nusse and Clevers, 2017; Stamos and Weis, 2013). In the absence of Wnt signaling βCat levels
are kept low by the βCat destruction complex (Fig. 1.2A). At the core of this complex are the
tumor suppressor APC, the scaffold Axin, and two kinases, GSK3 and CK1. This complex
recruits βCat, where it is sequentially phosphorylated by CK1 and then GSK3. Once βCat is
phosphorylated, it is transferred to the Cullin-based E3 Ligase SCFβTrCP, polyubiqitinated, and
then recognized by the proteasome and degraded. As a result, Wnt-regulated transcription is
OFF.
Wnt ligands bind both the 7 transmembrane Frizzled (Fz) and the single-pass
transmembrane LRP5/6 receptors (Fig. 1.2B; reviewed in DeBruine et al., 2017; Nusse and
Clevers, 2017). The Wnt/Fz/LRP complex recruits the cytoplasmic protein Disheveled (Dvl in
mammals, fly Dsh). GSK3 phosphorylates LRP5/6 (fly Arrow), creating a binding site for Axin
and recruiting it to the membrane. This downregulates destruction complex activity. The primary
mechanism of downregulation is not yet clear, as data support diverse mechanisms ranging
from disassembly of the complex, inhibition of GSK3 kinase activity, Axin degradation,
sequestration of destruction complex core proteins, and loss of E3-ligase interaction.
Destruction complex inhibition allows βCat levels to rise and it enters the nucleus, binding to T
-Cell Factor (TCF)/Lymphoid Enhancer factor (LEF) family DNA binding proteins. TCF/LEF
proteins bind Wnt regulated genes, initiating different multiprotein complexes in cells where Wnt
signaling is ON or OFF (reviewed by Gammons and Bienz, 2017). Thus the ultimate output of
Wnt signaling occurs when the TCF/LEF:βCat complex activates transcription of Wnt target
7
Figure 1.2
Figure 1.2: Textbook model of Wnt signaling. A) In the absence of a Wnt ligand, the
destruction complex (APC, Axin, CK1, GSK3) recruits βcat for phosphorylation. Once
phosphorylated, βcat can be recognized by the E3 ligase, SCFβTrCP, ubiquitinated, and then
passed to the proteasome for ultimate protein degradation. B) Wnt signaling induces the formation of the Wnt receptor complex of Wnt/Frizzled/LRP5/6/Dvl. This complex recruits Axin
and induces down-regulation of the destruction complex. Levels of cytoplasmic βcat rise,
8
Although the field agrees on the main components of the destruction complex, key
questions about its function and regulation remain unanswered. For example, what is the role of
APC within the destruction complex? The fact that APC is mutated in >80% of colon cancers
emphasizes that it is essential for destruction complex function (Kandoth et al., 2013), yet what
APC does within the complex remained unclear. Second, Axin has a self-polymerizing domain,
and loss of this domain reduces destruction complex function. This suggests that the destruction
complex is larger than a simple hetero-tetramer of Axin, APC and the two kinases, but how does
the sum of these parts create and affect complex function? Another key question involves the
primary mechanism used to down regulate destruction complex function. A few prominent
theories were mentioned above, but which is the primary mechanism? New research is
shedding light on these and other areas, providing insights into the mechanisms of destruction
complex function and regulation.
The Wnt-regulatory destruction complex—is it a biomolecular condensate?
Looking back with hindsight at our unfolding understanding of the components, regulation
and function of the destruction complex reveals striking parallels between its properties and
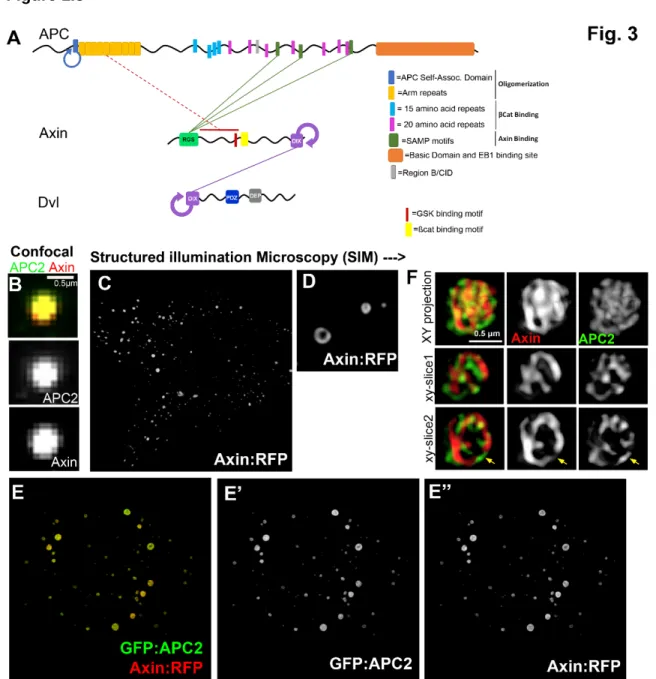
many of those of biomolecular condensates. Two key non-enzymatic components, APC and
Axin, are complex multidomain scaffolding proteins containing folded domains that bind
other proteins along with long intrinsically disordered regions that contain binding sites for
other destruction complex proteins, including βCat (Fig. 1.3A, reviewed in Stamos and Weis,
2013). For example, human APC is predicted to have 50% disordered content (Piovesan et al.,
2018). Axin has an N-terminal Regulator of G-protein signaling (RGS) domain that binds APC, a
C-terminal DIX domain that mediates head-to-tail polymerization, and an intervening intrinsically
disordered region containing binding sites for βCat and the two key kinases, as well as for the
phosphatase PP2A (Behrens et al., 1998; Fagotto et al., 1999; Hart et al., 1998; Ikeda et al.,
9
Williams, 1999; Zeng et al., 1997; Fig.1.3A). Axin’s multiple binding sites allow it to bring βCat
into proximity to the kinases GSK3 and CK1. APC has an conserved N-terminal region that
mediates oligomerization (Kunttas-Tatli et al., 2014) and includes an Armadillo (Arm) repeat
domain that can bind diverse partners. This is followed by a long intrinsically disordered region,
embedded within which are multiple copies of two distinct types of binding sites for βCat (Fig.
