R E S E A R C H
Open Access
Aberrant methylation of
Pax3
gene and
neural tube defects in association with
exposure to polycyclic aromatic
hydrocarbons
Shanshan Lin
1,2, Aiguo Ren
1*, Linlin Wang
1*, Chloe Santos
3, Yun Huang
1, Lei Jin
1, Zhiwen Li
1and Nicholas D. E. Greene
3Abstract
Background:Neural tube defects (NTDs) are common and severe congenital malformations.Pax3is an essential
gene for neural tube closure in mice but it is unknown whether altered expression or methylation ofPAX3 contributes to human NTDs. We examined the potential role of hypermethylation ofPax3in the development of NTDs by analyzing human NTD cases and a mouse model in which NTDs were induced by benzo[a]pyrene (BaP), a widely studied polycyclic aromatic hydrocarbon (PAH).
Methods:We extracted methylation information ofPAX3in neural tissues from array data of ten NTD cases and eight non-malformed controls. A validation study was then performed in a larger independent population comprising 73 NTD cases and 29 controls. Finally, we examined methylation patterns and expression ofPax3in neural tissues from mouse embryos of dams exposed to BaP or BaP and vitamin E.
Results:Seven CpG sites inPAX3were hypermethylated in NTD fetuses as compared to controls in the array data. In the validation phase, significantly higher methylation levels in the body region ofPAX3were observed in NTD cases than in controls (P= 0.003). And mean methylation intensity in the body region ofPAX3in fetal neural tissues was positively correlated with median concentrations of PAH in maternal serum. In the mouse model, BaP-induced NTDs were associated with hypermethylation of specific CpG sites within both the promoter and body region of Pax3. Supplementation with vitamin E via chow decreased the rate of NTDs, partly recovered the repressed total antioxidant capacity in mouse embryos exposed to BaP, and this was accompanied by the normalization ofPax3 methylation level and gene expression.
Conclusion:Hypermethylation ofPax3may play a role in the development of NTDs; DNA methylation aberration
may be caused by exposure to BaP, with possible involvement of oxidative stress.
Keywords:Neural tube defects,Pax3gene, Methylation, Benzo[a]pyrene, Polycyclic aromatic hydrocarbon
* Correspondence:[email protected];[email protected] 1
Institute of Reproductive and Child Health, National Health Commission Key Laboratory of Reproductive Health, and Department of Epidemiology and Biostatistics, School of Public Health, Peking University Health Centre, Peking University, Beijing 100191, China
Full list of author information is available at the end of the article
Background
Neural tube defects (NTDs) arise from a failed or disor-dered closure of the neural tube during embryogenesis. The occurrence of NTDs is around 0.5–2/1000 pregnan-cies worldwide [1]. Fetuses affected with an NTD are often stillborn (e.g., anencepahlics), and most surviving infants suffer from life-long disabilities. The etiology of
NTDs is complex, involving both genetic and
non-genetic factors [2]. Over 300 genes have been iden-tified to be involved in the regulation of neural tube closure in mouse NTD mutants [3, 4]; however, only a few of these genes have successfully been validated in human NTDs. In addition to potential additive effects of multiple risk alleles, increasing attention has also fo-cused on the potential for epigenetic alterations to con-tribute to the occurrence of NTDs by mediating the interplay of fetal genetics and environmental factors [5].
Epigenetic modification can cause changes in gene ex-pression that are not directly related to the DNA se-quence itself, of which DNA methylation is one of the best understood epigenetic mechanisms [6]. During early development, a tight regulation of genome-wide erasure of epigenetic footprints with resetting of the methylation signature takes place, making developing fetuses particu-larly susceptible to epigenetic dysregulation as a conse-quence of environmental exposure [7, 8]. Abnormal genome-wide methylation during embryogenesis has been linked to developmental abnormalities at birth, in-cluding NTDs. Recent studies have demonstrated that global DNA hypomethylation, evaluated using LINE-1 methylation as an indicator in human fetuses, was asso-ciated with an increased risk for NTDs [9]. Aberrations of methylation at specific genes are reported to be in-volved in NTDs, including imprinted genes [10, 11], DNA repair genes [12], planar-cell polarity genes [13,
14], andHOXgenes [15].
Pax3 is a paired-homeodomain-containing transcrip-tion factor essential for promoting neural crest induc-tion, maintenance, migrainduc-tion, and differentiation [16]. Previous studies in mice have shown that Pax3 function is required for neural tube closure. Several alleles of
Pax3 cause NTDs in mice and homozygous Splotch
(Sp2H) embryos develop NTDs with 100% penetrance [17]. However, the role of PAX3 in human NTDs re-mains unclear. Recessive and dominant mutations in
PAX3in humans are known to cause Waardenburg syn-drome, an autosomal dominant condition that affects neural crest-derived structures and also includes spina bifida as part of its phenotypic spectrum [18]. A 5-bp deletion in exon 5 of the PAX3gene was reported in a patient with spina bifida [19]. In contrast, the results from a case-control study including 74 infants with spina bifida and 87 non-malformed controls indicated that variants in PAX3 were not strong risk factors for
human spina bifida [20]. Therefore, exploring mecha-nisms other than coding sequence variants, such as methylation modification, in PAX3 may provide novel insight into the etiology of human NTDs.
We hypothesized that aberrant DNA methylation of
Pax3 plays a role in the formation of NTDs. To test this hypothesis, we first compared the difference in methyla-tion levels of CpG sites within PAX3 using genomic methylation array data with DNA from neural tissues of NTD cases and non-malformed controls. The methylation status of the CpG sites that were found to differ was then validated in a larger NTD case-control population. We tested whether there was any correlation between CpG site methylation levels in fetal neural tissues and maternal serum concentrations of polycyclic aromatic hydrocarbons (PAHs), a class of ubiquitous environmental pollutants that have been shown to be associated with the risk for NTDs in epidemiological studies [21]. Finally, we assessed the methylation level and gene expression of Pax3 in neural tissues from mouse embryos exposed in utero to benzo[a]pyrene (BaP), a widely studied PAH that induces NTDs [22]. In addition, we assayed markers of oxidative stress in fetal mice to further explore the possible mecha-nisms by which BaP might affect methylation regulation.
Results
Methylation ofPAX3gene in genomic microarray
A detailed description on the genome-wide methylation re-sults obtained from Infinium HumanMethylation450 Bead-Chip (HM450K), using DNA isolated from neural tissues from ten NTD cases and eight unrelated non-malformed controls can be found elsewhere [23]. In brief, out of 485,199 CpG sites across the entire genome, 23,294 (4.8%) were differentially methylated between cases and controls. Of the differentially methylated CpG sites, 12,383 (53.2%) were significantly hypermethylated and 10,911 (46.8%) were significantly hypomethylated in NTD cases when compared to controls. The characteristics of the NTD cases and con-trols in phase 1 are presented in Additional file1: Table S1.
