O R I G I N A L PA P E R
The effect of electromagnetic radiation in the mobile
phone range on the behaviour of the rat
Willie M. U. Daniels&Ianthe L. Pitout& Thomas J. O. Afullo&Musa V. Mabandla
Received: 2 June 2009 / Accepted: 26 August 2009 #Springer Science + Business Media, LLC 2009
Abstract Electromagnetic radiation (EMR) is emitted from electromagnetic fields that surround power lines, household appliances and mobile phones. Research has shown that there are connections between EMR exposure and cancer and also that exposure to EMR may result in structural damage to neurons. In a study by Salford
et al. (Environ Health Perspect 111:881–883, 2003) the authors demonstrated the
presence of strongly stained areas in the brains of rats that were exposed to mobile phone EMR. These darker neurons were particularly prevalent in the hippocampal area of the brain. The aim of our study was to further investigate the effects of EMR. Since the hippocampus is involved in learning and memory and emotional states, we
hypothesised that EMR will have a negative impact on the subject’s mood and
ability to learn. We subsequently performed behavioural, histological and biochem-ical tests on exposed and unexposed male and female rats to determine the effects of EMR on learning and memory, emotional states and corticosterone levels. We found no significant differences in the spatial memory test, and morphological assessment of the brain also yielded non-significant differences between the groups. However, in some exposed animals there were decreased locomotor activity, increased grooming and a tendency of increased basal corticosterone levels. These findings suggested that EMR exposure may lead to abnormal brain functioning.
Keywords Electromagnetic radiation . Brain development . Behaviour . Hippocampus . HPA axis . Mood disorders
DOI 10.1007/s11011-009-9164-3
W. M. U. Daniels (*)
:
I. L. Pitout:
M. V. MabandlaDepartment of Human Physiology, University of KwaZulu-Natal, Westville Campus, Durban 4000, South Africa
e-mail: [email protected] T. J. O. Afullo
Department of Electrical, Electronic and Computer Engineering, University of KwaZulu-Natal, Westville Campus, Durban 4000, South Africa
Introduction
There is a growing interest in today’s society for the potential deleterious effects of
radio frequency electromagnetic fields to the health of the general public. More specifically, there is great concern for the effects of electromagnetic radiation (EMR) on the development and function of the brain, especially with the current widespread use of cellular telephones where Rf exposure is directly to the head. In 2006 WHO issued a Research Agenda for priority research into the effects of radio frequency fields. Among the research needs identified were pre- and post-natal animal studies for radio frequency exposure and its potential effects on cognitive performance.
Therefore the rationale for the present study was that “behavioural studies with
immature animals may provide a useful indicator of possible cognitive effects in
children”(WHO 2006Research Agenda for Radio Frequency Fields). Despite this
call, a recent search on Pubmed (24 August 2009) for studies relating to mobile phone exposure yielded only 623 publications since 1992. This statistic further emphasized the need for additional research.
Salford et al. (2003), found evidence that in rats, exposure to GSM 900 EMR,
caused structural damage to the brain. The hippocampus in particular showed the presence of darker, shrunken neurons in the pyramidal cell layer. These authors postulated that the damage was caused by albumin leakage from the blood brain barrier into the brain. In a previous study with EMR the same authors showed that radio frequency fields increase the permeability of the blood brain barrier which allows large plasma proteins like albumin to leak into the brain and cause structural
damage (Eberhardt et al.2008).
The hippocampus is an integral part of the brain’s limbic system and its
glucocorticoid and mineralocorticoid receptors are involved in behaviour regulation as well as regulation of the hypothalamo-pituitary adrenal (HPA) axis (Sapolsky et
al.1984; Gewirtz et al.2000; Gold et al. 2002; Ziegler and Herman2002; Kellner
and Wiedemann2008). It is also a site of long-term potentiation (LTP)—the cellular
mechanism believed to underlie learning and memory. Damage to neurons in the hippocampus may therefore lead to impaired learning and memory, behavioural disturbances, as well as negatively impact the functioning of the HPA axis.
Another aspect considered in our study was the effect EMR may have on brain development. Clinical and laboratory animal studies have shown that environmental conditions during early life can alter brain and behavioural development (Heim et al.