3A), multiple binding sites for Axin, and other conserved sites for which the binding partners
remain undetermined (reviewed in Stamos and Weis, 2013). The multivalent nature of APC
and Axin and their intrinsically disordered regions are shared features with known
components of biomolecular condensates, suggesting that they may also form a condensate.
The localization of key destruction complex players is also striking when considered through
the lens of phase separation. When Axin is expressed in many different cell types, both in vitro
and in vivo, it forms large protein “puncta”, and recruits into them APC and other
destruction complex proteins, thus increasing their effective local concentrations (Figs.
1.3 and 1.4, e.g. Cliffe et al., 2003; Fagotto et al., 1999; Faux et al., 2008; Thorvaldsen et al.,
2015). Recent analysis by correlative fluorescence and electron microcopy confirmed these
puncta are not enclosed in a membrane (Thorvaldsen et al., 2015). When Axin is expressed
at endogenous levels, puncta are also seen (Faux et al., 2008). As we discuss in detail below,
APC is required for puncta assembly in vivo, and puncta localization is regulated by Wnt
signaling, consistent with puncta as active players in Wnt regulation (Cliffe et al., 2003;
Mendoza-Topaz et al., 2011; Schaefer et al., 2018). Puncta formation and destruction complex
function depend at least in part on the ability of Axin’s C-terminal DIX domain to oligomerize
(Sakanaka and Williams, 1999). The DIX/DAX (Dishevelled/Axin; referred to as DIX below)
domain was initially defined because it is conserved with Dvl, a positive effector of Wnt
signaling. Dvl also forms puncta, both when expressed in cells (Axelrod et al., 1998;
10
formation also depends on its DIX domain (Schwarz-Romond et al., 2005). Strikingly, Dvl and
Axin physically interact and co-localize in puncta (Fig. 3A) (Fagotto et al., 1999; Julius et al.,
2000; Kishida et al., 1999). Dvl puncta are recruited to the membrane by the Wnt receptor
(Axelrod et al., 1998; Miller et al., 1999; Yang-Snyder et al., 1996). Early work suggested that
Dvl’s DIX domain associated with vesicles (Capelluto et al., 2002). However, subsequent work
failed to reveal any co-localization of Dvl with vesicular markers. Instead, live imaging revealed
that Dvl puncta are protein oligomers that can grow by fusion (Schwarz-Romond et al.,
2005). Puncta containing Axin and APC can also fuse (Kunttas-Tatli et al., 2014). FRAP
analysis further revealed that Dvl, Axin, and APC all freelydiffuse into and out of puncta
(Pronobis et al., 2015; Schwarz-Romond et al., 2007b). Together, these data suggest that Dvl
and Axin puncta meet most of the criteria for biomolecular condensates (Fig. 3A) and
demonstrate that puncta assembly is key for destruction complex function and regulation.
The destruction complex is an internally ordered structure that assembles by polymerization
Some biomolecular condensates form via a network of multivalent interactions without a
strong underlying structural scaffold, while others can assemble into a more gel-like polymerized
state (Banani et al., 2017). Early studies of the destruction complex suggested it is more
structured, supporting a model of ‘signaling by reversible polymerization’ (Schwarz-Romond et
al., 2007a). The DIX domains of Dvl and Axin polymerize by head to tail interactions, forming
filaments that can be visualized by EM or X-ray crystallography (Schwarz-Romond et al.,
2007a), similar to tubulin, actin, and septins. Critically, mutations in their respective DIX
domains that block polymerization reduce Dvl’s ability to promote Wnt signaling and attenuate
Axin’s ability to inhibit Wnt signaling (Fiedler et al., 2011; Schwarz-Romond et al., 2007a).
These data suggested that destruction complex puncta have an internal structure conferred by
11
Figure 1.3
Figure 1.3: APC, Axin, and Dvl are multidomain proteins with intrinsically disordered regions that accumulate in structured non-membrane bound puncta. A) Cartoon of the structures of APC, Axin and Dvl highlighting domains mediating self-interaction as well as interaction sites with other proteins. Proteins found in condensates often have intrinsically disordered regions (black lines) and can polymerize and/or oligomerize other components in the condensates. Domains and motifs are as labelled. Solid green lines indicate direct interactions. Dotted orange line indicates identified interaction regions. Circled arrows = regions of self-interaction. B-D. Drosophila APC2 and Axin constructs expressed in SW480 cells. A) Close up of APC and Axin puncta visualized using standard confocal microscopy shows colocalization, but no underlining structure (Image originally published in Pronobis et al., 2015: DOI:
Co-12
transfection of APC2 and Axin. They accumulate together in puncta. Note that puncta are larger and fewer than in B. E) SIM images of a punctum, revealing that APC2 and Axin each form intertwined cables with multiple potential interaction sites, thus revealing the internal structure of the destruction complex. (Image originally published in Pronobis et al., 2015.: DOI:
13
resolution microscopy. Structured illumination super-resolution microscopy (SIM) of Drosophila
Axin and APC2 expressed in SW480 cells resolved the puncta into structured entities (Fig.
1.3B-F) (Pronobis et al., 2015). When expressed alone, Axin formed puncta with an internal
structure that resembled toroids or knots, potentially representing DIX domain filaments (Fig.