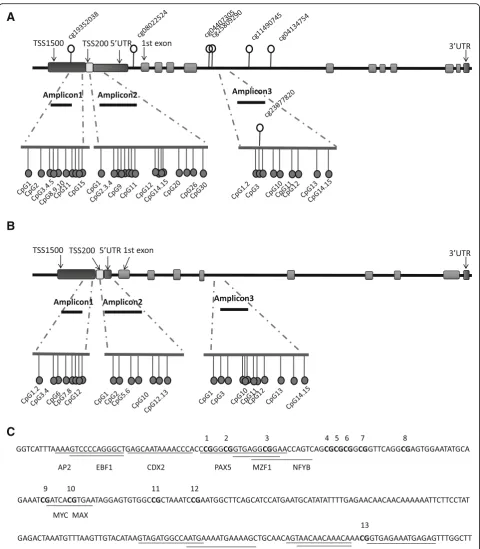
In thePAX3gene, the focus of this study, a total of 54 CpG sites were extracted from the HM450K array data, of which 47 CpG sites (87.0%) were found to be hyper-methylated. And 7 out of the 47 CpG sites exhibited sta-tistically greater methylation in NTD fetuses than in controls (Additional file 2: Table S2). Analysis of the genomic location of the seven significantly hypermethy-lated CpG sites showed that one CpG was located at TSS1500, one in the 5′UTR, and the remaining five CpGs were within the body of the gene (Fig.1a).
Validation ofPAX3methylation in an independent cohort of NTD cases and controls
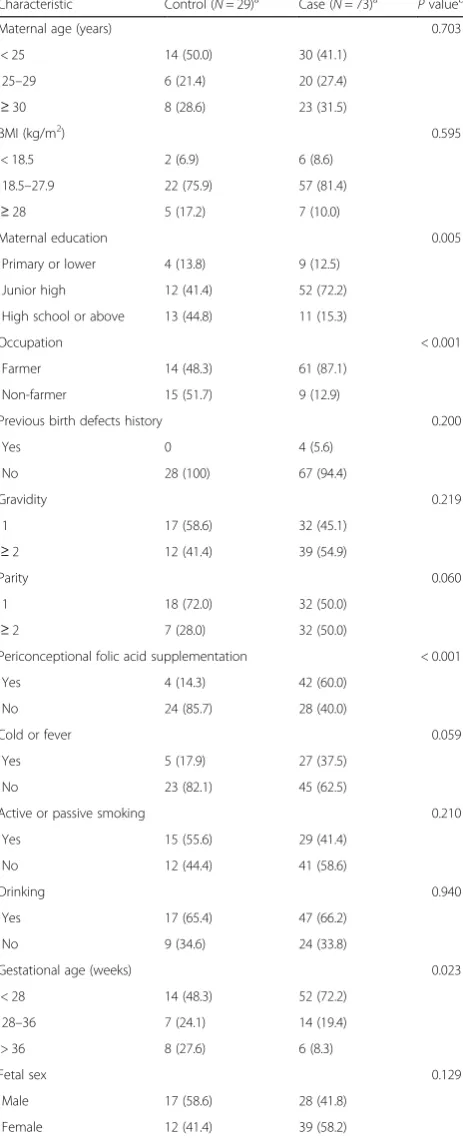
In order to validate these findings, 73 NTD cases and 29
A
B
C
methylated regions of PAX3 identified in phase 1 using Sequenom EpiTYPER. The demographic characteristics of the subjects are described in Table 1. Three DNA amplicons were developed that cover 35 CpGs (Fig. 1a). In the present study, TSS1500, TSS200, 5′UTR, and the 1st exon were defined as the promoter region of PAX3
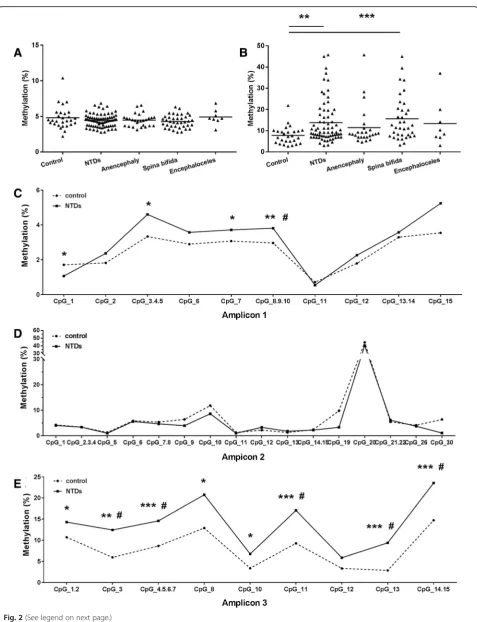
gene. As shown in Fig. 2a, no significant difference in overall mean methylation levels in the promoter region ofPAX3was observed between NTD cases and controls. However, NTD samples exhibited significantly higher DNA methylation levels (13.9 ± 10.1%) than control sam-ples (7.8 ± 4.2%) in the gene body region (P = 0.003) (Fig. 2b). Furthermore, within the gene body, all CpG sites except CpG_12 showed significantly higher levels of methylation in the NTD samples compared to con-trols (Fig. 2e). After Benjanmini-Hochberg correction, methylation levels in five of the nine CpG sites were still significantly higher in NTD cases.
The relationship between hypermethylation of PAX3
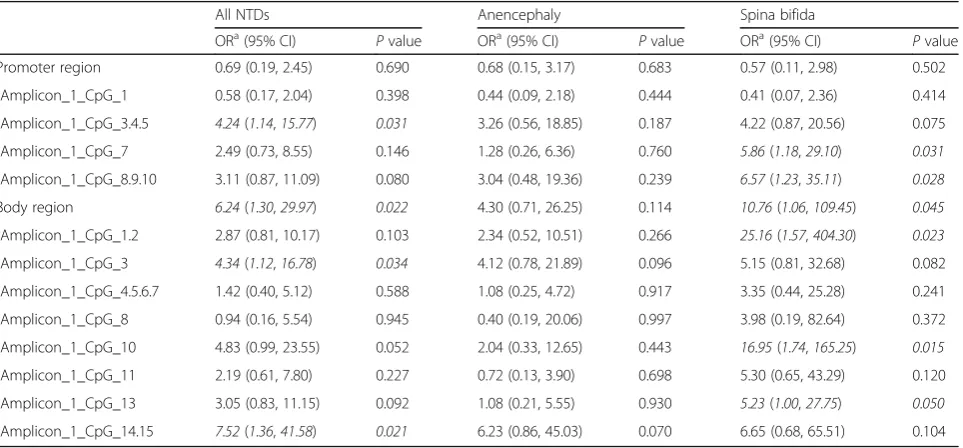
and the risk of NTDs was examined. Methylation levels were categorized according to the median methylation level of the controls. As shown in Table2, a higher level of methylation in the gene body region was associated with 6.24-fold increased risk for NTDs (95% CI 1.30– 29.97).