1997; Daniels et al. 2004). For instance children who have had traumatic
experiences show an increase in depressive disorders and anxiety disorders in
adulthood (Nemeroff2004; Wals and Verhulst2005). It appears therefore that brain
development, once altered, remains so for life. Basic studies using mostly rodents have also demonstrated early life trauma to affect behaviour in adulthood (Jezová et
al. 2002; Sullivan et al.2006). In a previous study (Daniels et al. 2004) we have
subjected rat pups to maternal separation for 3 h between postnatal day 2 and 14. This mild procedure resulted in significant long term changes in behaviour and brain chemistry. These findings suggested that the brain is vulnerable to environmental factors at this age, and that inflicted insults can cause long lasting damage to brain function. In view of these observations we wondered whether exposure to EMR, as an environmental factor, would yield similar results.
In the light of this, we exposed rat pups to EMR from post-natal day 2 to14 for 3 h per day. We hypothesized that exposure to EMR early in the life of rat pups will lead to abnormal brain development, impacting negatively on their behaviour during adulthood.
Materials and methods
Ethical clearance for this study was obtained from the University of KwaZulu-Natal
(Reference number 047/08/Animal). Male and femaleSprague-Dawleyrat pups from
post-natal day 1 (P1) were obtained from the Biomedical Resources Unit and housed under standard laboratory conditions. On P22 the rats were weaned and separated into 6 males and 6 females per group (control and exposure groups) and housed in the Biomedical Resources Unit until P58 when the behavioural assessments commenced. To aid the analysis of the behavioural assessments a video camera was used to record sessions. On P62 the rats were sacrificed by means of decapitation for the collection of plasma and brain tissue.
Electromagnetic radiation (EMR) exposure
The rat pups and their dames were exposed to EMR for 3 h per day from P2 to P14. A radio frequency signal generator (Rohde & Schwarz, Germany) was used to emit an 840 MHz signal via an antenna that directed the signal to the cages. Control animals underwent the same transportation, habituation and handling procedure without switching on the signal generator i.e. no EMR exposure.
The antenna was placed 2.5 m above the ground and 0.9 m away from the animal cage. It was positioned to emit directly towards the side of the cage. The radiation parameters were calculated as follows:
Power Transmitted; Pt¼ 4:2 dBm¼0:38 mW
Power Density; Pd;at r¼0:93 m from Transmit Antenna¼60mW
m2 From the Friis transmission formula; Pd ¼ 4GPtpr2
)G¼ 4pr 2 ð ÞPd Pt ¼ 4p0:932 ð ÞðP60x106Þ 0:38x103 ¼1:7162:345 dB The received power; Pr¼ð ÞPd s¼ 4GPtpr2
l2
4p ¼1:01x106W
¼1:01x103mW 29:96dBm
For 75Ωreceiver; E dBð mV=mÞ ¼109þPr¼79:04 dBmV=m
Note: the assumption is that the received power gain, G=1.716 dB, while the
transmit antenna gain is cancelled by the cable losses and impedance mismatch. Also the estimated received field strength is close to the calculated field strength due to a cellular phone operating in the proximity of the human head. For a typical cell
phone, operating at a frequency of 850 MHz, (wavelength, λ=0.353 m), the
dimension of the typical cell phone, D≈10 cm. The far field for rf radiation of
wavelengthλ, is related to the maximum dimensions, D, through the relation:
R>2D 2 l ¼
2 0ð Þ:1 2
Thus, for radiation below 5.67 cm, the operation would be in the near field, while distances greater than 5.67 cm would yield far-field radiation.
These parameters were frequently checked throughout the experiments. The device used for measurement is the HF32D, the high-frequency analyzer for frequencies in the band 800 MHz to 2.5 GHz. Manufacturers are Gigahertz Solutions, of Langenzenn, Germany. From a biological perspective, this frequency range is relevant because cellular phones, cordless phones, microwave ovens, Bluetooth and Wireless LAN fall
within this measurement range. This instrument measures the“peak”field strength or
power density at a given location in space. This is important since the field at any given point in space (especially closed areas, like indoors) is prone to fluctuations depending on the orientation of the antenna, the polarization of the received signal, and the reflections in the neighbourhood of the receiver.