1.3C-D). Co-expressing Axin and APC2 led to their co-recruitment in puncta and puncta
assembly was enhanced, with the largest puncta on the order of micron size. Strikingly,
intertwined homo-filaments of Axin and APC2 were resolved, further supporting the idea that
regulated polymerization underlies destruction complex assembly (Fig. 1.3E-F). In parallel,
scientists examined endogenous Axin puncta stabilized by inhibiting the enzyme Tankyrase, a
known regulator of Axin levels, using both SIM and correlative fluorescence and electron
microscopy (Thorvaldsen et al., 2015). These data also revealed micron scale puncta in which
Axin, βCat, and Tankyrase formed an intermeshed network of filaments, which electron
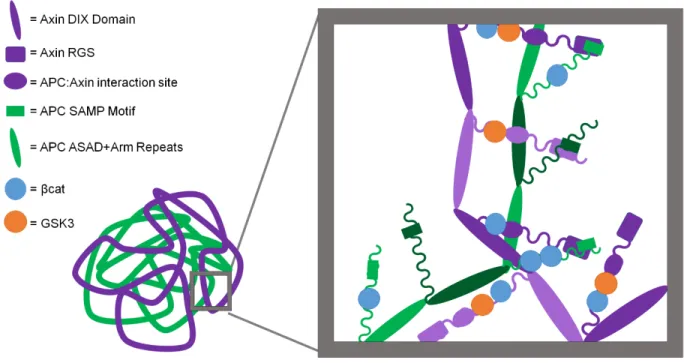
microscopy verified are not membrane-bounded. Together, these data suggest that the
destruction complex has a structured scaffold, built around intertwined polymers of Axin and
APC (Fig. 1.5).
A functional destruction complex contains many more than four proteins
Early work in Xenopus oocytes suggested that destruction complex assembly is limited by
Axin’s very low protein abundance compared to all other destruction complex proteins—up to
5000-fold lower (Lee et al., 2003; Salic et al., 2000). This suggested that Axin was exquisitely
rate-limiting, a factor built into many mathematical models of signaling (e.g.Lee et al., 2003).
Interestingly, recent work in both flies and mammals suggest Axin levels are in fact quite similar
to those of APC (Kitazawa et al., 2017; Schaefer et al., 2018; Tan et al., 2012). These new data
will power updated models and perhaps new insights of Wnt signaling and its regulation.
For decades textbook models of the destruction complex represented it as a simple
14
Axin polymerization is necessary for efficient βCat regulation. Defining the number of molecules
in a functional destruction complex has been a challenge. Some destruction complex proteins
localize to other locations where they have distinct roles --e.g., GSK3 regulates Wnt, Hedgehog,
Insulin, PI3K, and Erk signaling (Cormier and Woodgett, 2017) and APC regulates the
cytoskeleton (Nelson and Nathke, 2013). Thus not every molecule of GSK3 or APC in the cell
localizes to the destruction complex. Second, effective antibodies to key players were not
available. This was particularly true for Axin. To overcome the lack of antibodies to endogenous
Axin, investigators over-expressed Axin and/or APC, hoping the larger complexes formed would
serve as expanded representations of endogenous complexes. This provided important insights.
However, new reagents recently allowed scientists to look at Axin expressed at endogenous or
near endogenous levels, using either a new antibody against fly Axin (Wang et al., 2016a) or
epitope-tagged Axin expressed at near-endogenous levels (Schaefer et al., 2018; Wang et al.,
2016a; Yang et al., 2016). This revealed that in Wnt-OFF cells Axin assembles into puncta
similar to those that assemble after overexpression in cultured cells, and these puncta recruit
APC (Schaefer et al., 2018). This validates early work examining endogenous Axin in MDCK
cells (Faux et al., 2008) and supports the idea that the destruction complex is a supermolecular
machine containing tens to hundreds of molecules rather than a simple 4-protein complex. But
how large is this complex? The ability to express GFP-tagged Axin at near endogenous levels
provided insight. Fluorescence intensity measurements compared to GFP-labeled complexes of
known molecular composition revealed that in Drosophila embryos, active destruction complex
puncta contain on average ~260 Axin molecules (range ~60-930; Schaefer et al., 2018). Mass
spectroscopy provides an alternate mechanism of putting numbers on destruction complex
proteins—recent analyses suggest HEK293 cells each contain ~13,000 Axin proteins (Kitazawa
et al., 2017)—however, with current technology this assessment requires many simplifying
15
machine, consistent with it being a form of biomolecular condensate.
Like many biomolecular condensates, the destruction complex assembles via many
multivalent interactions. This makes it surprisingly robust to removal of some but not all protein
interaction motifs. For example, individually deleting most of APC’s βCat binding sites, Axin’s
βCat binding site, or Axin’s RGS domain have only modest effects in vivo (Kremer et al., 2010;
Kunttas-Tatli et al., 2012; Peterson-Nedry et al., 2008; Roberts et al., 2011; Yamulla et al.,
2014). However, some binding sites are essential individually (Axin’s GSK3 binding site) or
when deleted in concert (Axin∆RGS∆Arm; Peterson-Nedry et al., 2008). This powered synthetic
biology approaches to design a “minimal βCat destruction machine” which retains function in
colorectal cancer cells (Pronobis et al., 2017).