Correlation between PAH concentrations in maternal serum andPAX3methylation in fetal neural tissues
A previous study from our team found that higher PAH concentrations in maternal serum were associated with an increased risk for NTDs [24]. We therefore con-ducted a correlation analysis between PAH concentra-tions in maternal serum and PAX3 methylation levels in fetal neural tissues (N= 51 mother-fetus pairs) to further our understanding of the potential relationship of PAH exposure, gene methylation, and NTDs risk. Notably, a significant positive correlation was found be-tween mean methylation levels in thePAX3 gene body and median concentrations of high-molecular-weight
PAHs in maternal serum (r= 0.310. P= 0.027)
(Additional file3: Table S3).
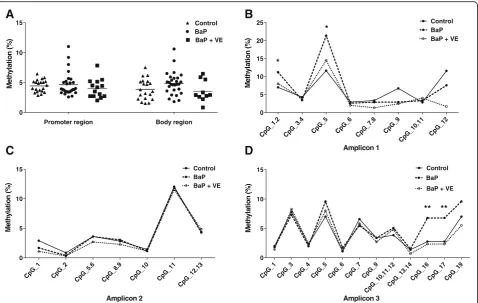
Disturbed methylation ofPax3in mouse embryos exposed to BaP
We further investigated the finding of a correlation
be-tween maternal serum PAH concentrations and PAX3
hypermethylation in humans by utilizing a BaP-induced NTD mouse model. Three amplicons developed accord-ing to the mouse genome were used (Fig. 1b). The body region of Pax3 showed a trend toward higher levels of methylation in the BaP-treated group compared with controls (Fig. 3a), but this did not reach statistical sig-nificance. Furthermore, in terms of specific CpG sites, two significantly hypermethylated CpG sites within the promoter and two within the body region of Pax3were respectively detected in the BaP-exposed mouse fetuses when compared to the non-exposed fetuses (Fig.3b–d).
The role of oxidative stress in BaP-related disturbedPax3 methylation and expression
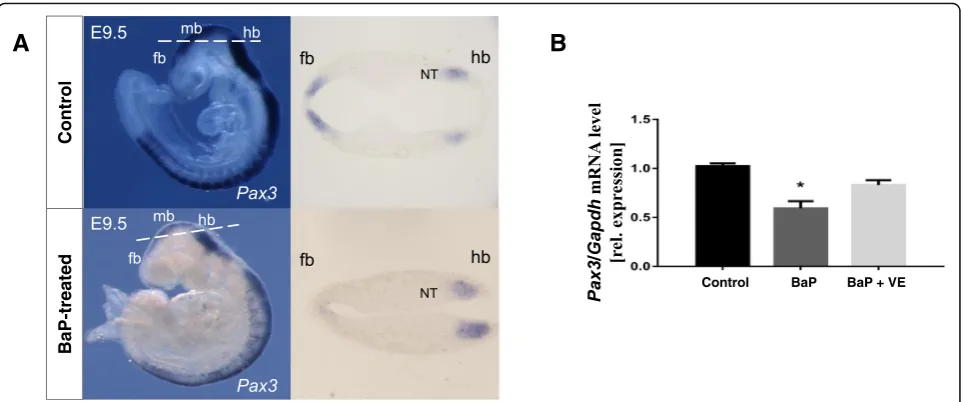
Since BaP exposure was hypothesized to favor the gener-ation of reactive oxygen species (ROS) in the embryos, we inferred that oxidative stress may get involved in the aberrantPax3methylation induced by BaP. As shown in Fig.4, compared to the control group, the levels of total antioxidant capacity (TAC) and the activity of super-oxide dismutases (SODs) were decreased. Real-time PCR and whole mount in situ hybridization showed that BaP exposure significantly decreased the transcription of
Pax3 (P= 0.008) (Fig. 5). After co-supplemented with vitamin E, a commonly used antioxidant, the repressed levels of TAC and SODs were partly restored (Fig. 4). Coincidentally, the hypermethylation of Pax3in specific CpG sites induced by BaP treatment was recovered and
Pax3expression was also normalized by vitamin E sup-plementation (Figs. 3 and 5), which suggested a causal effect of oxidative stress on both Pax3 methylation and expression.
Discussion
Pax3 is a key gene, encoding a transcription factor re-quired for neural tube closure [17]. In this study, we evaluated the methylation status ofPAX3in a two-phase (See figure on previous page.)
design study to examine whether there is a potential role
of PAX3 methylation in the development of human
NTDs. Differentially hypermethylated CpG sites were found in the promoter and gene body region within
PAX3 in the neural tissues of NTD cases. Association analysis showed that a higher methylation level in the gene body region was associated with an elevated risk for NTDs. Moreover, PAH concentrations in maternal serum, which were known to be associated with in-creased risk of NTDs, were positively correlated to
methylation levels at several CpG sites. In the
BaP-induced NTD mouse model, hypermethylation of
Pax3 gene and suppressed gene expression were ob-served in embryos with BaP treatment, along with re-duced TAC level. We previously showed that vitamin E supplementation could decrease the rate of NTDs and alleviate oxidative stress in BaP exposed embryos [22]. Here, we further demonstrated that vitamin E mitigated the shifts inPax3methylation and gene expression.
Studies on the role of the Pax3in NTDs have mostly focused on gene depletion or loss of function mutants [17]. In mice, mutations inPax3give rise to theSplotch
(Sp) phenotype, which includes exencephaly, spina bifida, and neural crest abnormalities in homozygous mutant embryos [25, 26]. The human PAX3 gene
ex-hibits high homology with mouse Pax3. Mutations
within the humanPAX3gene have been found in Waar-denburg syndromes, a condition which is occasionally associated with NTDs [18]. A previous study identified two spina bifida patients who had small interstitial chromosomal deletions involving PAX3 [27]. Exon se-quencing of PAX3 in 114 cases with spina bifida also identified two common variants; however, without un-affected individuals, the influence of these variants on the risk of spina bifida could not be determined [28]. To the contrary, the screening results from 74 spina bifida cases and 87 control infants suggested that variant in
PAX3was not a major contributor to the overall burden of NTDs at population level [20]. Recently, evidence from animal studies suggested that aberrant methylation of Pax3 was involved in the development of NTDs in-duced by hyperglycemia [29, 30]. However, to date, no study investigating the role of PAX3 methylation in the etiology of human NTDs has been reported. In our study, we analyzed the methylation status of PAX3gene in neural tissues from human fetuses, and our results showed that the methylation level in PAX3 was signifi-cantly higher in NTD cases than in non-malformed controls.