Equal exposure to each rat pup cannot be guaranteed by our methodology. We did however make sure that the radiation was evenly spread throughout the cage by measuring the field strength at different positions and angles. The usual metal lids covering the cages were exchanged for plastic ones during exposure. We preferred not to remove the dam as we did not want to add another confounding factor (maternal separation) to the study. Since the pups and dams in the various groups were passive most of the time, we could arrange/position the cage in such a way that the pups were facing towards the antenna, anticipating direct exposure to the radiation.
The Morris water maze
The Morris water maze (MWM) was used to assess spatial memory and the ability to
learn a specific task (Morris 1984). The MWM was performed in a pool with
dimensions 240 cm (diameter) × 35 cm (height). A clear perspex platform was placed in the target quadrant 15 cm from the side of the pool and approximately 1 cm below the water surface. The temperature of the water was 22°C. The rats were towel dried on exiting the pool and then placed in pre-warmed cages with paper towel, where they were given a recuperation period of at least 30 min between trials. The MWM was performed as follows:
Trials 1–4: Each rat was placed in the start position, facing the side of the pool
and allowed to swim for 60 s. If the rat had not found the platform in the given time, it was picked up and placed on the platform for 10 s, allowing it to orientate itself. The rat was then dried and returned to its cage. This procedure was repeated 4 times for each rat.
Tests 1 and 2: Each rat was placed in the start position and allowed 60 s in which to
find the platform. The time taken to find the platform was recorded. The scores of test 1 and 2 were averaged to give an indication of the average time it took each individual rat to reach the platform. This
recording was used as an indicator of the rat’s learning capability.
Test 3: The platform was removed. The rat was placed in the start position
and allowed to swim for 60 s. The time spent in the target quadrant
was recorded as a test of the rat’s spatial memory i.e. the rat was
expected to spend more time in the target quadrant than the other 3 quadrants.
While executing the MWM tests, some animals exhibited freezing behaviour. The animals would simply float in the water with no apparent movement by any limb. The time spent in this behavioural state was subsequently recorded.
The open field test
The Open field test was used to assess anxiety-like behaviour and mood disturbances
(Prut and Belzung2003). The Open field used was a 1 m × 1 m square with sides of
50 cm high. It had demarcated inner and outer zones and the floor was divided into squares of equal size. In this study each rat was placed in the Open field for 5 min and the following parameters were assessed:
– Locomotor activity—the number of squares the rat crossed was counted and used as
a measure of locomotor activity. In anxious or stressed animals locomotor activity is abnormal. Some studies report an increase in locomotor activity in stressed animals
whilst others demonstrate it to be decreased (Marmendal et al.2006).
– Exploratory behaviour—stressed or anxious animals usually show a decrease in
exploratory behaviour when placed in a novel environment such as the Open field. Two parameters were assessed as an indication of exploratory behaviour: a) entries into the inner zone and b) rearing.
– Grooming—this activity was recorded as increased grooming may either
indicate relaxed behaviour or self-soothing behaviour exhibited by stressed, anxious animals. The time spent grooming was subsequently measured. Corticosterone determination
At the end of the behavioural tests, the rats were decapitated, their plasma was
collected, quickly frozen in liquid nitrogen and stored in a biofreezer at−80°C until
assaying. Blood collection always occurred between 09h00 and 11h00 in the morning to account for possible variations in circadian secretion of the steroid. Since this was an age controlled study, we assumed that both controls and exposed female rats were in a similar stage of their oestrous cycle, therefore these two groups were considered comparable. Corticosterone levels in the plasma were measured using a commercially available 96 well Enzyme Immunoassay (EIA) kit (Assay Designs, Ann-Arbor, USA). The standards were run in duplicate and the samples in triplicate. Histology
Standard histological techniques were performed to evaluate the integrity of the hippocampal neurons. Rats were decapitated, the brains removed from the skull and placed into 4% paraformaldehyde. The brains were blocked (~6 mm) and processed via the following steps:
i) Dehydration: the brain was dehydrated in ascending concentrations of alcohol
and cleared in Xylene.