Stabilizing destruction complex supermolecular assembly is a key factor in βCat destruction
Axin’s DIX domain is necessary for its self-polymerization but how is polymerization
regulated? Newly synthesized Axin molecules can either nucleate a new Axin filament or add to
an existing polymer. Several studies focused on APC’s role in forming or stabilizing Axin
filaments. APC is required for assembly of Axin puncta and therefore active destruction
complexes (Mendoza-Topaz et al., 2011). Co-expression revealed that APC2 stabilizes Axin
assembly, as measured by destruction complex volume, increased complexity of Axin filaments,
and decreased Axin turnover (Pronobis et al., 2015). Strikingly, as APC co-expression
increased the size of Axin puncta, it simultaneously decreased the number of puncta, consistent
with the idea that APC2 promotes Axin addition to existing polymers over nucleation of new
polymers (Pronobis et al., 2015).
When the destruction complex was visualized, both Axin and APC2 appeared as intertwined
filaments, suggesting that APC may also polymerize (Fig. 3E-F). Human APC1 has an
16
Drosophila family members. However, another N-terminal region of APC, the APC
self-association domain (ASAD), is conserved in Drosophila, Xenopus, and humans, mediates
Drosophila APC2 self-association (Kunttas-Tatli et al., 2014), and together with APC’s Arm
repeats can mediate puncta formation (Pronobis et al., 2015). The ASAD and adjacent Arm
repeats are required for destruction complex function (Kunttas-Tatli et al., 2014; McCartney et
al., 2006; Roberts et al., 2012a). Interestingly, loss of APC’s oligomerization domain eliminated
APC’s ability to stabilize Axin within the destruction complex (Kunttas-Tatli et al., 2014; Pronobis
et al., 2015). These data suggest that APC polymerization is required to initiate and stabilize
formation of functional destruction complexes.
APC’s stabilization of the destruction complex requires two different Axin:APC interactions
mediated by different domains (Fig.3A; Pronobis et al., 2015). The first is via the
well-established interaction of the Axin-RGS domain with APC’s SAMPS (Spink et al., 2000).
Retention of at least one SAMP is essential for APC function in both mice (Smits et al., 1999)
and flies (Roberts et al., 2011). The second APC:Axin interaction is between APC’s Arm repeats
and a less well-defined region in Axin’s central IDR? intrinsically disordered region (Pronobis et
al., 2015). Recent data suggest that not all SAMP motifs are functionally similar. Fly APC2 has 2
SAMPs, and data suggest one recruits Axin while the other aids in efficient βCat destruction by
an unknown mechanism (Kunttas-Tatli et al., 2015). Intriguingly, in colorectal cancer cells the
SAMP interaction is dispensable if APC and Axin are fused into a single minimal polypeptide
(Pronobis et al., 2017). The ability of APC to stabilize Axin in the destruction complex is further
enhanced by a bridging interaction involving βCat’s ability to bind both APC and Axin, an
interaction disrupted by the vertebrate-specific βCat binding protein ICAT (Ji et al., 2018).
Together, these data support the idea that one key role of APC in the destruction complex is to
17
The destruction complex serves a second role as a sink for cytoplasmic βCat
Another mystery with regard to APC is the role of its multiple βCat binding motifs (Fig. 3A).
Most APC proteins have multiple copies of two distinct types of binding sites for βCat embedded
in the central intrinsically disordered region, the 15- and 20-amino acid repeats (15R and
20R;Eklof Spink et al., 2001; Ha et al., 2004; Liu et al., 2006; Xing et al., 2004). Each 20R has a
different affinity for βCat, with an affinity range of 100-fold (Liu et al., 2006). Phosphorylation of
20Rs by GSK3 dramatically increases their affinity for βCat (Ha et al., 2004; Liu et al., 2006).
This led to the hypothesis that high affinity binding sites are needed when βCat levels are low
and the others come into use when βCat levels are high, helping sequester βCat in the
cytoplasm (Ha et al., 2004; Krieghoff et al., 2006). This model was tested in colorectal cancer
cells and in Drosophila, by systematically deleting 15R and 20R βCat binding sites. Strikingly,
the highest affinity βCat binding sites are dispensable in targeting βCat for destruction—instead
the binding sites collaborate to fine-tune Wnt signals in an additive fashion by cytoplasmic
retention of βCat, supporting the sequestration hypothesis (Kunttas-Tatli et al., 2012; Roberts et
al., 2011; Yamulla et al., 2014). Interestingly, a fly APC2 mutant lacking all the 15R and 20Rs
retains ability to restore APC function in colorectal cancer cells, although it is not is not fully
functional in destruction in vivo in Drosophila (Yamulla et al., 2014). Axin also plays a role in
cytoplasmic retention of βCat in Drosophila (Tolwinski and Wieschaus, 2001).The single βCat
binding site in Axin’s intrinsically disordered region (Xing et al., 2003) may serve a redundant
role, as a designed synthetic minimal destruction complex containing essential regions of APC
and Axin that restored βCat regulation in colorectal cancer cells solely utilized Axin’s βCat
binding site (Pronobis et al., 2017). In vivo analysis of DrosophilaAPC2 mutants lacking βCat
binding sites revealed an aspect of in vivo regulation that remains to be understood—rather
than restoring the graded levels of βCat seen in wildtype, they led to a sharp ON/OFF transition
18
(Orsulic and Peifer, 1996). This is consistent with some sort of threshold feedback response.
Other conserved sequences in APC’s intrinsically disordered region also play key functions
In addition to the βCat and Axin binding sites, APC’s intrinsically disordered region also
contains another highly conserved motif which did not have a known binding partner, variously
called conserved sequence B (B) or the catenin inhibitory domain (CID, Fig. 3A). Strikingly the
B/CID motif is essential for APC function in Wnt regulation in both flies and mammals (Kohler et
al., 2009; Roberts et al., 2011) and may be the sequence targeted for removal in the protein
truncations found in colorectal tumors (Kohler et al., 2009). Intriguingly, an immediately adjacent
motif, 20R2, which resembles other 20Rs but lacks key residues that mediate binding to βCat
(Kohler et al., 2008; Liu et al., 2006), is also essential for Wnt regulation. Together, B and 20R2
may form a binding site for an unidentified partner. Further examination revealed that 20R2/B
regulate one of the two APC:Axin binding interactions, that between APC’s Arm repeats and
Axin’s mid-region. The function of 20R2/B requires phosphorylation by GSK3 (Pronobis et al.,
2015). Together, these data led to a model in which phosphorylation of the B and 20R2 motifs
triggers a conformational change in APC, releasing one of the two APC;Axin interactions and
allowing transfer of phosphorylated βCat to the E3 ubiquitin ligase, as part of a catalytic cycle.
