Unicentre
CH-1015 Lausanne
http://serval.unil.ch
Year :
2019
Progressive Multiple Sequence Alignment with Indel Evolution
Maiolo Massimo
Maiolo Massimo, 2019, Progressive Multiple Sequence Alignment with Indel Evolution
Originally published at : Thesis, University of Lausanne
Posted at the University of Lausanne Op
Document URN : urn:nbn:ch:serval-BIB_D24577D3A8854
Droits d’auteur
L'Université de Lausanne attire expressément l'attention des utilisateurs sur le fait que tous les
documents publiés dans l'Archive SERVAL sont protégés par le droit d'auteur, conformément à la
loi fédérale sur le droit d'auteur et les droits voisins (LDA). A ce titre, il est indispensable d'obtenir
le consentement préalable de l'auteur et/ou de l’éditeur avant toute utilisation d'une oeuvre ou
d'une partie d'une oeuvre ne relevant pas d'une utilisation à des fins personnelles au sens de la
LDA (art. 19, al. 1 lettre a). A défaut, tout contrevenant s'expose aux sanctions prévues par cette
loi. Nous déclinons toute responsabilité en la matière.
Copyright
The University of Lausanne expressly draws the attention of users to the fact that all documents
published in the SERVAL Archive are protected by copyright in accordance with federal law on
copyright and similar rights (LDA). Accordingly it is indispensable to obtain prior consent from the
author and/or publisher before any use of a work or part of a work for purposes other than
personal use within the meaning of LDA (art. 19, para. 1 letter a). Failure to do so will expose
offenders to the sanctions laid down by this law. We accept no liability in this respect.
!
Département de biologie computationnelle
Progressive Multiple Sequence Alignment
with Indel Evolution
Thèse de doctorat ès sciences de la vie (PhD)
présentée à la
Faculté de biologie et de médecine
de l’Université de Lausanne
par
Massimo Maiolo
Master of Science SUPSI in Engineering
Jury
Prof. Jérôme Goudet, Président
Prof. Christophe Dessimoz, Directeur de thèse
Prof. Nicolas Salamin, Co-directeur
Dr. Maria Anisimova, Co-directeur
Prof. Olivier Gascuel, expert
Prof. Anna-Sapfo Malaspinas, expert
©
2018 by Massimo Maiolo. All rights reserved
Acknowledgments
“Gratitude is when memory is stored in the heart and not in the
mind.”
– Lional Hampton
The outcomes of this thesis could not have been attained without the many
people who have been involved in my life throughout the years. In different
ways these people have encouraged, mentored, guided, instructed, influenced
and loved me.
Firstly, I have great pleasure in acknowledging my gratitude to Maria
Anisi-mova and Manuel Gil, my PhD advisors, for supporting and assisting me
dur-ing the past four years. I have been fortunate to have been advised by you.
I want to express my gratitude to Lorenzo Gatti who gave me valuable
in-puts in many occasions. We have spent together many many hours coding and
debugging our software package. I am particularly indebted to Simone Ulzega
for helping me a lot especially with the STFT project. He is the colleague that
I have my morning ‘doppio espresso’ with. I would like to express my thanks
to Xiaolei Zhang, our brilliant Master student, for the great collaboration in
the 3D-DP algorithm design. I have appreciated the collaboration with Tiziano
Leidi and Diego Frei, for the impressive experience and for inspirational
dis-cussions with me regarding the code parallelisation.
I am grateful for conversations with Victor Garcia, Spencer Bliven and
Norman Juchler. They have been helpful in giving advises many times during
my time at ZHAW. I’m also glad to have worked with our master students, Ta
Cam Phuong Nguyesan and Jithin Mathew Peechatt.
The members at the Institute of Applied Simulations have contributed to my
professional development, they have been always very kind and friendly as well
as good collaborator. In particular, I am grateful to the administrative
assis-tants Rita Schoepfer Braschler, Simone Frei, Cyril Burkhard and Natyra Ajvazi
at ZHAW along with Susanna Bachmann at UZH, Livia Ioni and Alexandra
Cassoli at UNIL. They kept me organized and were always ready to help.
vi
ACKNOWLEDGMENTS
I am much obliged also to my former colleagues at SUPSI, I miss Melissa
Gajewsky for the heated discussions and the fun we had together, she has been
a special inspiration to me. I will forever be indebted to my former research
advisor Prof. Andrea Danani, I am very grateful for his scientific advise and
many insightful discussions and suggestions. I still think fondly of my time
spent in his lab. He has been the one letting me pursuing a career in research. I
really enjoyed the time spent with Andrea Bernasconi. The joy and enthusiasm
he has for scientific research was motivational for me. I earnestly thank him
for his influence on my professional life. I am especially thankful to Alberto
Vancheri for the excellent example he has provided as person and admirable
researcher. Alberto was and remains my best role model as scientist, mentor,
and teacher. I never encountered another person so clever and at the same
time so modest. He is extremely knowledgeable in just about everything, that’s
the reason I used to call him ‘Vancher-pedia’.
I especially thank my advisors, Prof. Christian von Mering at the
univer-sity of Zurich and Prof. Christophe Dessimoz at the Univeruniver-sity of Lausanne.
Also, the members of my oral defense committee deserve special thanks, Prof.
Olivier Gascuel, Prof. Anna-Sapfo Malaspinas, Prof. Nicolas Salamin and
Prof. Gerome Goudet. Prof. Andreas Wagner deserve my heartfelt thanks as
being an additional member of my oral exam committee at the University of
Zurich.
I have greatly appreciated also the contributions of time and suggestions of
Prof. Ziheng Yang. I would also like to acknowledge Prof. Marco Comini for
sharing the L
A
TEX template I used for writing this thesis. I acknowledge the
agency that funded my PhD studies. I was financial supported by the Swiss
National Science Foundation.
I would like to express my deepest gratitude to my mom Helen and my dad
Vincenzo. My hard-working parents have sacrificed their lives for my brother,
my sisters and myself. They instilled in me care for and curiosity in the world
around us. It is very sad that my mom left so soon, she would have been proud
of this important achievement. It would be inappropriate if I omit to mention
also my wife’s family, Rosa and Federico, who all have been supportive and
caring.
Nevertheless, the most special thank goes to my wife Teresa and my
daugh-ter Noemi. Definitely, Teresa is the right person for me. All these years,
Teresa has been a true and great supporter. She unconditionally love me
dur-ing my good and bad times. She has faith in me and my intellect, and she
always encouraged me in all possible ways from my Bachelor to my PhD
stud-ies. These past several years have not been an easy ride, both academically,
financially and personally. But she was always at my side. We have both
learned a lot from each other about life and this experience has strengthened
our relationship. Noemi, my darling little princess, she gave and still gives me
a reason to fight in the most difficult moments. Decisively, she is the light that
brightens my heart. I hope, I can give a better future now that I attained this
important milestone. I love you dearly and thank thoroughly for always
foster-ing me. Your presence was indispensable in a process that is often perceived
as considerably solitaire. My family has always been someone I can count on.
There are no words to convey how deeply I love you.
GOD, I am truly grateful for your exceptional love during this hard but
won-drous journey.
Massimo Maiolo
University of Lausanne
December 2018
To my wife
Teresa: for
your
uncondi-
tional love,
end-less support and con- stant
encourage-ment. You always believed in me and trusted
the decisions I made. You empowered me to
work hard for the things I aspire to achieve.
You are my perfect wife and mother of our
child. To my daughter Noemi: your smile is
the source of my joy and pride. You gave
me the energy I needed for dealing with
all this work. I love you all dearly. You
both are very special and lovely,
the most precious and
valu-able persons in my life. I
am truly thankful
for having
you!
Abstract
Here we present, for the first time, a frequentist progressive Multiple Sequence
Alignment (MSA) method based on a rigorous and explicit mathematical
for-mulation of insertions and deletions, namely the Poisson Indel Process (PIP).