ii) Infiltration: the brain tissue was infiltrated with melted paraffin (Shandon
iii) Sectioning: 5 µm sections were cut using a sledge mictrotome (Microm).
iv) Staining: Sections were stained with cresyl violet (Sigma-Aldrich, Missouri,
USA) and mounted with DPX resin. Statistical analysis
Graphpad Prism (V4) was used to analyse the data. In general a non-parametric test (Kruskal Wallis followed by the Man-Whitney test) was used to analyse the data. However since the majority of rats in the groups other than the exposed male group, did not display freezing behaviour, i.e. scored 0 s, it was considered more
appropriate to use ANOVA followed by Bonferroni’s Multiple Comparison test for
this data. Data is presented as mean ± SEM (n=6). p<0.05 was considered as
significant. Results Behaviour
There was no significant difference between any of the groups (male vs females; exposed vs unexposed controls) with the respect to the time taken to find the hidden
platform in the Morris water maze (Fig. 1a). Neither was there any significant
difference in time spent in the target quadrant (Fig.1b).
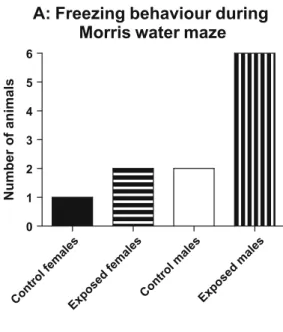
However all the male animals that were exposed to EMR displayed freezing behaviour in the MWM, whilst fewer animals exhibited this behaviour in the
other groups (Fig. 2a). The mean time spent exhibiting freezing behaviour of this
group was significantly more when compared to control males (Fig. 2b).
Immobilization in water typically reflects a mood disturbance and is used as a parameter for assessing depression and helplessness in the Porsolt forced swim test
(Porsolt et al. 1977). Since the freezing behaviour in our study was akin to
immobilization, we chose to further investigate the emotional states of the rats in the Open field test.
Male rats showed significantly less locomotor activity in the open field test when
compared to their unexposed controls (Fig.3). Male rats that were exposed to EMR
spent significantly more time grooming than any of the other groups (Fig.4). Overall
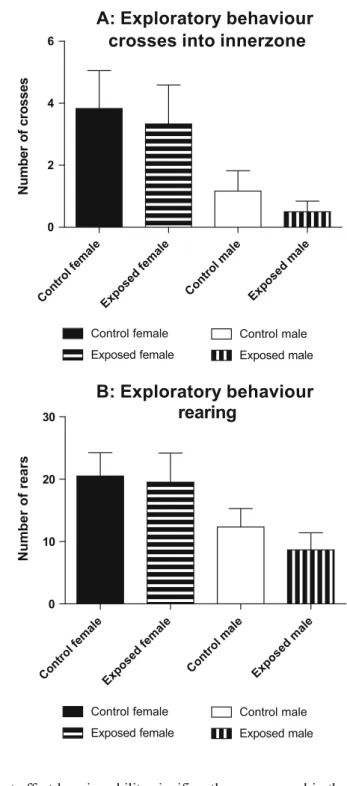
males displayed significantly less exploratory behaviour than females as evidenced by decreased crossings into the inner zone of the open field test and reduced rearings. However exposure to EMR had no significant effect on these parameters
(Fig.5a and b).
HPA axis activity
In general male rats had lower corticosterone concentrations than their female counterparts. The decrease was significant when control unexposed males were compared control unexposed females, but just missed significance between the
exposed groups (p<0.06). Exposure per se did not have any significant effect on
Histological assessment
Cresyl violet stained sections were analysed under a light microscope (Nikon DXM 1200C and Nikon AZ100) and the images captured with NIS-Element software (D 3.0). Morphological analysis of the hippocampal granular and pyramidal cells,
A: Time taken to find hidden platform
0 10 20 30 40 50
B: Time spent in target quadrant
0 10 20 30 40 Time (seconds) Time (seconds) Con trol females Exposed females Contro l m ales Exp osed ma les Con trol females Exposed females Contro l m ales Exp osed ma les Control females Exposed females Control males Exposed males Control females Exposed females Control males Exposed males
Fig. 1 Graphs depicting the average time taken for rats to find the platform (a) and the time spent in the target quadrant once the platform had been removed (b) in the testing session in the Morris water maze. Results represent mean ± SEM (n=6 per group)
and the cortical region revealed that the cells were intact and that there were no significant differences in neuron structure between control and EMR exposed brains.