This model is consistent with other data, revealing that loss of GSK3 in fly embryos leads to
βCat accumulation in the destruction complex (Schaefer et al., 2018), and suggesting that
inhibiting βCat release to the E3 ligase is a key step by which Wnt signaling inactivates the
destruction complex (Li et al., 2012b). This model also helps explain a paradox in the field—
colorectal cancer cells are defective in βCat destruction but not in βCat phosphorylation (Yang
et al., 2006).
These data left open the identity of the interacting partner of conserved region B and/or
19
(αcat; Rubinfeld et al., 1993), another component of cadherin-based cell-cell junctions, but the
function of the APC:αcat interaction remained a mystery. In 2013 evidence emerged that αcat
binds to the B/CID region, and assays in cultured cells supported the idea that αcat facilitates
βCat ubiquitination and proteolysis (Choi et al., 2013). These data further suggested that αcat
binds APC via its VH1 domain and that αcat/βCat interaction is also critical for βCat destruction.
These data are intriguing, but the physiological role of αcat in Wnt regulation remains in
question. In Drosophilaneither zygotic αcat mutants (Desai et al., 2013; Sarpal et al., 2012) nor
zygotic βCat mutants deleting the αcat binding site (Orsulic and Peifer, 1996) have defects in
Wnt signaling or its regulation. Mutations in αcat in C. elegans also do not cause obvious
defects in Wnt signaling (Costa et al., 1998). A role for αcat in transcriptional regulation of Wnt
target genes has also been proposed, supported by mass spectrometry suggesting αcat forms a
complex with TCF/LEF family members (Choi et al., 2013). Similar nuclear roles have been
suggested for APC (e.g. Sierra et al., 2006), but sequestering APC at a variety of cytoplasmic
locations does not disrupt regulation of βCat destruction in flies or mammalian cells (Roberts et
al., 2012b) suggesting a nuclear role of APC is not essential. Continued work is needed to
further clarify the relevant binding partner of B/20R2 and its function in destruction complex
function.
APC may play additional positive and negative roles in Wnt signaling
Current data support roles for APC in stabilizing the destruction complex, promoting transfer
of βCat to the E3 ligase, and sequestering βCat in the cytoplasm. However, these may not
encompass its full range of functions. Recent work suggested that APC also acts at the level of
the Wnt receptor, inhibiting baseline activity in the absence of Wnt ligands by promoting
clathrin-dependent receptor endocytosis (Saito-Diaz et al., 2018)—however, this role is only exhibited
by certain APC family members. Another intriguing hypothesis is that APC both inhibits and
20
defining a positive regulatory role of APC is in designing experiments that allow it to be
distinguished from its essential negative regulatory role. Genetic studies in which the levels of
both Drosophila APC proteins, APC1 and APC2, were manipulated in parallel provided the right
system. For example, reducing APC2 function attenuated activated Wnt signaling in fly eyes
induced by loss of APC1 (Takacs et al., 2008). The mechanism by which this occurs was not
fully defined, though effects on Axin stability and phosphorylation were suggested
(Tacchelly-Benites et al., 2018; Takacs et al., 2008; Wang et al., 2016a). Interestingly, over-expressing
APC2 in Drosophilaembryos enhanced accumulation of βCat only in Wnt-ON cells, supporting
the idea that APC2 can aid in turning down of the destruction complex activity in response to
Wnt signals (Schaefer et al., 2018). These data further illustrate the intricate nature of Wnt
regulation.
Regulating a biomolecular condensate: Wnt signaling changes destruction complex localization and assembly
The data above summarize our knowledge of active destruction complexes. Another
challenge is to define how its function is down-regulated by Wnt signaling. Many of us initially
spoke of turning the destruction complex “OFF”, but this is inaccurate. Wnt signaling does not
fully inactivate the complex—it retains, at least initially, the ability to phosphorylate βCat (Kim et
al., 2013; Li et al., 2012b), such that rate of βCat turnover is reduced but not halted (Hernandez
et al., 2012). In retrospect, this was apparent in early work in Drosophila, as mutational
inactivation of GSK3 or APC function led to much higher levels of βCat accumulation than those
seen in cells receiving Wnt signals (e.g. Ahmed et al., 2002; Akong et al., 2002). How is
destruction complex activity repressed? After Wnt ligands bind their receptors, Axin is recruited
into a second protein complex with many properties of a biomolecular condensate, the
“signalasome” (reviewed in Gammons and Bienz, 2017). Axin recruitment to the Fz:LRP
21
et al., 2005; Mao et al., 2001; Tamai et al., 2004; Zeng et al., 2005) and by a less well defined
role of Dvl (Bilic et al., 2007; Cliffe et al., 2003). Consistent with this, Drosophila Axin (at
endogenous or near endogenous levels) is found in cytoplasmic puncta in the absence of Wnt
signaling, while in cells receiving Wnt signals Axin puncta are recruited to the plasma
membrane (Schaefer et al., 2018). Interestingly, in Wnt receiving cells, the number of Axin
molecules in puncta is reduced (Schaefer et al., 2018) while the cytoplasmic Axin pool is
increased (Schaefer et al., 2018; Wang et al., 2016a; Yang et al., 2016)) suggesting that after
Axin is recruited to the membrane some change occurs that either inhibits Axin
self-polymerization or inhibits its stability within condensates. But what is the nature of this change?