Having designed our algorithm in the Maximum Likelihood (ML) framework
has enabled us to avoid the time-consuming Markov Chain Monte Carlo
sam-pling of alignments. Our proposed algorithm aligns two homology paths,
rep-resented by their corresponding MSAs in polynomial time by ML under the
PIP model (Maiolo et al. 2017). The procedure has been integrated into a
pro-gressive procedure that, traversing a given phylogenetic tree, produces at each
internal node the optimal pairwise MSA ending at the root with the alignment
of all the input sequences. The integration of PIP equations into a Dynamic
Programming (DP) approach is not straightforward since the marginal
like-lihood is non-monotonic in the alignment length. Therefore, to account for
the dependence on alignment length we have extended the DP matrices with
a third dimension.
In order to reduce the computational complexity, the algorithm predicts
candidate homologous segments for the purpose of filtering out non-promising
regions in the DP matrix prior to the effective alignment process. Although
our method has been strongly inspired by MAFFT, we have introduced a
number of improvements like for example the use of a multi-scale short-time
Fourier transform (STFT) for the automatic detection of candidate
homolo-gous patterns. Moreover, the use of the multiple-resolution STFT rather than
the Fourier transform improves the detection of homologous regions especially
in the presence of noise and in case of relative short patterns. We have also
defined a more sophisticated and general approach to generate logically sound
paths to connect homologous blocks and resolve overlaps between them.
To mitigate the intrinsic greediness brought by the progressive DP
ap-proach we have also implemented a Stochastic backtracking (Mueckstein et al.
2002) version of the algorithm under the PIP model. In this way, our
pro-gressive algorithm generates at each visited node a distribution of candidate
sub-optimal alignments. Aligning sub-optimal solutions increases the chances
to escape from local maxima and in our opinion provides a valid strategy to
reduce the progressive bias.
Finally, to account for the among-sites substitution rate variation (ASRV)
we have applied a Gamma distribution to all the rates, insertion and deletion
rates included. However, further analysis are still needed to investigate the
impact of ASRV on the inferred alignments. Our hope is that this feature
could mimic to some extent a long insertion, that is, an insertion of more than
a single character at a time.
The use of a sound mathematical model of indel, namely the Poisson Indel
Process model, is providing more realistic and accurate estimates of MSAs,
phylogenies and model parameters. As a consequence, our new algorithms will
allow not only more accurate phylogeny and alignment inference but it will
also facilitate the estimation of statistical supports of inferred tree partitions
and the ancestral reconstruction of insertions-deletions and substitution
his-tory. Our tool has been developed in a user-friendly software package and is
applicable to large genomic and metagenomic datasets.
R´
esum´
e
Nous pr´esentons ici, pour la premi`ere fois, une m´ethode d’alignement
progres-sif de s´equences multiples (MSA) bas´ee sur une formulation math´ematique
rigoureuse et explicite des insertions et des suppressions, `a savoir le proc´ed´e
Poisson Indel (PIP). Le fait d’avoir conu notre algorithme dans le cadre du
maximum de vraisemblance (ML) nous a permis d’´eviter l’´echantillonnage
fas-tidieux des alignements par la chane de Markov Monte Carlo. L’algorithme
que nous proposons aligne deux trajectoires homologiques, repr´esent´ees par
leurs MSA correspondantes en temps polynomial par ML sous le mod`ele PIP
(Maiolo et al. 2017). La proc´edure a ´et´e int´egr´ee dans une proc´edure
progres-sive qui, en traversant un arbre phylog´en´etique donn´e, produit `a chaque noeud
interne le MSA optimal par paires se terminant `a la racine avec l’alignement
de toutes les s´equences d’entr´ee. L’int´egration des ´equations PIP dans une
ap-proche de programmation dynamique (DP) n’est pas simple puisque la
prob-abilit´e marginale est non monotone dans la longueur de l’alignement. Par
cons´equent, pour tenir compte de la d´ependance `a la longueur d’alignement,
nous avons ´etendu les matrices DP avec une troisi`eme dimension.
Afin de r´eduire la complexit´e des calculs, l’algorithme pr´edit les segments
homologues candidats afin de filtrer les r´egions non prometteuses dans la
ma-trice DP avant le processus d’alignement efficace. Bien que notre m´ethode
soit fortement inspir´ee du MAFFT, nous avons introduit un certain
nom-bre d’am´eliorations comme par exemple l’utilisation d’une transform´ee de
Fourier multi-´echelle de courte dur´ee (STFT) pour la d´etection automatique
des mod`eles homologues candidats. De plus, l’utilisation de la STFT
mul-tir´esolution plutt que de la transform´ee de Fourier am´eliore la d´etection des
r´egions homologues, en particulier en pr´esence de bruit et dans le cas de
mo-tifs relativement courts. Nous avons ´egalement d´efini une approche plus
so-phistiqu´ee et plus g´en´erale pour g´en´erer des chemins logiquement sains pour
connecter des blocs homologues et r´esoudre les chevauchements entre eux.
Pour att´enuer l’avidit´e intrins`eque apport´ee par l’approche progressive de
la PD, nous avons ´egalement mis en uvre une version r´etrospective
stochas-tique (Mueckstein et al. 2002) de l’algorithme selon le mod`ele PIP. De cette
faon, notre algorithme progressif g´en`ere `a chaque nud visit´e une distribution
des alignements optimaux candidats. L’alignement de solutions
sous-optimales augmente les chances d’´echapper aux maxima locaux et, `a notre
avis, fournit une strat´egie valable pour r´eduire le biais progressif.
Enfin, pour tenir compte de la variation du taux de substitution entre les
sites (ASRV), nous avons appliqu´e une distribution gamma `a tous les taux,
taux d’insertion et de suppression inclus. Toutefois, une analyse plus
appro-fondie est encore n´ecessaire pour ´etudier l’impact de l’ASRV sur les trac´es
d´eduits. Nous esp´erons que cette caract´eristique pourrait imiter dans une
cer-taine mesure une insertion ” longue ”, c’est- `a-dire une insertion de plus d’un
caract`ere `a la fois.
L’utilisation d’un mod`ele math´ematique solide de l’indel, `a savoir le mod`ele
du processus de Poisson Indel, fournit des estimations plus r´ealistes et plus
pr´ecises des ASM, des phylog´enies et des param`etres du mod`ele. Par cons´equent,
nos nouveaux algorithmes permettront non seulement une phylog´enie et une
inf´erence d’alignement plus pr´ecises, mais ils faciliteront ´egalement l’estimation
des supports statistiques des partitions d’arbres inf´er´ees et la reconstruction
ancestrale des insertions-suppressions et de l’historique des substitutions. Notre
outil a ´et´e d´evelopp´e dans un progiciel convivial et est applicable aux grands
ensembles de donn´ees g´enomiques et m´etag´enomiques.
Contents
List of Symbols
xxvii
List of Acronyms
xxxv
Introduction
xxxvii
Main result of the thesis
lv
Outline
lvii
I
Progressive Multiple Sequence Alignment with
In-del Evolution
1
1 Progressive Dynamic Programming under PIP model
3
1.1
Introduction
. . . .
3
1.2
Likelihood computation of a column
p
(
c
)
. . . .
5
1.3
Likelihood computation of a column
p
(
c
∅)
. . . .