Unexpectedly, in our study, compared with promoter region, the body region of PAX3 was more consistently hypermethylated in NTD cases and the differentially methylated region was mainly located in intron 4. Our results imply that a methylation change in the body Table 1Characteristics of NTD cases and controls in phase 2 for
methylation assay
Characteristic Control (N= 29)a Case (N= 73)a Pvalueb
Maternal age (years) 0.703
< 25 14 (50.0) 30 (41.1)
25–29 6 (21.4) 20 (27.4)
≥30 8 (28.6) 23 (31.5)
BMI (kg/m2) 0.595
< 18.5 2 (6.9) 6 (8.6)
18.5–27.9 22 (75.9) 57 (81.4)
≥28 5 (17.2) 7 (10.0)
Maternal education 0.005
Primary or lower 4 (13.8) 9 (12.5)
Junior high 12 (41.4) 52 (72.2)
High school or above 13 (44.8) 11 (15.3)
Occupation < 0.001
Farmer 14 (48.3) 61 (87.1)
Non-farmer 15 (51.7) 9 (12.9)
Previous birth defects history 0.200
Yes 0 4 (5.6)
No 28 (100) 67 (94.4)
Gravidity 0.219
1 17 (58.6) 32 (45.1)
≥2 12 (41.4) 39 (54.9)
Parity 0.060
1 18 (72.0) 32 (50.0)
≥2 7 (28.0) 32 (50.0)
Periconceptional folic acid supplementation < 0.001
Yes 4 (14.3) 42 (60.0)
No 24 (85.7) 28 (40.0)
Cold or fever 0.059
Yes 5 (17.9) 27 (37.5)
No 23 (82.1) 45 (62.5)
Active or passive smoking 0.210
Yes 15 (55.6) 29 (41.4)
No 12 (44.4) 41 (58.6)
Drinking 0.940
Yes 17 (65.4) 47 (66.2)
No 9 (34.6) 24 (33.8)
Gestational age (weeks) 0.023
< 28 14 (48.3) 52 (72.2)
28–36 7 (24.1) 14 (19.4)
> 36 8 (27.6) 6 (8.3)
Fetal sex 0.129
Male 17 (58.6) 28 (41.8)
Female 12 (41.4) 39 (58.2)
aData were presented in number (percentage). Total number may not be
equal to the total of cases or controls due to missing or unknown data
bCases and controls were compared by Pearson’sχ2test, or Fisher’s exact test
region ofPAX3may be an epigenetic component of hu-man NTDs, which is reminiscent with findings for the
HOXB7 gene in a myelomeningocele (spina bifida) case-control study [15]. In the latter study, a genome-wide methylation assay found three CpGs in
HOXB7 gene body to be hypomethylated in myelome-ningocele patients when compared to controls, and these were further verified in a larger population, using the Sequenom EpiTYPER platform. Studies on
GRHL3 and SOX18 also reported that altered methy-lation within the gene body was associated with the risk for NTDs [14, 31].
It is generally accepted that DNA methylation changes in the promoter and gene body regions may have differ-ential effects on gene expression. DNA methylation in promoter regions is usually negatively associated with gene expression, whereas in the gene body, the impact is not consistent, with both positive and negative impacts having been reported [32, 33]. In the present study, a negative correlation between gene body DNA methyla-tion and transcripmethyla-tion level of Pax3 was observed in mouse embryos, which was in line with the finding in hyperglycemia-induced NTDs [29]. The underlying mechanisms of gene body methylation in regulating gene
expression have not been well understood. Recently, in-creasing evidence demonstrating the role of DNA methylation in alternative splicing regulation has been reported, which is essential for providing tissue-specific features for some genes [34]. DNA methylation is nor-mally more abundant in exons compared to the flanking introns, which is a marker for distinguishing exons from introns. Previous studies proposed that the change of exon methylation levels would affect the recognition of exons while splicing [35]. It is thus reasonable to hypothesize that the methylation status of intron might also be crucial for alternative splicing. Further,cis-acting element has been recognized in intron, which could regulate gene expression [34]. Within intron 4 ofPAX3, a number of transcription factors were predicted to bind at several CpG sites examined in the present study, with high predictive values (predictive value > 8, JASPAR, Fig.1c), which might be responsible for the observed re-pressed gene expression. However, more research is ne-cessary to examine this point in detail.
PAX3 hypermethylation by itself is not likely to be a sole cause for NTDs but rather be part of a complex combination of environmental and epigenetic risk
fac-tors. Our previous epidemiological studies have
(See figure on previous page.)
Fig. 2PAX3methylation pattern assayed by Sequenom EpiTYPER in NTD cases and controls.aMean methylation level ofPAX3promoter region (amplicon 1 and amplicon 2) between NTD cases and controls.bMean methylation level ofPAX3body region (amplicon 3) between NTD cases and controls.c–eMethylation levels for each CpG site between NTD cases and controls in promoter (c,d) and body region (e). TSS1500, TSS200, 5′URT, and the 1st exon were defined as the promoter region ofPAX3gene in this study. The significance of differences was calculated using Independent t-test. *P< 0.05, **P< 0.01, ***P< 0.001, compared with control group.#P< 0.05, compared with control group after FDR adjustment
Table 2Risks of NTDs associated with methylation level ofPAX3gene in fetal neural tissues
All NTDs Anencephaly Spina bifida
ORa(95% CI)
Pvalue ORa(95% CI)
Pvalue ORa(95% CI)
Pvalue Promoter region 0.69 (0.19, 2.45) 0.690 0.68 (0.15, 3.17) 0.683 0.57 (0.11, 2.98) 0.502
Amplicon_1_CpG_1 0.58 (0.17, 2.04) 0.398 0.44 (0.09, 2.18) 0.444 0.41 (0.07, 2.36) 0.414
Amplicon_1_CpG_3.4.5 4.24(1.14,15.77) 0.031 3.26 (0.56, 18.85) 0.187 4.22 (0.87, 20.56) 0.075
Amplicon_1_CpG_7 2.49 (0.73, 8.55) 0.146 1.28 (0.26, 6.36) 0.760 5.86(1.18,29.10) 0.031
Amplicon_1_CpG_8.9.10 3.11 (0.87, 11.09) 0.080 3.04 (0.48, 19.36) 0.239 6.57(1.23,35.11) 0.028
Body region 6.24(1.30,29.97) 0.022 4.30 (0.71, 26.25) 0.114 10.76(1.06,109.45) 0.045
Amplicon_1_CpG_1.2 2.87 (0.81, 10.17) 0.103 2.34 (0.52, 10.51) 0.266 25.16(1.57,404.30) 0.023
Amplicon_1_CpG_3 4.34(1.12,16.78) 0.034 4.12 (0.78, 21.89) 0.096 5.15 (0.81, 32.68) 0.082
Amplicon_1_CpG_4.5.6.7 1.42 (0.40, 5.12) 0.588 1.08 (0.25, 4.72) 0.917 3.35 (0.44, 25.28) 0.241
Amplicon_1_CpG_8 0.94 (0.16, 5.54) 0.945 0.40 (0.19, 20.06) 0.997 3.98 (0.19, 82.64) 0.372
Amplicon_1_CpG_10 4.83 (0.99, 23.55) 0.052 2.04 (0.33, 12.65) 0.443 16.95(1.74,165.25) 0.015
Amplicon_1_CpG_11 2.19 (0.61, 7.80) 0.227 0.72 (0.13, 3.90) 0.698 5.30 (0.65, 43.29) 0.120
Amplicon_1_CpG_13 3.05 (0.83, 11.15) 0.092 1.08 (0.21, 5.55) 0.930 5.23(1.00,27.75) 0.050
Amplicon_1_CpG_14.15 7.52(1.36,41.58) 0.021 6.23 (0.86, 45.03) 0.070 6.65 (0.68, 65.51) 0.104
a
suggested that maternal exposure to PAHs was associ-ated with an elevassoci-ated risk of NTDs in the offspring [21,
24]. Cohort studies reported that prenatal PAH exposure was associated with lower global methylation and hyper-methylation ofinterferonγin umbilical cord white blood cells [36,37]. Several toxicological studies have also sug-gested that BaP exposure could disrupt DNA methyla-tion status [38,39]. Importantly, in the present study, we
found that differentially methylated CpG sites in PAX3