Wnt signaling causes a switch in the destruction complex mix, destabilizing it
One potential change involves a switch in binding partners. Dvl, one of the first proteins
implicated in Wnt signaling, inhibits destruction complex function in response to Wnt signaling
(reviewed in Mlodzik, 2016). As noted above, Dvl and Axin both have a DIX domain. This
shared domain mediates both self-polymerization and hetero-polymerization (Fiedler et al.,
2011; Kishida et al., 1999; Schwarz-Romond et al., 2007a; Schwarz-Romond et al., 2007b;
Smalley et al., 1999). Dvl is essential for Wnt receptor phosphorylation, Axin recruitment to the
membrane and signalasome endocytosis, thus turning down destruction complex function (Bilic
et al., 2007; Cliffe et al., 2003). Dvl binding to Fz, via Dvl’s DEP and/or PDZ domains (Axelrod et
al., 1998; Wong et al., 2003), is followed by a conformational change that crosslinks Dsh
polymers, increasing local concentration of Dvl at the receptor as it is endocytosed (Gammons
et al., 2016). DEP-mediated Dvl cross-linking may drive Axin recruitment by increased avidity,
driving Dvl:Axin hetero-polymerization. FRAP analysis revealed that Dvl:Axin co-assembly
enhances Axin turnover in puncta (Schwarz-Romond et al., 2007b). Dvl destabilization of Axin
puncta thus provides one mechanism by which Dvl could inactivate the destruction complex. In
22
Figure 1.4
Figure 1.4: In vivo recruitment of APC2 into Axin:GFP puncta. A) Model illustrating Wnt signaling in a Drosophila embryo. One row of cells per body segment produce and secrete the
Wnt Wingless (Wg). It forms a graded distribution and stabilizes the fly βCat (Arm), leading to
graded Arm levels across the body segment. B) Stage 9 Drosophila embryos expressing
23
(Pronobis et al., 2015). Intriguingly, co-expressing Axin, APC, and Dvl2 in cultured cells
revealed that APC:Axin:Dvl2 complexes are rare while Axin:APC or Axin:Dvl2 complexes are
more frequent, consistent with a competition between APC and Dvl for interaction with Axin
(Mendoza-Topaz et al., 2011). Competition for Axin binding is also consistent with the fact that
protein levels of APC:Axin:Dsh in Drosophila embryos are all in the same order of magnitude
(Schaefer et al., 2018). The possibility that DIX:DIX interactions between Dvl and Axin inhibit
Axin is also consistent with fact that DrosophilaAxin∆DIX is constitutively active in βCat
destruction (Peterson-Nedry et al., 2008). One remaining question is how Axin decides between
binding its different partners? Condensates form via multiple multivalent interactions. Perhaps
changes in Axin ADP-ribosylation or phosphorylation in response to Wnt signaling, altering the
charge of the intrinsically disordered region, reduce interaction between Axin:APC or promote
Axin:Dvl interaction. Future research into the rules regulating the competition between
assembly/disassembly of the destruction complex and that of the signalasome will provide
essential insights.
One consequence of supermolecular assembly: the kinase GSK3 plays both positive and
negative roles in the destruction complex via its access to many targets
GSK3, first discovered as a kinase regulating glycogen metabolism, plays pleiotropic roles in
the cell, regulating multiple signaling pathways (Cormier and Woodgett, 2017). GSK3 was one
of the first proteins with a known biochemical role to be placed in the Wnt pathway and the first
negative regulator, a role defined via genetic analysis in Drosophila (Peifer et al., 1994;
Siegfried et al., 1992; Siegfried et al., 1990; Siegfried et al., 1994). This raises the question of
how is pathway specificity maintained? Recruitment into different supermolecular complexes
provides a mechanism. Both CK1 and GSK3 are recruited by Axin into the destruction complex,
where they sequentially phosphorylate βCat, priming it for destruction. However, subsequent
24
it is not surprising that recruiting an active kinase into a multiprotein complex allows it to
phosphorylate many proteins within. Within the active destruction complex, GSK3
phosphorylates Axin to keep Axin “open” for βCat interaction (Kim et al., 2013), phosphorylates
APCs 20Rs to increase affinity for βCat (Ha et al., 2004; Liu et al., 2006; Xing et al., 2004), and
phosphorylates R2/B to facilitate βCat release to the E3 ligase (Pronobis et al., 2015). However,
GSK3 is also recruited into the Wnt signalsome, where it plays important roles. In response to
Wnt signaling, CK1 and GSK3 phosphorylate the tail of LRP5/6, creating a binding site that
facilitates Axin recruitment to the receptor complex for inactivation (Tamai et al., 2004; Zeng et
al., 2005), a process that is visualized in Drosophila as GSK3-dependent recruitment of Axin
puncta to the membrane (Cliffe et al., 2003; Schaefer et al., 2018). Intriguingly, the LRP5/6
phosphorylated tail then can act as a GSK3 inhibitor (Piao et al., 2008; Stamos et al., 2014; Wu
et al., 2009), providing another mechanism by which Wnt activation turns down the destruction
complex. GSK3 phosphorylation of Axin also allows it to be “open” for binding LRP5/6 (Kim et
al., 2013). In these two roles GSK3 is a positive effector of Wnt signaling.Vertebrates have two
GSK isoforms, and questions remain about the roles they play (or do not) in regulating this
pathway. Genetic analysis demonstrated that the two isoforms are largely redundant for Wnt
regulation, with single mutants having tissue-specific defects (Doble et al., 2007; Hoeflich et al.,
2000), though recent studies with isoform specific inhibitors suggest possible differences in
isoform function in Wnt regulation (Chen et al., 2017).