7
2 3D Dynamic Programming under PIP
11
2.1
Introduction
. . . 11
2.2
Alignment at node
v
3
. . . 14
2.3
Alignment at node
v
5
. . . 20
2.4
Alignment at node Ω
. . . 24
2.5
Tracebacking
. . . 25
2.6
Early stop condition
. . . 27
3 Stochastic backtracking DP algorithm
29
3.1
Introduction
. . . 29
3.2
Partition functions
. . . 30
3.3
The partition functions
Z
M
,
Z
X
and
Z
Y
. . . 31
3.4
Forward recursion
. . . 33
3.5
Backward recursion
. . . 38
xvi
Contents
4 Marginal Likelihood with Rate Variation Across Sites
49
4.1
Introduction
. . . 49
4.2
PIP equations under ASRV
. . . 51
5 Multi-scale STFT based homologous blocks detection
55
5.1
Introduction
. . . 55
5.2
Fourier transform
. . . 58
5.3
Short-time Fourier transform
. . . 59
5.3.1
The Heisenberg Uncertainty Principle
. . . 62
5.4
Algorithm overview
. . . 63
5.4.1
Sequence residues to signal conversion
. . . 68
5.4.2
Signal padding
. . . 70
5.4.3
Fourier transform based homology detection
. . . 71
5.4.4
Multi-scale STFT based homology detection
. . . 73
5.4.5
Homology matrix and optimal path
. . . 83
5.4.6
Final alignment
. . . 91
5.5
STFT vs. FT approach for block detection
. . . 93
5.5.1
Noise sensitivity
. . . 93
5.5.2
Pattern length sensitivity
. . . 94
6 Progressive bias analysis
97
6.1
Introduction
. . . 97
6.2
Alignment with unbalanced tree
. . . 99
6.2.1
Global alignment
. . . 99
6.2.2
Progressive alignment
. . . 101
6.2.3
Progressive bias with unbalanced tree
. . . 102
6.3
Alignment with balanced tree
. . . 104
6.3.1
Global alignment
. . . 104
6.3.2
Progressive alignment
. . . 104
Discussion & Conclusions
107
II
Appendices
113
A Some technicalities
115
A.1 Detailed derivation of the marginal likelihood function
ϕ
(
v
)
. . . 115
A.2 Detailed derivation of the survival probability function
β
(
v
)
. . 120
Contents
xvii
B Reversibility of TKF91 and PIP
125
B.1 Introduction to TKF91
. . . 125
B.2 Time-reversible evolutionary process
. . . 128
B.3 Reversibility of TKF91
. . . 129
B.4 Reversibility of PIP
. . . 133
C Characterization of Indel rates
137
C.1 Introduction
. . . 137
C.2 lnferring indel rates from a given MSA
. . . 139
D Doob-Gillespie method and PIP description
145
D.1 Doob-Gillespie method
. . . 145
D.1.1 When does the next event happen?
. . . 146
D.1.2 What kind of event happens next?
. . . 148
D.2 Local PIP description
. . . 148
E Grantham’s distance
151
E.1 Introduction
. . . 151
E.2 Grantham’s distance computation
. . . 152
F Homologous blocks overlap resolution
159
G Multiple sequence alignment evaluation
165
G.1 Overview
. . . 165
G.2 Benchmarks
. . . 167
Bibliography
173
III
Journal article
189
Progressive multiple sequence alignment with indel evolution
. . . 191
List of Figures
1
Tree of life.
. . . xxxviii
2
Genetic variations at the molecular level.
. . . .
xl
3
DNA, RNA and protein synthesis.
. . . xlii
4
MSA and corresponding homology paths.
. . . xlv
5
Progressive Dynamic Programming.
. . . xlviii
1.1
Rooted topology used to illustrate the PIP-DP formulation.
. .
4
2.1
Phylogenetic tree
τ
.
. . . 12
2.2
Four three-dimensional sparse DP matrices.
. . . 12
2.3
An example of
ϕ
(
|
m
|
).
. . . 13
2.4
Cells computed at layer
m
= 0.
. . . 14
2.5
Homology paths at layer
m
= 0.
. . . 16
2.6
Cells computed layer
m
= 1.
. . . 17
2.7
Homology path representing a match.
. . . 18
2.8
Homologous scenarios at the node
v
3. 1.
. . . 19
2.9
Homologous scenarios at the node
v
3
. 2.
. . . 19
2.10 Cells computed at layer
m
= 2.
. . . 21
2.11 Homology paths at node
v
5. 1.
. . . 22
2.12 Homology paths at node
v
5. 2.
. . . 23
2.13 Homology paths at node
v
5
. 3.
. . . 24
2.14 Homology paths at node
v
5. 4.
. . . 24
2.15 Homology paths at the root Ω.
. . . 25
2.16 Possible starting points of the traceback phase.
. . . 26
2.17 Cells with likelihood different from 0.
. . . 27
2.18 Likelihood values of the last column.
. . . 28
3.1
Six partition functions.
. . . 33
3.2
Paths connected to
P
M
(3
,
3
,
4).
. . . 34
3.3
Cell dependencies in the 3D-DP matrices.
. . . 34
3.4
Possible homologous paths at
Z
M
(3
,
3
,
4).
. . . 37
3.5
Possible homologous paths at
Z
X
(3
,
3
,
4).
. . . 38
3.6
Possible homologous paths at
Z
Y
(3
,
3
,
4).
. . . 39
3.7
Last column in the 3D-DP matrices.
. . . 40
xx
List of Figures
3.9
Probability distortion as function of the temperature.
. . . 44
3.10 Probability distortion at
T
= 0
.
1.
. . . 45
3.11 Probability distortion at
T
= 0
.
3.
. . . 45
3.12 Probability distortion at
T
= 10.
. . . 46
3.13 Stochastic backtracking paths at different temperatures.
. . . . 47
4.1
Gamma probability distribution functions.
. . . 51
5.1
Theoretical speed-up as function of the number of blocks.
. . . . 57
5.2
Windowed signal.
. . . 60
5.3
Tiling of the time-frequency space.
. . . 63
5.4
FT coefficients
f
k
.
. . . 65
5.5
Column-gap content.
. . . 66
5.6
Overlapping and non-overlapping blocks.
. . . 67
5.7
Padding scheme with
S
w
= 64.
. . . 71
5.8
Padding scheme with
S
w
= 32.
. . . 71
5.9
Window functions examples.
. . . 74
5.10 Boundaries effects at different resolution scales.
. . . 75
5.11 STFT spectrogram.
. . . 76
5.12 First analysis at two different window size.
. . . 77
5.13 Window functions at different resolution level.
. . . 78
5.14 Spectrogram slice at different sliding length.
. . . 79
5.15 Noise threshold convergence.
. . . 80
5.16 Multi-scale STFT surfaces.
. . . 81
5.17 Blocks boundaries analyzed at different scales.
. . . 82
5.18 Multi-resolution boundary detection.
. . . 84
5.19 Example of overlapping blocks.
. . . 85
5.20 Tree of blocks representing possible paths.
. . . 86
5.21 Example of overlapping blocks and their resolution. i).
. . . 87
5.22 Example of overlapping blocks and their resolution. ii).
. . . 87
5.23 Example of overlapping blocks and their resolution. iii).
. . . 88
5.24 Example of overlapping blocks and their resolution. iv).
. . . 88
5.25 Overlap-free paths.
. . . 89
5.26 Homologous and linking blocks.
. . . 90
5.27 Magnification of connecting corner.
. . . 90
5.28 Homologous and linking blocks and magnification.
. . . 90
5.29 MSA obtained with/without STFT.
. . . 92
5.30 Noisy pattern experiment.
. . . 93
5.31 Noise sensitivity.
. . . 94
List of Figures
xxi
5.33 Pattern length.
. . . 95
6.1
Topologies used to quantify the progressive bias.
. . . 98
6.2
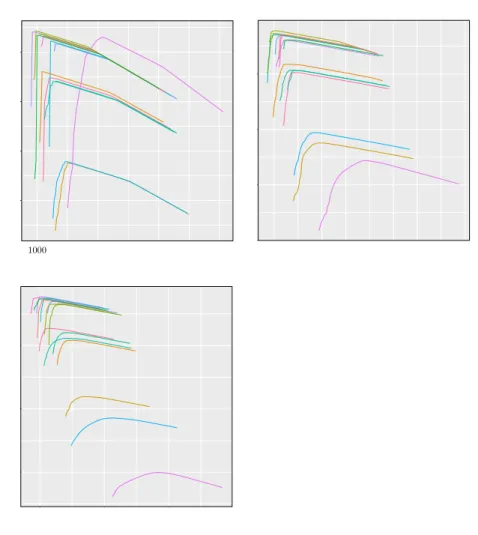
Marginal likelihood curves at the root node Ω.
. . . 101
6.3
Marginal likelihood curves at the root node
v
2.