gene in fetal neural tissues were positively correlated with PAH concentrations in maternal serum. In line with these findings in human subjects, our mouse experiment showed that the methylation level ofPax3 was elevated after BaP treatment, indicating that the presence of BaP impacts Pax3 methylation regulation. These findings support the hypothesis that hypermethylation of Pax3
Promoter region Body region
0 5 10 15
BaP + VE
M e th yl at io n (% ) Control BaP CpG_ 1.2 CpG_ 3.4 CpG_ 5 CpG_ 6 CpG_ 7.8 CpG_ 9 CpG_ 10.1 1 CpG_ 12 0 5 10 15 20 25 Control BaP BaP + VE
Me th yl at io n (%) * * CpG_ 1 CpG _2 CpG_ 5.6 CpG_ 8.9 CpG_ 10 CpG_ 11 CpG _12.1 3 0 5 10 15 Me th y la ti o n (% ) Control BaP BaP + VE
CpG _1
CpG_3CpG_4CpG _5 CpG _6 CpG_ 7 CpG_ 9 CpG_ 10.1 1.12 CpG _13. 14 CpG_1 6 CpG_ 17 CpG _19 0 5 10 15 Control BaP
BaP + VE
M e th y la tio n (% ) ** **
A
B
C
D
Amplicon 1Amplicon 2 Amplicon 3
Fig. 3The effects of BaP and vitamin E onPax3methylation in E10.5 mouse embryos.aMean methylation level of promoter region (amplicon 1 and amplicon 2) and body region (amplicon 3) withinPax3.b–dMethylation level for specific CpG sites in promoter region (b,c) and body region (d) among control group, BaP-treated group and vitamin E co-supplementation group (n= 24–28). TSS1500, TSS200, 5′URT, and the 1st exon were defined as the promoter region ofPax3gene in this study. The significance of differences was calculated using ANOVA. *P< 0.05, **P
< 0.01, ***P< 0.001, compared with control group.VEvitamin E
Fig. 4The effects of BaP and vitamin E on TAC and SOD activity in E10.5 mouse embryos. Data were expressed as mean ± SD (n= 8–10). *P< 0.05, **P< 0.01, ***P< 0.001, compared with control group;#
P< 0.05,##
P< 0.01,###
gene is involved in abnormal closure of neural tube sec-ondary to PAH exposure.
In considering how BaP exposure could affect methy-lation, accumulating evidence has demonstrated that oxidative stress in response to various environmental in-sults or maternal dietary factors is responsible for
aber-rant DNA methylation [40–43]. In our study,
hypermethylation of Pax3 and repressed gene expres-sion, along with reduced TAC level, were observed in BaP-treated mouse embryos. Co-administration with vitamin E could rescue NTDs induced by BaP, partly normalized the TAC level, and attenuate the hyperme-thylation ofPax3and the repressed gene expression. All these findings support the postulation that the oxidative stress may be causally involved in aberrant DNA methy-lation in the BaP-treated group, which might be the underlying mechanism for the development of NTDs caused by BaP. Consistent with this idea, a previous in-vestigation revealed that epigallocatechin gallate, which is the major polyphenol in green tea and is believed to act as an antioxidant, could block hypermethylation of several neural tube closure essential genes induced by maternal diabetes, including Pax3 [30]. Although it is not precisely known, previous investigations proposed that oxidative stress could influence the synthesis of S-adenosylmethionine [44], the expression of DNA methyltransferase [45], and the activity of the ten eleven translocation enzymes [46], which are all critical for DNA methylation. On the other hand, epigenetic regula-tion may also contribute to impairment of antioxidant
gene expression [47]. In this regard, the ROS production and epigenetic regulation establish an interconnected cycle, which would amplify external factors toward the progression of pathological disorders.
DNA methylation is considered tissue-specific and there-fore appropriate biological samples for methylation study are crucial. One advantage of our study is that neural tis-sues from fetuses were used for assessment of methylation, which makes our methylation study more relevant to the outcome, as compared to those that used blood DNA as the surrogate [15]. The limitation is that we did not investi-gate the expression ofPAX3 gene in our human subjects, as fresh neural tissue samples of terminated NTD cases for RNA assay are extremely difficult to collect. Although our mouse experiment may provide a clue, additional studies are needed to correlate the methylation levels of thePAX3
gene body with PAX3 gene or protein expression values. Another limitation is that more case mothers reported folic acid supplementation than control mothers, which might cause the differences in methylation levels between the two groups. However, when analyses were performed by folic acid supplementation status in the case group, no differ-ences in the methylation level ofPAX3were found between those with or those without folic acid use (Additional file4: Figure S1), suggesting that folic acid has no impact on methylation level in the present study.
Conclusions
In conclusion, we found that hypermethylation of Pax3
and the downregulation of the gene may be important
A
B
Fig. 5The Effects of BaP and vitamin E onPax3gene expression in mouse embryos.aWhole mount in situ hybridization ofPax3in E9.5 embryos. In control embryos,Pax3mRNA was detected in the dorsal neural tube and somites as expected, while less gene expression was seen in the forebrain and midbrain of embryos exposed to BaP. The dashed lines on the whole mount panels indicate the orientation of respective sections.bRelative quantitative real-time PCR ofPax3mRNA from E10.5 embryos exposed to BaP and co-supplemented with vitamin E. Values were normalized againstGapdhand represented as mean ± SE (n= 5–8). *P< 0.05, compared with control group.fbforebrain,mbmidbrain,hb
events in the development of NTDs during embryogen-esis. A relationship exists between higher maternal serum PAH concentrations and PAX3hypermethylation in fetal neural tissues, which is further supported by our mouse experiments. In addition, oxidative stress may be involved in the process of environmental exposure and methylation modification. Our study provides novel evi-dence on the interaction between genetic and environ-mental factors in the etiology of NTDs.
Methods Study subjects
The human subjects were recruited from rural counties in Shanxi province of northern China (Pinding, Zezhou, Xiyang, Shouyang, and Taigu), where NTD prevalence is among the highest in the world [48]. As described in more detail in our previous report [48], NTD cases were terminated fetuses affected by an NTD; controls were terminated non-malformed fetuses. Information on ma-ternal sociodemographic characteristics, lifestyle, and
folic acid supplementation was collected through
in-person interviews and by viewing medical records. Maternal venous blood samples were collected at preg-nancy termination. Fetal spinal cord and brain tissues were collected by autopsy performed by experienced pa-thologists. All samples were stored at−80 °C until assay. The study protocol was approved by the Institutional Review Board of Peking University, and written in-formed consent was obtained from all participating women.