Axin post-translational regulation plays complex roles
Axin features in most studies of destruction complex downregulation, with models ranging
from enhanced Axin degradation, to its dissociation from GSK3, or changes in its assembly
state (reviewed in MacDonald and He, 2012; Nusse and Clevers, 2017). These diverse data
may reflect differences in different animals and/or tissues or, as we think more likely, may reflect
25
Many models of destruction complex inhibition involve Axin regulation via post-translational
modifications, including phosphorylation/dephosphorylation, ADP-ribosylation, or ubiquitination.
As noted above, Axin phosphorylation plays important but diverse roles. Interestingly a recent
report suggested that all of these Axin phosphorylation events are dependent on APC
(Tacchelly-Benites et al., 2018).
One common consequence of long-term Wnt signaling is down-regulation of Axin protein
levels (Kofron et al., 2007; Liu et al., 2005; Mao et al., 2001; Tolwinski et al., 2003; Yang et al.,
2016). GSK3 phosphorylation of Axin can stabilize the protein, while Axin de-phosphorylation by
PP2A leads to degradation (Willert et al., 1999; Yamamoto et al., 1999). Recent work highlights
the role of kinases as “dissolvases” of other biomolecular condensates (Rai et al., 2018). Two
other post-translational modifications of Axin, ADP-ribosylation and ubiquitination, are also
proposed to regulate Axin stability and thus Wnt signaling. Tankyrase is a poly(ADP-ribose)
polymerase, adding ADP-ribose moieties to target proteins, which are then often ubiquitinated
and destroyed (Hsiao and Smith, 2008; Mariotti et al., 2017). Tankyrase binds Axin and
ADP-ribosylates it (Huang et al., 2009), targeting it for ubiquitination by RNF146, a
poly(ADP-ribose)-directed E3 ligase (Callow et al., 2011; Zhang et al., 2011; Zhou et al., 2011). Wnt signaling
induces Axin ADP-ribosylation within 30 min (Yang et al., 2016). Thus it seemed plausible that
Tankyrase caused the Axin degradation seen after longer exposure to Wnt signaling. Consistent
with this, Tankyrase inhibition stabilizes Axin puncta formation in both mammalian and
Drosophila cells (de la Roche et al., 2014; Thorvaldsen et al., 2015; Waaler et al., 2011).
However, things are not that simple. ADP-ribosylation of Axin also enhances Axin’s ability to
immunoprecipitate with active LRP5/6 (Yang et al., 2016). Further, Axin is still degraded after
long Wnt signaling exposure in fly Tankyrase mutants, or when Axin lacks its Tankyrase binding
domain (AxinΔTBD) (Wang et al., 2016b). This suggests Tankyrase is not solely responsible for
26
shortly after Wnt exposure is now seen in all cells, not just those receiving Wg, suggesting other
potential effects of ADP-ribosylation (Yang et al., 2016). Intriguingly, Tankyrase can also bind
APC (Croy et al., 2016). It is important to keep in mind that Drosophila lacking Tankyrase are
viable and fertile (Feng et al., 2014), and mice lacking both Tankyrase proteins, while embryonic
lethal, survive to E10 without obvious defects in Wnt signaling. Thus while Tankyrase can finely
regulate destruction complex activity, as it does in the Drosophila intestine (Wang et al., 2016c),
it is not an essential regulator of the pathway.
If Tankyrase is not essential for Axin degradation, then how are Axin levels decreased after
Wnt signaling? This may be triggered by the RING domain E3 ligase Seven in absentia (Sina)
homolog SIAH 1/2. SIAH’s potential role in Wnt signaling was first identified in a yeast 2-hybrid
screen for novel APC binding partners. Further exploration of this interaction in human cells and
Xenopusembryos indicated that SIAH can ubiquitinate βCat, labeling it for proteasomal
degradation, independent of βTrCP (Liu et al., 2001; Matsuzawa and Reed, 2001). However,
recently SIAH 1 and SIAH 2 were identified as novel regulators of Axin levels in HEK293T cells.
Co-crystallization revealed that SIAH directly binds Axin near its GSK3 binding site.
Interestingly, while SIAH and GSK3 can simultaneously interact with Axin, there is competition
between the two, since the binding of one can allosterically inhibit binding of the other (Ji et al.,
2017). Based on these data, one could hypothesize that after Wnt signaling inhibits GSK3
interaction with Axin, SIAH binds Axin and labels it for proteasomal degradation. Once again,
however, it is important to note that the single Drosophila Sina family member, is adult viable
without obvious defects in Wnt signaling (Carthew and Rubin, 1990).
While long-term Wnt exposure decreases Axin levels, recent studies in vivo in Drosophila
suggest that Axin levels initially increase after Wnt signaling is activated (Wang et al., 2016a;
Yang et al., 2016). Axin levels in the cytoplasm initially increase in cells receiving Wnt signals,
27
over the same time frame, but it is hard to rule out that this is not simply an effect of activating
zygotic Axin expression in all cells. This degree of Axin elevation is at the threshold at which
Axin can inhibit Wnt signaling (3-9x; Peterson-Nedry et al., 2008; Schaefer et al., 2018; Wang et
al., 2016b). Why should Wnt signaling elevate Axin levels when this should enhance βCat
destruction and thus inhibit Wnt signaling? One possibility is that the increased pool of
cytoplasmic Axin is largely “inactivated” Axin molecules that cannot form stable puncta. In fact,
membrane-localized Axin puncta in Wnt-ON cells harbor only half the number of Axin molecules
as cytoplasmic puncta in Wnt-OFF cells, while cytoplasmic Axin levels rise (Schaefer et al.,
2018). Together, these data reveal many levels at which Axin is regulated by post-translational
modification and open up questions for future research, defining which changes are the initial
response to Wnt signals and which are adjustments allowing longer term modulation of
signaling.