. . . 103
6.4
Marginal likelihood curves at the root node
v
1.
. . . 103
6.5
Marginal likelihood curves at node
v
1.
. . . 105
6.6
Marginal likelihood curves at the root Ω.
. . . 106
6.7
Progressive bias analysis.
. . . 106
A.1 Graphical representation of
U
and
W
.
. . . 120
B.1 Time reversibility.
. . . 129
B.2 Four pairwise alignments.
. . . 131
B.3 Two pairwise alignments.
. . . 132
B.4 Topology used to test the reversibility of PIP.
. . . 133
B.5 Representation of likelihood under PIP.
. . . 135
C.1 Possible fate of characters.
. . . 138
C.2 Number of characters through time.
. . . 140
C.3
n
H,
n
G
and
n
H
+
n
G
as function of
I
. . . 142
C.4
n
H,
n
G
and
n
H
+
n
G
as function of
τ
. . . 142
C.5
n
H,
n
G
and
n
H
+
n
G
as function of
λ
. . . 143
C.6
n
H,
n
G
and
n
H
+
n
G
as function of
µ
. . . 143
D.1 Doob-Gillespie method under PIP. Step 1.
. . . 150
D.2 Doob-Gillespie method under PIP. Step 2.
. . . 150
D.3 Doob-Gillespie method under PIP. Step 3.
. . . 150
D.4 Doob-Gillespie method under PIP. Step 4.
. . . 150
E.1 Standardized and non standardized volumes.
. . . 153
E.2 Cross-correlation vs. Grantham’s distance approach.
. . . 154
E.3 Grantham’s distance coefficients.
. . . 157
F.1 Overlaps table.
. . . 163
G.1 BALiBASE benchmark dataset RV911-115: phylogenetic tree.
. 168
G.2 Evolutionary vs Structural alignment. Columns (1-200).
. . . . 168
G.3 Evolutionary vs Structural alignment. Columns (201-400).
. . . 169
G.4 Evolutionary vs Structural alignment. Columns (401-600).
. . . 169
G.5 Evolutionary vs Structural alignment. Columns (601-800).
. . . 170
xxii
List of Figures
G.7 Evolutionary vs Structural alignment. Columns (1001-1200).
. . 171
List of Tables
5.1 Amino-acids physiochemical properties.
. . . 69
6.1 MSAs potentially be generated at node
v
1.
. . . 99
6.2 MSAs potentially be generated at node
v
2.
. . . 99
6.3 MSAs potentially be generated at the root Ω.
. . . 100
List of Algorithms
1
Doob-Gillespie-PIP procedure
. . . 149
2
Overlap resolution procedure
. . . 160
3
Create block tree procedure
. . . 160
4
Expand node procedure
. . . 161
5
Resolve all overlaps procedure
. . . 161
6
Resolve single path procedure
. . . 161
7
Resolve pairs procedure
. . . 162
8
Resolve overlap pairs procedure
. . . 162
List of Symbols
H
(
v
) denotes an homology path of a single character generated by a
substi-tution-deletion continuous-time Markov Chain process along a the
phy-logeny. 123
L ρ
(
v
j
, v
k
)
length of the path that connects
v
j
to
v
k
. Corresponds to the sum
of branch length
b
(
v
) for all the edges connecting the two end points (
v
j
and
v
k
). 3
Γ symbol for the gamma distribution function. The density probability
func-tion is defined as
f
(
x
) =
θ
k
Γ(k)
1
x
k
−
1
e
−
x
θ
=
β
α
Γ(α)
x
α
−
1
e
−
βx
with (
k, θ
) and
(
α, β
) positive numbers. 51
Ω phylogenetic tree root. The root represents the most recent common
ances-tor of all of the taxa in the tree. 52–54, 123
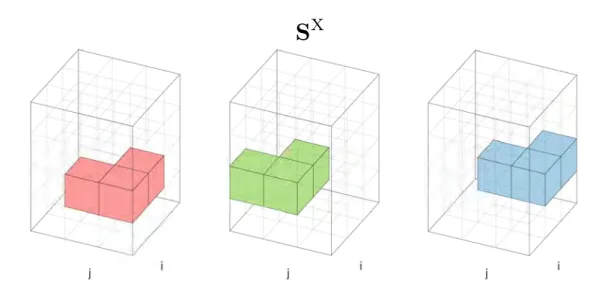
S
M
sparse 3-dimensional Dynamic Programming matrix. It contains the
like-lihood values for
match
state. xxvii, 14, 16, 18, 27
S
M,X,Y
short notation for
S
M
,
S
X
and
S
Y
. 16, 18, 21, 26–28
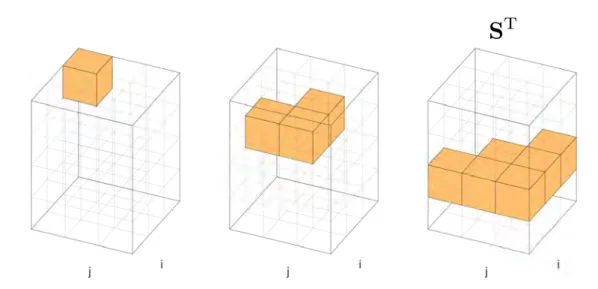
S
T
sparse 3-dimensional Dynamic Programming traceback matrix. It stores at
each position (
i, j, k
) which matrix contains the highest likelihood value.
By convention the index 1 stands for
S
M
, 2 for
S
X
and 3 for
S
Y
. 14,
25–27
S
X
sparse 3-dimensional Dynamic Programming matrix. It contains the
like-lihood values for
gapX
state. xxvii, 14, 16, 23, 27
S
Y
sparse 3-dimensional Dynamic Programming matrix. It contains the
like-lihood values for
gapY
state. xxvii, 14, 16, 27
k
ν
k
normalizing measure for the Poisson process. It corresponds to
k
ν
k
=
λ
k
τ
k
+
λ
µ
. 17, 18, 20, 116, 139, 140
k
τ
k
tree length obtained summing up all the branch lengths
b
(
v
) for
v
∈ V
.
xxxiii, 6, 18, 52, 116, 133, 134, 138–141
xxviii
List of Symbols
α
(
v
) corresponds to the sum of all the prior insertion probabilities
ι
(
v
) and
survival probabilities
β
(
v
) from the root to a particular node
v
on the
path
ρ
(
v,
Ω). It follows that for
v
∈ V
:
α
(
v
) =
k
τ
k
1/µ
+1/µ
. 7
∗
hadamard product between two arrays. The hadamard product between two
arrays
x
and
y
of same size corresponds to (
x
∗
y
)(
i
) =
x
(
i
)
·
y
(
i
). 5, 17,
22, 23, 133
β
(
v
) survival probability associated to the node
v
given an insertion, on a
random location, on the edge (pa
→
v
) of length
b
(
v
). See Appendix
A.2
for its detailed derivation.. xxviii, 4, 120, 123
π
extended steady state frequencies. It is represented as an array of size
|A
|×
1 obtained as the quasi-stationary distribution of
Q
.
π
(
i
) contains the
background frequency of the character at the position
i
in the extended
alphabet
A
. The steady state probability of a gap is always 0. See also
π
. 17, 133–135, 150
π
steady state frequencies. For instance using a nucleotide alphabet, the
stationary frequencies,
π
A
,
π
C
,
π
G
,
π
T
correspond to equilibrium base
compositions of the four states. The background frequencies are stored
into an array of size
|A| ×
1 and computed as the limiting distribution
when
t
→ ∞
, that is
π
exp(
Q
t
) =
π
. xxviii
◦
dot product between two arrays. The scalar (dot) product between two
arrays
x
and
y
of same size corresponds to
x
◦
y
=
P
i
x
(
i
)
·
y
(
i
). 5, 15,
17, 20–24, 52, 54, 133
δ
Ω
(d
t
) (continuous) dirac function defined on the topology. It returns 1 at the
root node and 0 elsewhere. 150
it means ‘putatively homologue to’. Hence, A
i
B
j
means that A
i
is
(puta-tively) homologue to B
j
. 16, 18, 19, 22, 31, 32, 36, 85
it denotes a gap state which is an absorbing state in the Markov chain under
the PIP model. It means that once the system enters in gap state it
cannot change anymore, or in other words, a gap never gives birth to
any character. 15, 123
η
(
v
) recursive formula associated to the node
v
that computes the character
deletion probability on the sub-tree rooted at node
v
. 8
List of Symbols
xxix
ι
(
v
) prior probability of a single character insertion on the branch associated
to
v
with
P
v
∈V
ι
(
v
) = 1. The prior insertion probability of a character
along the edge belonging to
v
∈ V \
Ω is proportional to its length
b
(
v
).
xxviii
λ
single character insertion rate.