Discussion
There is a growing concern for the potential deleterious health effects of radio frequency EMR exposure to the general public. Sources of these radiations in the environment include microwaves, power lines, telecommunication and broadcasting antennas and mobile phones. With the worldwide increase in mobile phone use
A: Freezing behaviour during
Morris water maze
0 1 2 3 4 5 6 Number of animals
B: Time spent exhibiting
freezing behaviour
0.0 2.5 5.0 7.5 10.0 12.5 15.0 17.5 Freezing Behaviour (in seconds) Con trol females Exposed females Control mal es Exp osed ma les Control fema les Exposed females Contro l m ales Exp osed ma les*
Fig. 2 Graph showing (a) the number of animals per group and (b) the mean time spent exhibiting freezing behaviour during the test trial in the Morris water maze. Statistical analysis revealed that exposed male rats displayed significantly more freezing behaviour than control males. *p<0.05, Significantly different from control males (ANOVA followed by Bonferroni’s Multiple Comparison test)
where EMR exposure is to the head, there is a need for research into the long term effects of EMR on the brain. Studies have shown that brain development is susceptible to mild insults during early life and that these onslaughts may affect
brain function later in life (Sullivan et al.2006). In the present study, we exposed
Sprague-Dawleyrat pups to electromagnetic frequency of 840 MHz equivalent to EMR exposure from mobile phones and obtained a number of interesting findings.
0 50 100 150 200 Locomotor Activity
No. of squares crossed
Control femaleExposed female Control mal e Exp osed ma le Control female Exposed female Control male Exposed male
Fig. 3 Graph depicting locomo-tor activity of the rats during the open field test. Exposure to electromagnetic radiation decreased locomotor activity in males when compared to unex-posed controls. Results represent mean ± SEM (n=6). *p<0.05, significantly different from control male group (Mann- Whitney test)
Grooming 0 10 20 30
*
Time spent grooming
(seconds)
Control femaleExposed female Contro l m ale Exp osed ma le Control female Exposed female Control male Exposed male
Fig. 4 Graph depicting the time spent grooming by various groups of animals. Exposure to electromagnetic radiation increased grooming behaviour in males when compared to unexposed controls. Results represent mean ± SEM (n=6). *p<0.05, significantly different from control male group (Mann- Whitney test)
Exposure to EMR did not affect learning ability significantly, as measured in the Morris water maze (MWM), in both gender groups. Whilst this result was considered unexpectedly, it may be possible that the time period of exposure was inadequate to impact this parameter significantly. We chose a time period of 3 h in
A: Exploratory behaviour
crosses into innerzone
Cont rol female Exposed female Control mal e Exp osed ma le Control fema le Exposed female Control mal e Exp osed ma le 0 2 4 6 Control female Exposed female Control male Exposed male Control female Exposed female Control male Exposed male Number of crosses
B: Exploratory behaviour
rearing
0 10 20 30 Number of rearsFig. 5 Graphs depicting exploratory behaviour of the various groups of rats during the open field test. Exposure to electromagnetic radiation did not affect inner zone crossings (a) or rearing (b) between any of the gender groups. Results represent mean ± SEM (N=6)
order to compare our observations with previous data on maternal separation
(Daniels et al.2004). The present result therefore indicated that these two paradigms
cannot be equated and hence a different exposure time ought to be considered in future experiments. Also it may be that the MWM is perhaps not sensitive enough to identify subtle changes in learning and memory especially in stressed animals
(D’Hooge and De Deyn 2001). We found in our study that the exposed animals
exhibited more freezing behaviour in the water than the unexposed counterparts and this became a confounding factor when analysing the parameters of time taken to find the platform and time spent in the target quadrant. Therefore whilst there were no significant differences in the MWM parameters, we remain cautious to conclude that EMR exposure did not affect learning and memory at all.