APC mutations in colorectal cancer target specific aspects of destruction complex function
Activating mutations in the Wnt pathway play roles in many cancers, including endometrial
and liver cancer, but are most prominent in colorectal tumors, where they initiate oncogenesis
(reviewed in Zhang and Shay, 2017). ~10% of colorectal tumors have gain-of-function βCat
mutations disrupting phosphorylation and thus destruction, a few have loss-of-function Axin
mutations, but >80% are APC mutant (Kandoth et al., 2013). These mutations have a very
striking feature—unlike most tumor suppressors where selection favors homozygous null
mutations, all or virtually all colorectal tumors have at least one APC allele expressing a
truncated protein. Intriguingly, the truncations occur in a small region of the protein, the mutation
cluster region (MCR; Kohler et al., 2008), leading researchers to explore what properties are
lost or retained to favor selection. Most now accept the “just right” hypothesis (Albuquerque et
al., 2002), which proposes that complete loss of APC function leads to such high levels of Wnt
28
protein retains some function. What functions are retained and which lost? One critical thing lost
in the truncated proteins are the SAMP motifs, the high affinity Axin binding sites. A mouse
mutant with one allele truncated to lack all SAMPs is tumor prone while mice carrying an allele
with a slightly longer truncation retaining one SAMP are not tumor prone (and in fact are
homozygous viable!; Smits et al., 1999). However, closer examination of the MCR suggested
more is going on, focusing attention on the B/CID motif, which is just N-terminal to the last
SAMP (Fig. 3A) and thus also disrupted in most or all tumor truncations (Kohler et al., 2008).
Thus selection may favor loss of both the SAMPS and B/CID. What function is retained by
truncated APC to prevent selection for null mutations? Truncated APC proteins like those in
tumors cannot promote βCat destruction but can still bind βCat (Roberts et al., 2011) and
mediate its phosphorylation (Yang et al., 2006). Retaining βCat in the cytoplasm may thus be
how destruction complexes containing truncated APC dampen but do not eliminate Wnt
signaling. It will be intriguing to further explore assembly and function of destruction complex
condensates carrying truncated APC, as deleting the SAMP motifs reduces but does not
eliminate APC incorporation into puncta (Pronobis et al., 2015; Roberts et al., 2011).
The destruction complex is a multifunctional machine with other targets including the cytoskeleton
While best known for roles in Wnt regulation, destruction complex proteins also have other
functions. The first evidence emerged even before the connection to Wnt signaling was made,
when scientists found that human APC co-localizes with and binds microtubules and the
microtubule plus end protein EB1(Munemitsu et al., 1994; Smith et al., 1994; Su et al., 1995).
Subsequent work on APC led to suggested roles in spindle orientation, chromosome
segregation, and polarity of neurons and migrating cells, but many of these studies rely on
overexpressing full-length or truncated APC (reviewed in Nelson and Nathke, 2013; Rusan and
Peifer, 2008). Use of Drosophila allowed analysis of complete loss of APC function, revealing
29
Diaphanous (McCartney et al., 2001; Poulton et al., 2013; Webb et al., 2009), mitotic spindle
orientation (Yamashita et al., 2003) and microtubule dynamics in neuronal dendrites (Mattie et
al., 2010; Weiner et al., 2016), but casting doubt on suggested roles in axon or dendrite polarity
(Rusan et al., 2008). C. elegans APR-1 helps orient mitotic spindles by attenuating cortical
spindle pulling forces (Sugioka et al., 2018; Sugioka et al., 2011). At least some APC proteins
also regulate actin dynamics, in part through a “rocket launching” mechanism in which they work
with Diaphanous to stimulate filament nucleation and extension (Breitsprecher et al., 2012;
Jaiswal et al., 2013), and thus regulate focal adhesion turnover (Juanes et al., 2017). When
considering cytoskeletal roles for APC, however, it is critical to rule out places where effects on
Wnt signaling lead to downstream cytoskeletal alterations (e.g., Elbaz et al., 2016; Eom et al.,
2014; Hayden et al., 2007; Nakagawa et al., 2017; Yokota et al., 2009).
Other destruction complex or signalasome proteins also may have cytoskeletal roles. For
example, Axin has suggested roles in mouse oocyte meiosis (He et al., 2016) and in axon and
dendrite morphogenesis and intermediate neuronal progenitor differentiation in the cerebral
cortex (Chen et al., 2015; Fang et al., 2013; Fang et al., 2011). Once again, however, one must
be cautious in differentiating Wnt-independent from Wnt dependent roles. The Wnt regulator Dvl
and receptor Fz have well known roles in planar cell polarity and in cilia genesis/orientation, but
these appear to be largely independent of the destruction complex and canonical βCat signaling
(reviewed in Adler and Wallingford, 2017). There is one common cytoskeletal thread among
different destruction complex proteins and regulators—APC, Axin, Dvl and βCat are all reported
to localize to centrosomes in at least some cell types (reviewed in Bryja et al., 2017; Mbom et
al., 2013). While some data suggests roles for centrosomes and cilia in canonical Wnt signaling,
both flies and mice lacking centrosomes develop without strong defects in Wnt signaling (Basto
30
Figure 1.5
Figure 1.5: A revised model of the destruction complex. Polymers of Axin and of APC, mediated by polymerization of their respective DIX and ASAD/Arm repeat domains, intertwine. Polymers interact via the RGS:SAMP and Arm repeat-Axin motif interactions. Polymers