λ
is kept constant during the entire
evolutio-nary process (static or time-invariant parameter) and does not depend
on the particular character inserted. 116, 138–141
E
denotes the expectation function. 50, 51, 115
P
symbol for the probability function. 126
Q
extended substitution rate matrix with an extra column and an extra row
to account for the gap character (absorbing state
). With the extended
alphabet of nucleotide for instance
Q
has dimension of 5
×
5. See also
Q
. 52, 123
Q
infinitesimal generator matrix completely describing the Markov process.
Q
contains the rates a continuous time Markov chain moves between states.
Therefore
Q
i,j
is the rate at which character
i
mutates into character
j
.
By using the nucleotide for instance
Q
is a matrix of size 4
×
4. xxix
X
set of input sequences. A sequence is a string of characters belonging to a
given alphabet
A
. Typically there are three kind of standard alphabets:
nucleotides, amino-acids or codons alphabet. 115
Z
M,X,Y
short notation for
Z
M
,
Z
X
and
Z
Y
. 33, 36, 38, 41, 43
Z
M
sparse 3-dimensional dynamic matrix containing the partition function of
alignments ending in state
match
. 31, 32, 35, 36, 40
Z
X
sparse 3-dimensional dynamic matrix containing the partition function of
alignments ending in state
gapX
. 31, 32, 36, 37
Z
Y
sparse 3-dimensional dynamic matrix containing the partition function of
alignments ending in state
gapY
. 31, 32, 36, 37
Z
total partition function obtained summing up
Z
M
,
Z
X
and
Z
Y
. 35
A
extended alphabet. It is defined as
A
=
A ∪
where
is an absorbing
state (gap character). Under PIP a gap is considered as an extra state
added to the canonical ones. See also
A
. xxviii, xxxii, liii, 5, 123
xxx
List of Symbols
A
represents the alphabet. A sequence is commonly represented as a string of
characters belonging to a given alphabet. The three alphabets commonly
used are of type
nucleotides
,
amino-acids
or
codons
. For instance the
nucleotide alphabet contains the four letters
{
A, T, G, C
}
. xxix, xxxi,
liii, 3, 123, 133
B
set of candidate homologous blocks detected by the Fourier Transform or the
short-time Fourier Transform. Blocks are then aligned as independent
DP sub-problems.. 83, 159
E
set of edges. An edge is a link connecting
v
with pa(
v
) and has associated
a length indicated as
b
(
e
) =
b
(
v
). xxxi, xxxiii
I
set of vertices
I ∪V
where an insertion could have happened. It corresponds
to the set of vertices having a felsenstein’s weight different from 0. liv,
52, 123
L
set of leaves where
L ⊂ V
. Leaves are nodes without children. The input
sequences are associated at the leaves before aligning them for instance
with a progressive DP procedure. 8, 52, 123
P
set of candidate paths between candidate homologous blocks. Prior to
in-dependently align the single blocks an algorithm has to connect logically,
all the blocks belonging to a path
P
that respect the order of the columns
and avoid any duplication.. xxxii, 83, 86, 159, 160
T
nth tree that represents all possible paths connecting in a meaningful way
the detected homologous blocks.. 83
U
(0
, t
) uniform (continuous) distribution on the interval (0
, t
), with
t >
0. 120
V
finite subset of vertices (branching points in the topology) where
V ⊂
τ
, in
fact under PIP the topology is considered as a continuous set of points.
52, 54, 116, 123
W
(
µ
) exponential distribution with parameter
µ
, describes for instance the
(waiting) time between consecutive events in a Poisson point process.
120
W
L
denotes the Lambert-W function. The Lambert-W function is a set of
functions (branches) of the function
f
(
z
) =
z
exp(
z
) where
z
is a complex
number, therefore
z
=
f
−
1
(
z
exp(
z
)) =
W
L
(
z
exp(
z
))
.
For any complex
number
z
0
=
z
exp(
z
) we get
z
0
=
W
L
(
z
0
) exp
W
L
(
z
0
)
List of Symbols
xxxi
1
indicator function. The indicator function returns the value 1 if the function
argument is true, 0 otherwise. liv, 3–5, 52
−
character symbol for the gap character, its state is denoted with the simbol
. 15
Poi stands for Poisson probability/distribution. 137, 138, 150
child(
v
) child node of
v
, the next node (below), directed linked to
v
. The
topologies considered here are always binary trees, it follows that each
internal node
v
∈ V \
Ω has degree 3 (2 children). The root Ω has
degree 2 (no parent node) and leaves do not have any children (degree
1). Accordingly, pa(child(
v
)) =
v
for
v
∈ V \
Ω. xxxi
pa(
v
) parent node of
v
, the next node (above), directed linked to
v
. Only the
root Ω does not have any parent node. It follows that child(pa(
v
)) =
v
for
v
∈ V \
Ω (see also child(
v
)). 3, 4
µ
single character deletion rate.
µ
is kept constant during the entire
evolutio-nary process (static or time-invariant parameter) and does not depend
on the particular character deleted. 116, 120, 121, 126, 130, 132, 140,
141
ν
(d
t
) Poisson process intensity defined on the topology, the latter considered
a continuous set of point. Under PIP this intensity is defines as
ν
(d
t
) =
λ
·
τ
(d
t
) +
µ
1
·
δ
Ω
(d
t
)
. 150
ρ
(
v
j
, v
k
) it denotes the set of nodes in the path
v
j
to
v
k
where
v
j
, v
k
∈ V
.
A path is a connected sequence of edges
e
∈ E
and is defined on the
topology. 3
\
the set difference operator. It is defined as
A
\
B
=
{
x
:
x
∈
A
and
x /
∈
B
}
.