Experimental animals
ICR mice of 8–9 weeks old were used, as described pre-viously [22]. Briefly, BaP (Sigma, USA) dissolved in corn oil was intraperitoneally administered into pregnant mice from E7 for four consecutive days (250–350 mg/ kg). Mice in the vitamin E co-exposure group were fed
with chow supplemented with the water-soluble
(±)-α-tocopherol succinate form of vitamin E (Sigma, USA) beginning from E0.5 (0.125%, w/w) and treated with BaP from E7 (250 mg/kg). On E10.5, embryos were collected and carefully inspected for NTDs. All experi-mental procedures were approved by the Institutional Animal Care and Use Committee of Peking University (certificate no. LA2013-36).
DNA methylation level analysis
DNA from neural tissues of human subjects was ex-tracted with QIAamp DNA Mini Kit (QIAGEN, Hilden, Germany) and E10.5 mouse DNA was extracted from frozen neural tissues using TIANamp Genomic DNA Kit (TIANGEN Biotech, Beijing, China). Then, 500 ng of genomic DNA from each sample was bisulfite-treated with EZ DNA methylation kit (Zymo Research, CA,
USA). The bisulfite conversion reaction was performed in duplicate for each sample to minimize potential bias caused by variable conversion efficiency, and pooled bisulfite-treated DNA was used for subsequent analysis. Human methylation study was performed in two phases. In phase 1, Infinium HumanMethylation450 BeadChip assay (450K, Illumina, San Diego, CA, USA) was used for genomic methylation assay of bisulfite-treated DNA from ten NTD cases and eight non-malformed controls, which has been described in detail in our previous study [23]. Methylation data ofPAX3gene was extracted from the array data and used for validation in the next phase.
In phase 2, 73 NTD cases and 29 non-malformed con-trol fetuses were included. The locations of the ampli-cons to target the aberrant CpG regions are shown in Fig. 1a. Bisulphite DNA was amplified by PCR and primers for thePAX3gene were designed using the on-line tool Epidesigner (www.epidesigner.com). Primer se-quences are listed in Additional file 5: Table S4. After reverse transcription, fragmentation, and analysis on a mass spectrometer (Sequenom, Inc., San Diego, USA), EpiTYPER Analyzer software was used for translating mass signal patterns into quantitative DNA methylation levels of different CpG sites.
Methylation analysis of mouse neural tissues was assayed with the same methods performed in phase 2 of the human methylation study. The regions of gene se-quence analyzed were the same as those in humanPAX3
gene and three amplicons were designed, with two in the promoter region and one in the body region, as shown in Fig.1b. See Additional file6: Table S5 for PCR primer sequences.
PAHs analysis
PAH concentrations in maternal blood were determined with an Agilent 7890A-5975C gas chromatograph and mass spectrometer equipped with a HP-5MS capillary column (30 m × 0.25 mm × 0.25μm), as described previ-ously [24]. In the present study, 51 mother-fetus pairs were available for PAH-methylation correlation analysis.
Oxidative stress assessment
The antioxidant capacity was determined by the reduced ferric reducing antioxidant power (FRAP) assay accord-ing to the manufacturer’s instructions (Beyotime Insti-tute of Biotechnology, China) for determining the level of TAC of mouse embryos. The TAC aims to measure both small molecule and protein antioxidants, including polyphenols, flavonoids, vitamins, and enzymes like
glutathione peroxidase and superoxide dismutase.
The absorbance of the mixture was measured at 593 nm using a Synergy 2 Multi-Mode Microplate Reader (Bio-Tek, USA) after incubation for 4 min. The TAC of each sample was calculated from the standard curve con-structed using FeSO4 solution, and the results were
expressed as mmol/g protein, adjusted according to the protein concentration of the samples. All samples were loaded in duplicate. Protein concentration was deter-mined using the BCA protein assay kit (Beyotime Insti-tute of Biotechnology, China).
SOD activity was determined using the total super-oxide dismutase assay kit with WST-8 (Beyotime Insti-tute of Biotechnology, China). Briefly, 20 μl of sample (E10.5 embryo samples were homogenized in lysis buf-fer) was mixed with 160 μl of WST-8/enzyme working solution. Then, 20μl of reaction triggering working solu-tion was added. After incubasolu-tion at 37 °C for 30 min, the absorbance was determined at 450 nm using a Syn-ergy 2 Multi-Mode Microplate Reader (BioTek, USA). The SOD activity was expressed as U/mg protein.
RNA isolation and real-time PCR
RNA was isolated from E10.5 embryos neural tissues using Trizol (Invitrogen); genomic DNA was removed by DNase I digestion (DNA-free, Ambion) and then reverse-transcribed using random hexamers (Superscript VILO cDNA synthesis kit). The abundance of mRNA of
Pax3were analyzed using real-time PCR (iTaq™ Univer-sal SYBR Green Supermix, BioRad) on a 7500 Fast Real Time PCR system (Applied Biosystems), with each sam-ple analyzed in triplicate. Primers are listed in Add-itional file 7: Table S6. Relative quantification of gene expression level was normalized according to theGapdh
gene expression.
Whole mount in situ hybridization
Whole mount in situ hybridization on E9.5 control em-bryos and emem-bryos treated with BaP was performed ac-cording to the procedures described by Yun et al. [49].
Pax3probe was cloned by real-time PCR into pGEM-T (Promega) and used to generate digoxygenin-labeled cRNA probes by reverse transcription using T7 RNA polymerase (Roche). For detection, anti-digoxigenin-AP antibody (1:2000, Roche) in 1% sheep serum was used and incubated overnight. Color detection was carried out using NBT/BCIP developing solution (Roche) in NTMT. After color development, embryos were imaged with a DFC490 camera (Leica), and then embedded.
Sec-tions of 40 μm thickness were obtained using a
vibratome.
Statistical analyses
In the human subject study, differences in proportions of population characteristics between NTD cases and
controls were examined with Pearson’s χ2
test. Inde-pendent ttest was performed to evaluate the difference in methylation of CpGs between NTDs and controls in phase 1, and adjusted for multiple testing with the Benjamini-Hochberg false discovery rate (FDR) methods. In phase 2, Shapiro-Wilk test was used to examine the distribution of methylation values of NTD cases and controls, and independent samples t test was used to identify CpG sites that were differentially methylated be-tween cases and controls. Odds ratio (OR) was calcu-lated by logistic regression to evaluate the association between higher methylation levels ofPAX3with the risk of NTDs, adjusting for maternal education, occupation, periconceptional folic acid use, fetal sex, and gestational age. Correlation between differentially methylated CpG sites in neural tissues and PAH concentrations in mater-nal serum was amater-nalyzed with Spearman’s correlation ana-lysis. In the mouse study, data on methylation level of
Pax3gene, oxidative stress markers, and the abundance of mRNA were presented as mean ± SE (SD). A one-way analysis of variance (ANOVA) followed by LSD (equal variances assumed) or Dunnett’s T3 (equal variances not assumed) was used for testing the differences between groups. A two-tail P value of < 0.05 was considered sta-tistically significant. Statistical analyses were conducted using SPSS 18.0.