The marked increase in number of exposed animals that displayed freezing behaviour in the water and the amount of time spent doing so, prompted us to further investigate the behaviour of the rats. Immobilization is a parameter often used in the
Forced swim test to assess depression and helplessness (Porsolt et al.1977), thus the
freezing behaviour we observed suggested that there may be mood disturbances in our exposed animals. We performed an Open field test and assessed locomotor activity, exploratory behaviour and grooming. There was a significant decrease in the locomotor activity of the exposed male rats when compared to their unexposed controls. Whilst there were no differences in exploratory behaviour between the groups, exposed males spent more time grooming than their control counterparts. Decreased locomotor activity is usually indicative of disturbances in emotional states which may have been triggered by the stress of being placed in a novel environment
(Prut and Belzung 2003). Increased grooming is considered a feature of mood
disturbances and relates to self-soothing behaviour when faced with a stressful situation. The decrease in locomotor activity together with the increase in grooming therefore suggested that EMR exposure may lead to behavioural abnormalities at Plasma corticosterone concentration
Control femal e Exp ose d femal e Cont rol m ale Exposed ma le 0 2000 4000 6000 8000 10000 Control female Exposed female Control male Exposed male
*
Corticosterone levels (pg/ml)Fig. 6 Graph depicting the plasma corticosterone concentra-tion of the various groups. Whilst control males had significantly lower levels than control females, exposure to electromagnetic radiation did not affect the corticosterone levels within the gender groups. Results represent the mean ± SEM (n=6). *p>0.01, signifi-cantly lower than control female group (Mann- Whitney test)
least in males. This result is in agreement with a similar finding in a previous study where exposure to direct and indirect stress also caused a reduction in locomotor
activity and increased grooming behaviour (Daniels et al.2008).
Basal corticosterone concentrations were determined on the plasma of the animals to ascertain whether the activity of the HPA axis was altered. In contrast to what the behavioural assessment reflected, there were no significant differences in cortico-sterone levels between the groups, although a trend was noted for exposed female
rats to have higher corticosterone levels than the unexposed female controls (p<
0.06). Notwithstanding, the higher basal levels of corticosterone in the exposed females may partially indicate hyperactivity of the HPA axis or impaired negative feedback inhibition of corticosterone release. Such HPA axis abnormalities have been reported for animals that are chronically and perinatally stressed (Jezová et al.
2002). Interestingly it was noted that there were significant differences in basal
corticosterone levels between genders. Female rats showed higher corticosterone levels than the male rats which may be due to the hormonal differences in males and
females. These findings are in keeping with Jezová et al. (2002) that there is
increased HPA axis function in females when compared with males. Due to these gender differences interpretation of the results requires caution as it cannot be inferred that male rats experienced similar responses to stress levels or that the differences in stress levels is reflected by the behaviour of the male versus female rats. It is known that corticosterone levels in females may be influenced by the stage of their oestrous cycle. In view of this we specifically collected the blood in the morning at a time period during which the corticosterone concentration usually reaches its nadir and hence the impact of the oestrous cycle is minimal.
It has been proposed that EMR exposure of mobile phone frequency and intensity
causes damage to the hippocampus (Salford et al. 2003). Following cresyl violet
staining, Salford et al. (2003) reported the presence of shrunken, dark neurons
especially in the pyramidal cell layer. Our histological analysis of the hippocampal cell layers and the cortical layer showed, in contrast to the above mentioned study, that the cells had retained their integrity and there was no significant difference
between the control and exposed brains. Unlike the study of Salford et al. (2003) we
did not sacrifice the animals and collect their brain tissue immediately after exposure. In our experiments the animals were exposed during early childhood and only studied much later in adulthood. The extended time lapse between the period of exposure and the behavioural and histological assessments may therefore have allowed for compensation or repair in areas affected by the EMR exposure resulting in no obvious structural abnormalities under light microscopy.
Conclusion
Our study shows that EMR may cause behavioural changes as evidenced by a reduction in locomotor activity, increased grooming and freezing behaviour. These alterations in behaviour have been associated with animal models of stress-related disorders and therefore suggest that EMR exposure may be an environmental risk factor in the development of behavioural abnormalities. The present study recognises that the obtained evidence is limited, and that further investigation is required.