27, 52–54, 123
σ
represent a character from the set of characters of a given alphabet
A
or
A
. 123
τ
(d
t
) Lebesgue measure on the topology. Its value corresponds to the distance
measured from a given point to the root. 150
τ
phylogenetic tree. Under PIP the phylogeny
τ
is a continuous set of points,
its topology is denoted by (
L
,
E
), where
V ⊂
τ
is equal to the finite
subset of vertices (branching points), the leaves
L ⊂ V
and the root Ω,
and where
E
is the set of edges. 11, 51, 115, 138, 141
xxxii
List of Symbols
θ
is the collection of model parameters (branch lengths, substitution rate
ma-trix, insertion/deletion rate, . . . ). 126, 127, 130
ϕ p
(
c
∅
)
,
|
m
|
marginal likelihood of non-observable empty columns for an
a-lignment of length
|
m
|
where
p
(
c
∅
) is the likelihood of a single MSA
column full of gaps. See Appendix
A.1
for its detailed derivation. 53,
115, 116
|A
|
cardinality (number of elements) of the (extended) alphabet
A
. For the
nucleotide alphabet, for instance,
|A
|
= 5. See also
A
. 5
|B|
cardinality of the set
B
which corresponds to the total number of
homol-ogous blocks detected by the Fourier Transform or short-time Fourier
Transform. 83
|
m
|
number of (observed) columns in an alignment
m
. An alignment
m
con-tains
n
columns, where
|
m
|
are observable and
n
− |
m
|
are full of gaps
and hence in general are not represented in an alignment. 13, 14, 17, 21,
51, 115–117, 140
b
Z
M
sparse 3-dimensional dynamic matrix containing the partition function of
alignments starting in state
match
and ending once all the characters
have been inserted into the alignment. 31
b
Z
X
sparse 3-dimensional dynamic matrix containing the partition function of
alignments starting in state
gapX
and ending once all the characters
have been inserted into the alignment. 31
b
Z
Y
sparse 3-dimensional dynamic matrix containing the partition function of
alignments starting in state
gapY
and ending once all the characters
have been inserted into the alignment. 31
e
P
set of overlap-free paths. See Appendix
F
for a description of the algorithm
that resolve the block overlap for a path
p
∈ P
. 87, 160
ξ
(
v
) it represents the non-survival probability associated to the node
v
ob-tained as
ξ
(
v
) = 1
−
ζ
(
v
) which corresponds therefore at the
complemen-tary probability to the survival probability
ζ
. 4
ζ
(
v
) ‘pure’ survival probability associated to the node
v
given an insertion,
on a random location, on the path (pa
→
Ω). Differently from the
function
β
(
v
) the character is already present at pa(
v
) whereas in
β
(
v
)
the character is inserted on a random location along the edge belonging
to
v
. 3, 4
List of Symbols
xxxiii
b
(
v
) branch length associated to node
v
, which is the length of the edge from
pa(
v
) to
v
. 123
c
∅
MSA column containing only gap characters. Such columns are not
observ-able in an MSA and the number thereof is unknown. xxxiii, 9, 13–15,
18, 21, 39, 51, 53, 115, 123
e
it denotes an edge with
e
∈ E
between
v
and pa(
v
) with
b
(
e
) =
b
(
v
). 3, 120,
121
f
v
felsenstein’s weight. It corresponds to the likelihood under the given
topol-ogy of a given MSA column accounting for the process of substitution
and deletion. See Appendix
A.3
for more details. 123
n
G
number of columns containing gaps (but not full of gaps), corresponds to
n
G
=
λ
· k
τ
k
+
λ
µ
h
1
−
exp(
−k
τ
k
µ
)
i
. See Appendix
C
for more details.
141
n
H
number of columns without gaps, where
n
H
=
λ
µ
exp(
−k
τ
k
µ
). See
Ap-pendix
C
for more details. 141
p
τ
(
m
) marginal likelihood function under PIP of an alignment
m
given a
phy-logenetic tree
τ
, marginalized over ancestral states (see the original paper
for more details [
13
]). 115
p
v
(
c
∅
) likelihood of an MSA column full of gaps computed rooting the topology
at node
v
. When omitted,
v
= Ω. 9
List of Acronyms
ASRV
Across Sites Rate Variation. lii, 11, 49
CDF
cumulative distribution function. 146
CTMC
Continuous-Time Markov Chains. 125, 149
DP
Dynamic Programming. xlvii, 11, 23, 29, 33, 46, 56
FFT
Fast Fourier Transform. 56, 57, 64, 68, 151
indel
stands for insertions and/or deletions. xli, 140
ML
Maximum Likelihood. 56
MSA
Multiple Sequence Alignment. xliv, xlvi, 29
probability distribution function. 51, 121, 147
PIP
Poisson Indel Process (see [
13
]). 31, 146
SB
Stochastic Backtracking. 30, 31, 34, 36–39, 44–46
SP
sum-of-pairs. xlvi, xlvii, 165
Introduction
“We are at the very beginning of time for the human race. It is not
unreasonable that we grapple with problems. But there are tens of
thousands of years in the future. Our responsibility is to do what we
can, learn what we can, improve the solutions, and pass them on.”
– Richard P. Feynman
E
volution
is the genetic code alteration of a population over
time mainly driven by natural selection. The modern theory
of evolution is supported with evidences by many scientific
do-mains, not only from biological sciences but also, to mention
a few, from geology, anthropology, chemistry, physics,
math-ematics, astrophysics, psychology as well as behavioral and
social sciences. In 1859,
Charles Darwin
published the seminal book
On the
Origin of Species
where he elaborated the scientific theory of evolution by
nat-ural selection. Since then, many other discoveries in the life sciences endorsed
or extended his hypothesis, that is, all organisms are to some extent related,
species evolve as response to natural selection. To quote
Theodosius
Dobzhan-sky
, a prominent Ukrainian-American geneticist and evolutionary biologist,
“
Nothing in biology makes sense except in the light of evolution
”.
Nonetheless, natural selection is not the single evolutionary driving force for
the biodiversity, variations occur for example also via mutations, sexual
se-lection, genetic drift, and artificial selection. However, selection can occur
only if traits exhibits some genetic variability within a population. Although
evolution happens at the level of population rather than at the single
individ-ual level, heritable changes take place in individindivid-uals and those bringing any
phenotypic advantage are more likely to be passed at the next generations.
Therefore, the frequency of advantageous alleles increases with the time while
that of disadvantageous ones tends to diminish. These individual phenotypic
changes within a population are modifying the distribution of phenotypic
val-ues over time. When the phenotypic shift brings some benefit to the fitness,
than it is likely that this modification is conserved over generations.
xxxviii
Introduction
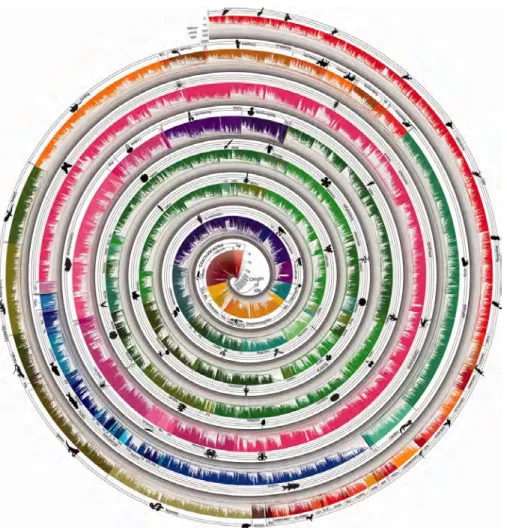
Figure 1: Tree of life reveals clock-like speciation and diversification. A timetree
of 50,632 species synthesized from times of divergence published in 2,274 studies.
Evolutionary history is compressed into a narrow strip and then arranged in a spiral
with one end in the middle and the other on the outside. Therefore, time progresses
across the width of the strip at all places, rather than along the spiral. Time is shown
in billions of years on a log scale and indicated throughout by bands of gray. Major
taxonomic groups are labeled and the different color ranges correspond to the main
taxonomic divisions of our tree.
Image and caption reproduced with permission from Blair Hedges et al. [
51
].
Along with natural selection processes, also catastrophic events or
environ-mental changes may induce changes in the gene pools of a population. This
population gene diversity is actually crucial to survive within a dynamically
changing environment. A wide assortment of genes increases the probability
that some individuals possess the differentiated allele needed to better adapt
Introduction
xxxix
to the environment. In small population sizes the genetic variability in general
decreases as the inbreeding or the mating between individuals having similar
genetic makeup is more likely to occur. Less biodiversity implies, in turn, less
chances of coping with environmental variability.
Genetic variation increases with the spontaneous generation of new alleles
but also because of gene flowing (horizontal/lateral gene transfer) from other
individuals which introduce in this way new traits from foreign populations.
These phenomena along with natural selection, establish part of the natural
process yielding towards evolution and speciation. Indeed, life continues to
evolve through speciations and extinctions, two key mechanisms controlling
biodiversity. These events are depicted graphically be means of phylogenetic
trees. An example is provided in Figure
1
.
However, speciation and extinction are events happening at different rates.
Speciation and extinction rates establish the frequency at which new species
originate and are lost over evolutionary time. Typically, species that promote
speciation are also highly specialized to a particular environment, are often
isolated or have a low population size. The same characteristics are associated
also with extinction. Actually, specialized species are susceptible to
environ-mental change and therefore are predisposed to disappear rapidly. Speciation
can occur slowly and gradually often described by point mutations or appear
with silent periods interrupted by event “bursts”. As a result, when two
pop-ulations that have diverged from a common ancestor become reproductively
distincts, i.e. capable of interbreeding and exchanging genetic information to
produce fertile offspring, a new species arise.
