In vitro evaluation of the effects of Lavandula officinalis and Origanum vulgare essential oils on ruminal fermentation using concentrate and roughage type substrates
Full text
Figure
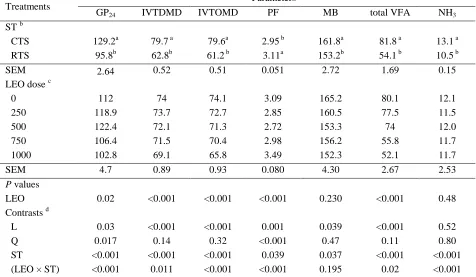
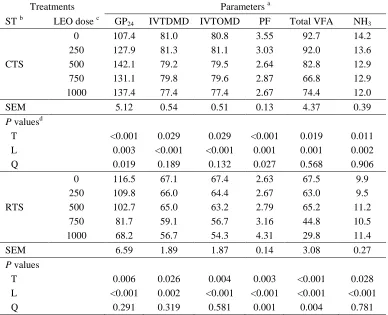
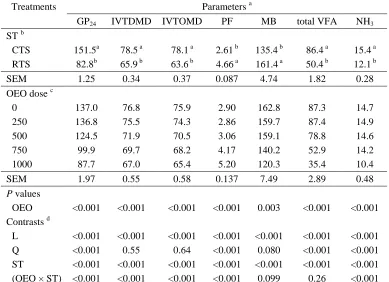
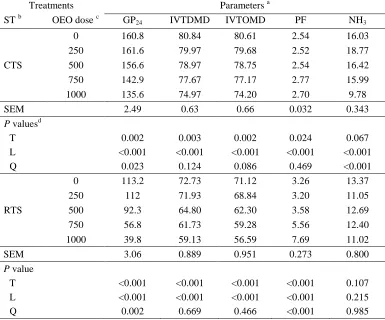
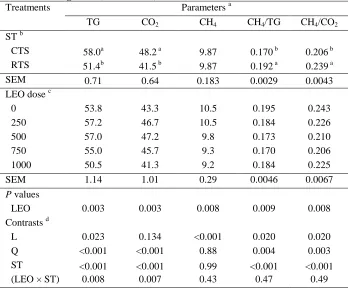
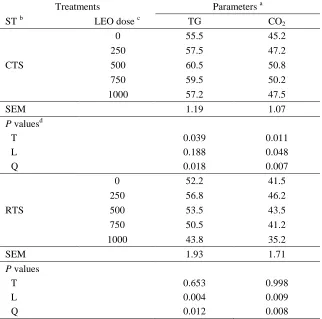
Related documents
Using disc diffusion method to show the antibacterial effect of ethanol extract of Spirulina platensis against different bacterial strains ( Escherichia coli ,
When 17 women in Australia were interviewed about their experiences of health care after being diagnosed with breast cancer during pregnancy, they reported particularly disliking
Linear stapler with color indicator 30mm, 3.5mm staple height Linear stapler with color indicator 30mm, 4.8mm staple height Linear stapler with color indicator 45mm, 3.5mm staple
As regard sunflower-mungbean intercropping system all inter- crops gave more economic returns than sole cropping of sunflower and mungbean and planting 6 sunflower plants + 30
Our study follows a factorial design to assess the benefit of each data stream, but also to evaluate the potential of as- similating more than one data stream and its effect on
To prepare for using video conferencing as a learning delivery mode for farmer groups, Margaret Brown, the leader of the FeedSmart project, spent time at North Dakota State
The tasks were organized by discipline (propulsion, structures, aeromechanics, acoustics, and handling qualities) although all elements are connected because of the
This program offers employers access to a pool of applicants with disabilities, pertinent disability employment information, a network of companies sharing information on