Additional files
Additional file 1:Table S1. Characteristics of NTD cases and controls in phase 1 for methylation assay. (DOCX 18 kb)
Additional file 2:Table S2. Methylation ofPAX3gene in phase 1 using the HumanMethylation450 BeadChip assay. (DOCX 24 kb)
Additional file 3:Table S3. Correlation analysis of DNA methylation of
PAX3in fetal neural tissues and PAH concentrations in maternal serum (N
= 51 mother-fetus pairs). (DOCX 22 kb)
Additional file 4:Figure S1.PAX3methylation pattern assayed by Sequenom EpiTYPER in NTD cases with and without folic acid supplementation. (DOCX 148 kb)
Additional file 5:Table S4. The PCR primer sequences in Sequenom EpiTYPER sequencing. (DOCX 15 kb)
Additional file 6:Table S5. The PCR primer sequences for mouse methylation analysis. (DOCX 15 kb)
Additional file 7:Table S6. Sequences of primers for real-time PCR in mouse study. (DOCX 15 kb)
Abbreviations
BaP:Benzo[a]pyrene; FDR: False discovery rate; FRAP: Ferric reducing antioxidant power; HM450K: HumanMethylation450 BeadChip; NTDs: Neural tube defects; PAH: Polycyclic aromatic hydrocarbon; ROS: Reactive oxygen species; SOD: Superoxide dismutase; TAC: Total antioxidant capacity
Acknowledgements
Funding
This work was supported by the National Key Research and Development Program, Ministry of Science and Technology, the Peoples’Republic of China (Grant No. 2016YFC1000501), and grants from the National Natural Science Foundation of China (Grant No. 31371523 and 81773441), the Ministry of Education of China (Grant No. 20130001110064), and the UK Medical Research Council (N003713, K022741). NG is supported by Great Ormond Street Hospital Children’s Charity and by the National Institute for Health Research Biomedical Research Centre at Great Ormond Street Hospital for Children NHS Foundation Trust and University College London.
Availability of data and materials
The data sets supporting the results of this article are included within the article and its additional files.
Authors’contributions
AR and LW conceived and designed the study. SL performed the experiments, conducted data analyses, interpreted the data, and drafted the manuscript. YH and CS helped with the experiments. AR, LW, CS, and NG critically revised the paper. NG helped provide reagents, materials, and experimental infrastructure. LJ and ZL contributed to human subject recruitment. All authors read and approved the definitive version of the manuscript.
Ethics approval and consent to participate
The study protocol was approved by the Institutional Review Board of Peking University, and written informed consent was obtained from all participating women. The animal experiments were approved by the Institutional Animal Care and Use Committee of Peking University (certificate no. LA2013-36).
Consent for publication Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author details
1Institute of Reproductive and Child Health, National Health Commission Key
Laboratory of Reproductive Health, and Department of Epidemiology and Biostatistics, School of Public Health, Peking University Health Centre, Peking University, Beijing 100191, China.2Division of Birth Cohort Study, and
Department of Neonatal Surgery, Guangzhou Women and Children’s Medical Centre, Guangzhou Medical University, Guangzhou, China.3Developmental Biology and Cancer Programme, UCL Great Ormond Street Institute of Child Health, University College London, London WC1N 1EH, UK.
Received: 21 August 2018 Accepted: 8 January 2019
References
1. Mitchell LE. Epidemiology of neural tube defects. Am J Med Genet C Semin Med Genet. 2005;135c(1):88–94.
2. Greene ND, Stanier P, Copp AJ. Genetics of human neural tube defects. Hum Mol Genet. 2009;18(R2):R113–29.
3. Harris MJ, Juriloff DM. Mouse mutants with neural tube closure defects and their role in understanding human neural tube defects. Birth Defects Res A Clin Mol Teratol. 2007;79(3):187–210.
4. Wilde JJ, Petersen JR, Niswander L. Genetic, epigenetic, and environmental contributions to neural tube closure. Annu Rev Genet. 2014;48:583–611. 5. Greene ND, Stanier P, Moore GE. The emerging role of epigenetic
mechanisms in the etiology of neural tube defects. Epigenetics. 2011;6(7): 875–83.
6. Weinhold B. Epigenetics: the science of change. Environ Health Perspect. 2006;114(3):A160–7.
7. Morgan HD, Santos F, Green K, Dean W, Reik W. Epigenetic reprogramming in mammals. Hum Mol Genet. 2005;14 Spec No 1:R47–58.
8. Kim JK, Samaranayake M, Pradhan S. Epigenetic mechanisms in mammals. Cell Mol Life Sci. 2009;66(4):596–612.
9. Wang L, Wang F, Guan J, Le J, Wu L, Zou J, et al. Relation between hypomethylation of long interspersed nucleotide elements and risk of neural tube defects. Am J Clin Nutr. 2010;91(5):1359–67.
10. Wu L, Wang L, Shangguan S, Chang S, Wang Z, Lu X, et al. Altered methylation of IGF2 DMR0 is associated with neural tube defects. Mol Cell Biochem. 2013;380(1–2):33–42.
11. Bai B, Zhang Q, Liu X, Miao C, Shangguan S, Bao Y, et al. Different epigenetic alterations are associated with abnormal IGF2/Igf2 upregulation in neural tube defects. PLoS One. 2014;9(11):e113308.
12. Liu Z, Wang Z, Li Y, Ouyang S, Chang H, Zhang T, et al. Association of genomic instability, and the methylation status of imprinted genes and mismatch-repair genes, with neural tube defects. Eur J Hum Genet. 2012;20(5):516–20. 13. Shangguan S, Wang L, Chang S, Lu X, Wang Z, Wu L, et al. DNA
methylation aberrations rather than polymorphisms of FZD3 gene increase the risk of spina bifida in a high-risk region for neural tube defects. Birth Defects Res A Clin Mol Teratol. 2015;103(1):37–44.
14. Tian T, Wang L, Shen Y, Zhang B, Finnell RH, Ren A. Hypomethylation of GRHL3 gene is associated with the occurrence of neural tube defects. Epigenomics. 2018;10(7):891–901.
15. Rochtus A, Izzi B, Vangeel E, Louwette S, Wittevrongel C, Lambrechts D, et al. DNA methylation analysis of Homeobox genes implicates HOXB7 hypomethylation as risk factor for neural tube defects. Epigenetics. 2015; 10(1):92–101.
16. Phelan SA, Ito M, Loeken MR. Neural tube defects in embryos of diabetic mice: role of the Pax-3 gene and apoptosis. Diabetes. 1997;46(7):1189–97. 17. Greene ND, Massa V, Copp AJ. Understanding the causes and prevention of
neural tube defects: insights from the splotch mouse model. Birth Defects Res A Clin Mol Teratol. 2009;85(4):322–30.
18. Pingault V, Ente D, Dastot-Le Moal F, Goossens M, Marlin S, Bondurand N. Review and update of mutations causing Waardenburg syndrome. Hum Mutat. 2010;31(4):391–406.