The carriers of the genetic information are DNA and RNA, they constitute
the building elements of any genetic code and of different pathways. These
“building blocks” are conserved across all the species while the encoded
infor-mation may changes. Observing the conserved core processes and the essential
features shared among all organisms, both extinct or extant, suggests that all
organisms descent from a common ancestry (see Figure
1
). In fact, some of
these mechanisms are so sophisticated that the probability that they emerged
independently several times is almost negligible. All living beings endowing
these features, and from which diversification occurred, led to the development
of the three domains: Archaea, Bacteria, and Eucarya. The universal common
ancestor had as its genetic material the DNA which, through transcription and
translation, expressed its genetic code. Phylogenetic trees show graphically the
evolutionary history of acquired and lost traits during the evolution.
xl
Introduction
substitution
<latexit sha1_base64="EHTcu7lVtu0tS3V33CeHFezbMlE=">AAAB/HicbVDLSgMxFM3UV62v0S7dBIvgqsyIoMuiG5cV7APaoWTSTBuayQzJjViG+ituXCji1g9x59+YaWehrQcCh3Pu4d6cMBVcg+d9O6W19Y3NrfJ2ZWd3b//APTxq68Qoylo0EYnqhkQzwSVrAQfBuqliJA4F64STm9zvPDCleSLvYZqyICYjySNOCVhp4Fb7wB4h0ybUNm1ycTZwa17dmwOvEr8gNVSgOXC/+sOEmphJoIJo3fO9FIKMKOBUsFmlbzRLCZ2QEetZKknMdJDNj5/hU6sMcZQo+yTgufo7kZFY62kc2smYwFgve7n4n9czEF0FGZepASbpYlFkBIYE503gIVeMgphaQqji9lZMx0QRCravii3BX/7yKmmf133L7y5qjeuijjI6RifoDPnoEjXQLWqiFqJoip7RK3pznpwX5935WIyWnCJTRX/gfP4AL0qVvw==</latexit><latexit sha1_base64="EHTcu7lVtu0tS3V33CeHFezbMlE=">AAAB/HicbVDLSgMxFM3UV62v0S7dBIvgqsyIoMuiG5cV7APaoWTSTBuayQzJjViG+ituXCji1g9x59+YaWehrQcCh3Pu4d6cMBVcg+d9O6W19Y3NrfJ2ZWd3b//APTxq68Qoylo0EYnqhkQzwSVrAQfBuqliJA4F64STm9zvPDCleSLvYZqyICYjySNOCVhp4Fb7wB4h0ybUNm1ycTZwa17dmwOvEr8gNVSgOXC/+sOEmphJoIJo3fO9FIKMKOBUsFmlbzRLCZ2QEetZKknMdJDNj5/hU6sMcZQo+yTgufo7kZFY62kc2smYwFgve7n4n9czEF0FGZepASbpYlFkBIYE503gIVeMgphaQqji9lZMx0QRCravii3BX/7yKmmf133L7y5qjeuijjI6RifoDPnoEjXQLWqiFqJoip7RK3pznpwX5935WIyWnCJTRX/gfP4AL0qVvw==</latexit><latexit sha1_base64="EHTcu7lVtu0tS3V33CeHFezbMlE=">AAAB/HicbVDLSgMxFM3UV62v0S7dBIvgqsyIoMuiG5cV7APaoWTSTBuayQzJjViG+ituXCji1g9x59+YaWehrQcCh3Pu4d6cMBVcg+d9O6W19Y3NrfJ2ZWd3b//APTxq68Qoylo0EYnqhkQzwSVrAQfBuqliJA4F64STm9zvPDCleSLvYZqyICYjySNOCVhp4Fb7wB4h0ybUNm1ycTZwa17dmwOvEr8gNVSgOXC/+sOEmphJoIJo3fO9FIKMKOBUsFmlbzRLCZ2QEetZKknMdJDNj5/hU6sMcZQo+yTgufo7kZFY62kc2smYwFgve7n4n9czEF0FGZepASbpYlFkBIYE503gIVeMgphaQqji9lZMx0QRCravii3BX/7yKmmf133L7y5qjeuijjI6RifoDPnoEjXQLWqiFqJoip7RK3pznpwX5935WIyWnCJTRX/gfP4AL0qVvw==</latexit><latexit sha1_base64="EHTcu7lVtu0tS3V33CeHFezbMlE=">AAAB/HicbVDLSgMxFM3UV62v0S7dBIvgqsyIoMuiG5cV7APaoWTSTBuayQzJjViG+ituXCji1g9x59+YaWehrQcCh3Pu4d6cMBVcg+d9O6W19Y3NrfJ2ZWd3b//APTxq68Qoylo0EYnqhkQzwSVrAQfBuqliJA4F64STm9zvPDCleSLvYZqyICYjySNOCVhp4Fb7wB4h0ybUNm1ycTZwa17dmwOvEr8gNVSgOXC/+sOEmphJoIJo3fO9FIKMKOBUsFmlbzRLCZ2QEetZKknMdJDNj5/hU6sMcZQo+yTgufo7kZFY62kc2smYwFgve7n4n9czEF0FGZepASbpYlFkBIYE503gIVeMgphaQqji9lZMx0QRCravii3BX/7yKmmf133L7y5qjeuijjI6RifoDPnoEjXQLWqiFqJoip7RK3pznpwX5935WIyWnCJTRX/gfP4AL0qVvw==</latexit>insertion
<latexit sha1_base64="jvMxddWgcjF1afbc/JmDYHOnHSI=">AAAB+XicbVBNS8NAEN34WetX1KOXxSJ4KokIeix68VjBfkAbymY7bZduNmF3Uiyh/8SLB0W8+k+8+W/ctDlo64OFx3szszMvTKQw6Hnfztr6xubWdmmnvLu3f3DoHh03TZxqDg0ey1i3Q2ZACgUNFCihnWhgUSihFY7vcr81AW1ErB5xmkAQsaESA8EZWqnnul2EJ8yEMqBzZdZzK17Vm4OuEr8gFVKg3nO/uv2YpxEo5JIZ0/G9BIOM2XFcwqzcTQ0kjI/ZEDqWKhaBCbL55jN6bpU+HcTaPoV0rv7uyFhkzDQKbWXEcGSWvVz8z+ukOLgJ7FlJiqD44qNBKinGNI+B9oUGjnJqCeNa2F0pHzHNONqwyjYEf/nkVdK8rPqWP1xVardFHCVySs7IBfHJNamRe1InDcLJhDyTV/LmZM6L8+58LErXnKLnhPyB8/kDkGyUPw==</latexit><latexit