However it does alert us to the possibility that EMR exposure may have long term deleterious effects.
Acknowledgements The authors wish to thank the staff of the Biomedical Resource Unit of the University of Kwazulu-Natal for technical assistance.
References
Daniels WMU, Pietersen CY, Carstens ME, Stein DJ (2004) Maternal separation in rats leads to anxiety behaviour, and a blunted ACTH response and altered neurotransmitter levels in response to a subsequent stressor. Metab Brain Dis 19(1/2):13–24
Daniels WMU, Uys JDK, Van Vuuren P, Stein DJ (2008) The development of behavioral and endocrine abnormalities in rats after repeated exposure to direct and indirect stress. Neuropsychiatr Dis Treat 4 (2):451–464
D’Hooge R, De Deyn PP (2001) Applications of the Morris water maze in the study of learning and memory. Brain Res Rev 36(1):60–90
Eberhardt JL, Persson BR, Brun AE, Salford LG, Malmgren LO (2008) Blood-brain barrier permeability and nerve cell damage in rat brain 14 and 28 days after exposure to microwaves from GSM mobile phones. Electromagn Biol Med 27(3):215–229
Gewirtz J, McNish K, Davis M (2000) Is the hippocampus necessary for contextual fear learning? Behav Brain Res 110(1–2):83–95
Gold PW, Drevets WC, Charney DS (2002) New insights into the role of cortisol and the glucocorticoid receptor in severe depression. Biol Psych 52(5):381–385
Heim C, Owens MJ, Plotsky PM, Nemeroff CB (1997) The role of early adverse life events in the etiology of depression and posttraumatic stress disorder. Focus on corticotropin-releasing factor. Ann NY Acad Sci 821:194–207
Jezová D, Skultétyová I, Makatsori A, Moncek F, Duncko R (2002) Hypothalamo-pituitary-adrenocortical axis function and hedonic behaviour in adult male and female rats and prenatally stressed by maternal food restriction. Stress 5:177–183
Kellner M, Wiedemann K (2008) Mineralocorticoid receptors in brain, in health and disease: possibilities for new pharmacotherapy. Eur J Pharmacol 583(2–3):372–378
Marmendal M, Eriksson C, Fahlke CJ (2006) Early deprivation increases exploration and locomotion in adult male Wistar offspring. Pharmacol Biochem Behav 85(3):535–544
Morris R (1984) Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Methods 11:47–60
Nemeroff CB (2004) Early-life adversity, CRF dysregulation, and vulnerability to mood and anxiety disorders. Psychopharmacol Bull 38(1):14–20
Porsolt RD, Le Pichon M, Jalfre M (1977) Depression: a new animal model sensitive to antidepressant treatments. Nature 266(5604):730–732
Prut L, Belzung C (2003) The open field as a paradigm to measure the effects of drugs oon anxiety-like behaviours: a review. Eur J Pharmacol 463(1–3):3–33
Salford LG, Brun AE, Eberhardt JL, Malmgren L, Persson BR (2003) Nerve cell damage in mammalian brain after exposure to microwaves from GSM mobile phones. Environ Health Perspect 111(7):881–883 Sapolsky R, Krey L, McEwen B (1984) Glucocorticoid-sensitive hippocampal neurons are involved in
terminating the adreocortical stress response. PNAS 81:6174–6177
Sullivan R, Wilson DA, Feldon J, Yee BK, Meyer U, Richter-Levin G, Avi A, Michael T, Gruss M, Bock J, Helmeke C, Braun K (2006) The international society for developmental psychobiology annual meeting symposium: impact of early life experience on brain and behavioural development. Dev Psychobiol 48(7):583–602
Wals M, Verhulst F (2005) Child and adolescent antecedents of adult mood disorders. Curr Opin Psychiatry 18(1):15–19
WHO (2006) Research agenda for radio frequency fields. World Health Organisation, Genevahttp://www. who.int/pehemf/research/rf_research_agenda_2006.pdf
Ziegler D, Herman J (2002) Neurocircuitry of stress integration: anatomical pathways regulating the hypothalamic-pituitary-adrenal axis of the rat. Integr Comp Biol 42:541–551