19. Hol FA, Hamel BC, Geurds MP, Mullaart RA, Barr FG, Macina RA, et al. A frameshift mutation in the gene for PAX3 in a girl with spina bifida and mild signs of Waardenburg syndrome. J Med Genet. 1995;32(1):52–6. 20. Lu W, Zhu H, Wen S, Laurent C, Shaw GM, Lammer EJ, et al. Screening for
novel PAX3 polymorphisms and risks of spina bifida. Birth Defects Res A Clin Mol Teratol. 2007;79(1):45–9.
21. Ren A, Qiu X, Jin L, Ma J, Li Z, Zhang L, et al. Association of selected persistent organic pollutants in the placenta with the risk of neural tube defects. Proc Natl Acad Sci U S A. 2011;108(31):12770–5.
22. Lin S, Ren A, Wang L, Huang Y, Wang Y, Wang C, et al. Oxidative stress and apoptosis in benzo[a]pyrene-induced neural tube defects. Free Radic Biol Med. 2018;116:149–58.
23. Wang L, Lin S, Zhang J, Tian T, Jin L, Ren A. Fetal DNA hypermethylation in tight junction pathway is associated with neural tube defects: a genome-wide DNA methylation analysis. Epigenetics. 2017;12(2):157–65. 24. Wang B, Jin L, Ren A, Yuan Y, Liu J, Li Z, et al. Levels of polycyclic aromatic
hydrocarbons in maternal serum and risk of neural tube defects in offspring. Environ Sci Technol. 2015;49(1):588–96.
25. Fleming A, Copp AJ. Embryonic folate metabolism and mouse neural tube defects. Science. 1998;280(5372):2107–9.
26. Fleming A, Copp AJ. A genetic risk factor for mouse neural tube defects: defining the embryonic basis. Hum Mol Genet 2000;9(4):575–581. 27. Nye JS, Balkin N, Lucas H, Knepper PA, McLone DG, Charrow J.
Myelomeningocele and Waardenburg syndrome (type 3) in patients with interstitial deletions of 2q35 and the PAX3 gene: possible digenic inheritance of a neural tube defect. Am J Med Genet. 1998;75(4):401–8. 28. Agopian AJ, Bhalla AD, Boerwinkle E, Finnell RH, Grove ML, Hixson JE, et al.
Exon sequencing of PAX3 and T (brachyury) in cases with spina bifida. Birth Defects Res A Clin Mol Teratol. 2013;97(9):597–601.
29. Wei D, Loeken MR. Increased DNA methyltransferase 3b (Dnmt3b)-mediated CpG island methylation stimulated by oxidative stress inhibits expression of a gene required for neural tube and neural crest development in diabetic pregnancy. Diabetes. 2014;63(10):3512–22.
30. Zhong J, Xu C, Reece EA, Yang P. The green tea polyphenol EGCG alleviates maternal diabetes-induced neural tube defects by inhibiting DNA hypermethylation. Am J Obstet Gynecol. 2016;215(3):368. e1-.e10 31. Rochtus A, Winand R, Laenen G, Vangeel E, Izzi B, Wittevrongel C, et al.
factor with evidence for a complex (epi)genetic interplay to affect neural tube development. Clin Epigenetics. 2016;8:108.
32. Bell AC, Felsenfeld G. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature. 2000;405(6785):482–5. 33. Zhang X, Yazaki J, Sundaresan A, Cokus S, Chan SW, Chen H, et al.
Genome-wide high-resolution mapping and functional analysis of DNA methylation in arabidopsis. Cell. 2006;126(6):1189–201.
34. Lev Maor G, Yearim A, Ast G. The alternative role of DNA methylation in splicing regulation. Trends Genet. 2015;31(5):274–80.
35. Yearim A, Gelfman S, Shayevitch R, Melcer S, Glaich O, Mallm JP, et al. HP1 is involved in regulating the global impact of DNA methylation on alternative splicing. Cell Rep. 2015;10(7):1122–34.
36. Tang WY, Levin L, Talaska G, Cheung YY, Herbstman J, Tang D, et al. Maternal exposure to polycyclic aromatic hydrocarbons and 5’-CpG methylation of interferon-gamma in cord white blood cells. Environ Health Perspect. 2012;120(8):1195–200.
37. Herbstman JB, Tang D, Zhu D, Qu L, Sjodin A, Li Z, et al. Prenatal exposure to polycyclic aromatic hydrocarbons, benzo[a]pyrene-DNA adducts, and genomic DNA methylation in cord blood. Environ Health Perspect. 2012; 120(5):733–8.
38. Fang X, Dong W, Thornton C, Willett KL. Benzo[a]pyrene effects on glycine N-methyltransferase mRNA expression and enzyme activity in Fundulus heteroclitus embryos. Aquat Toxicol. 2010;98(2):130–8.
39. Sadikovic B, Rodenhiser DI. Benzopyrene exposure disrupts DNA methylation and growth dynamics in breast cancer cells. Toxicol Appl Pharmacol. 2006;216(3):458–68.
40. Dolinoy DC, Huang D, Jirtle RL. Maternal nutrient supplementation counteracts bisphenol A-induced DNA hypomethylation in early development. Proc Natl Acad Sci U S A. 2007;104(32):13056–61.
41. Bernal AJ, Dolinoy DC, Huang D, Skaar DA, Weinhouse C, Jirtle RL. Adaptive radiation-induced epigenetic alterations mitigated by antioxidants. FASEB J. 2013;27(2):665–71.
42. Franco R, Schoneveld O, Georgakilas AG, Panayiotidis MI. Oxidative stress, DNA methylation and carcinogenesis. Cancer Lett. 2008;266(1):6–11. 43. Angelini F, Pagano F, Bordin A, Milan M, Chimenti I. The impact of
environmental factors in influencing epigenetics related to oxidative states in the cardiovascular system. Oxid Med Cell Longev. 2017;2017:2712751. 44. Hitchler MJ, Domann FE. An epigenetic perspective on the free radical
theory of development. Free Radic Biol Med. 2007;43(7):1023–36. 45. Weitzman SA, Turk PW, Milkowski DH, Kozlowski K. Free radical adducts
induce alterations in DNA cytosine methylation. Proc Natl Acad Sci U S A. 1994;91(4):1261–4.
46. Chia N, Wang L, Lu X, Senut MC, Brenner C, Ruden DM. Hypothesis: environmental regulation of 5-hydroxymethylcytosine by oxidative stress. Epigenetics. 2011;6(7):853–6.
47. Cyr AR, Hitchler MJ, Domann FE. Regulation of SOD2 in cancer by histone modifications and CpG methylation: closing the loop between redox biology and epigenetics. Antioxid Redox Signal. 2013;18(15):1946–55. 48. Liu J, Zhang L, Li Z, Jin L, Zhang Y, Ye R, et al. Prevalence and trend of
neural tube defects in five counties in Shanxi province of northern China, 2000 to 2014. Birth Defects Res A Clin Mol Teratol. 2016;106(4):267–74. 49. Pai YJ, Leung KY, Savery D, Hutchin T, Prunty H, Heales S, et al. Glycine