sha1_base64="jvMxddWgcjF1afbc/JmDYHOnHSI=">AAAB+XicbVBNS8NAEN34WetX1KOXxSJ4KokIeix68VjBfkAbymY7bZduNmF3Uiyh/8SLB0W8+k+8+W/ctDlo64OFx3szszMvTKQw6Hnfztr6xubWdmmnvLu3f3DoHh03TZxqDg0ey1i3Q2ZACgUNFCihnWhgUSihFY7vcr81AW1ErB5xmkAQsaESA8EZWqnnul2EJ8yEMqBzZdZzK17Vm4OuEr8gFVKg3nO/uv2YpxEo5JIZ0/G9BIOM2XFcwqzcTQ0kjI/ZEDqWKhaBCbL55jN6bpU+HcTaPoV0rv7uyFhkzDQKbWXEcGSWvVz8z+ukOLgJ7FlJiqD44qNBKinGNI+B9oUGjnJqCeNa2F0pHzHNONqwyjYEf/nkVdK8rPqWP1xVardFHCVySs7IBfHJNamRe1InDcLJhDyTV/LmZM6L8+58LErXnKLnhPyB8/kDkGyUPw==</latexit><latexit sha1_base64="jvMxddWgcjF1afbc/JmDYHOnHSI=">AAAB+XicbVBNS8NAEN34WetX1KOXxSJ4KokIeix68VjBfkAbymY7bZduNmF3Uiyh/8SLB0W8+k+8+W/ctDlo64OFx3szszMvTKQw6Hnfztr6xubWdmmnvLu3f3DoHh03TZxqDg0ey1i3Q2ZACgUNFCihnWhgUSihFY7vcr81AW1ErB5xmkAQsaESA8EZWqnnul2EJ8yEMqBzZdZzK17Vm4OuEr8gFVKg3nO/uv2YpxEo5JIZ0/G9BIOM2XFcwqzcTQ0kjI/ZEDqWKhaBCbL55jN6bpU+HcTaPoV0rv7uyFhkzDQKbWXEcGSWvVz8z+ukOLgJ7FlJiqD44qNBKinGNI+B9oUGjnJqCeNa2F0pHzHNONqwyjYEf/nkVdK8rPqWP1xVardFHCVySs7IBfHJNamRe1InDcLJhDyTV/LmZM6L8+58LErXnKLnhPyB8/kDkGyUPw==</latexit><latexit sha1_base64="jvMxddWgcjF1afbc/JmDYHOnHSI=">AAAB+XicbVBNS8NAEN34WetX1KOXxSJ4KokIeix68VjBfkAbymY7bZduNmF3Uiyh/8SLB0W8+k+8+W/ctDlo64OFx3szszMvTKQw6Hnfztr6xubWdmmnvLu3f3DoHh03TZxqDg0ey1i3Q2ZACgUNFCihnWhgUSihFY7vcr81AW1ErB5xmkAQsaESA8EZWqnnul2EJ8yEMqBzZdZzK17Vm4OuEr8gFVKg3nO/uv2YpxEo5JIZ0/G9BIOM2XFcwqzcTQ0kjI/ZEDqWKhaBCbL55jN6bpU+HcTaPoV0rv7uyFhkzDQKbWXEcGSWvVz8z+ukOLgJ7FlJiqD44qNBKinGNI+B9oUGjnJqCeNa2F0pHzHNONqwyjYEf/nkVdK8rPqWP1xVardFHCVySs7IBfHJNamRe1InDcLJhDyTV/LmZM6L8+58LErXnKLnhPyB8/kDkGyUPw==</latexit>deletion
<latexit sha1_base64="9yC9uzlpvYuv/8HbW8KRT2w66S8=">AAAB+HicbZBNS8NAEIY39avWj1Y9egkWwVNJRNBj0YvHCvYD2lA2m0m7dLMJuxOxhv4SLx4U8epP8ea/cdPmoK0vLDy8M8PMvn4iuEbH+bZKa+sbm1vl7crO7t5+tXZw2NFxqhi0WSxi1fOpBsEltJGjgF6igEa+gK4/ucnr3QdQmsfyHqcJeBEdSR5yRtFYw1p1gPCIWQACcmM2rNWdhjOXvQpuAXVSqDWsfQ2CmKURSGSCat13nQS9jCrkTMCsMkg1JJRN6Aj6BiWNQHvZ/PCZfWqcwA5jZZ5Ee+7+nshopPU08k1nRHGsl2u5+V+tn2J45WVcJimCZItFYSpsjO08BTvgChiKqQHKFDe32mxMFWVosqqYENzlL69C57zhGr67qDevizjK5JickDPikkvSJLekRdqEkZQ8k1fyZj1ZL9a79bFoLVnFzBH5I+vzB5eMk64=</latexit><latexit sha1_base64="9yC9uzlpvYuv/8HbW8KRT2w66S8=">AAAB+HicbZBNS8NAEIY39avWj1Y9egkWwVNJRNBj0YvHCvYD2lA2m0m7dLMJuxOxhv4SLx4U8epP8ea/cdPmoK0vLDy8M8PMvn4iuEbH+bZKa+sbm1vl7crO7t5+tXZw2NFxqhi0WSxi1fOpBsEltJGjgF6igEa+gK4/ucnr3QdQmsfyHqcJeBEdSR5yRtFYw1p1gPCIWQACcmM2rNWdhjOXvQpuAXVSqDWsfQ2CmKURSGSCat13nQS9jCrkTMCsMkg1JJRN6Aj6BiWNQHvZ/PCZfWqcwA5jZZ5Ee+7+nshopPU08k1nRHGsl2u5+V+tn2J45WVcJimCZItFYSpsjO08BTvgChiKqQHKFDe32mxMFWVosqqYENzlL69C57zhGr67qDevizjK5JickDPikkvSJLekRdqEkZQ8k1fyZj1ZL9a79bFoLVnFzBH5I+vzB5eMk64=</latexit><latexit sha1_base64="9yC9uzlpvYuv/8HbW8KRT2w66S8=">AAAB+HicbZBNS8NAEIY39avWj1Y9egkWwVNJRNBj0YvHCvYD2lA2m0m7dLMJuxOxhv4SLx4U8epP8ea/cdPmoK0vLDy8M8PMvn4iuEbH+bZKa+sbm1vl7crO7t5+tXZw2NFxqhi0WSxi1fOpBsEltJGjgF6igEa+gK4/ucnr3QdQmsfyHqcJeBEdSR5yRtFYw1p1gPCIWQACcmM2rNWdhjOXvQpuAXVSqDWsfQ2CmKURSGSCat13nQS9jCrkTMCsMkg1JJRN6Aj6BiWNQHvZ/PCZfWqcwA5jZZ5Ee+7+nshopPU08k1nRHGsl2u5+V+tn2J45WVcJimCZItFYSpsjO08BTvgChiKqQHKFDe32mxMFWVosqqYENzlL69C57zhGr67qDevizjK5JickDPikkvSJLekRdqEkZQ8k1fyZj1ZL9a79bFoLVnFzBH5I+vzB5eMk64=</latexit><latexit sha1_base64="9yC9uzlpvYuv/8HbW8KRT2w66S8=">AAAB+HicbZBNS8NAEIY39avWj1Y9egkWwVNJRNBj0YvHCvYD2lA2m0m7dLMJuxOxhv4SLx4U8epP8ea/cdPmoK0vLDy8M8PMvn4iuEbH+bZKa+sbm1vl7crO7t5+tXZw2NFxqhi0WSxi1fOpBsEltJGjgF6igEa+gK4/ucnr3QdQmsfyHqcJeBEdSR5yRtFYw1p1gPCIWQACcmM2rNWdhjOXvQpuAXVSqDWsfQ2CmKURSGSCat13nQS9jCrkTMCsMkg1JJRN6Aj6BiWNQHvZ/PCZfWqcwA5jZZ5Ee+7+nshopPU08k1nRHGsl2u5+V+tn2J45WVcJimCZItFYSpsjO08BTvgChiKqQHKFDe32mxMFWVosqqYENzlL69C57zhGr67qDevizjK5JickDPikkvSJLekRdqEkZQ8k1fyZj1ZL9a79bFoLVnFzBH5I+vzB5eMk64=</latexit>inversion
<latexit sha1_base64="cW1WjwCPyOJBY69CV/SqgjQrmD0=">AAAB+XicbVDLSsNAFJ34rPUVdelmsAiuSiKCLotuXFawD2hDmUxv2qGTSZi5KZbQP3HjQhG3/ok7/8Zpm4W2Hhg4nHMuc+8JUykMet63s7a+sbm1Xdop7+7tHxy6R8dNk2SaQ4MnMtHtkBmQQkEDBUpopxpYHEpohaO7md8agzYiUY84SSGI2UCJSHCGVuq5bhfhCXOhitC051a8qjcHXSV+QSqkQL3nfnX7Cc9iUMglM6bjeykGOdMouIRpuZsZSBkfsQF0LFUsBhPk882n9NwqfRol2j6FdK7+nshZbMwkDm0yZjg0y95M/M/rZBjdBPasNENQfPFRlEmKCZ3VQPtCA0c5sYRxLeyulA+ZZhxtD2Vbgr988ippXlZ9yx+uKrXboo4SOSVn5IL45JrUyD2pkwbhZEyeySt5c3LnxXl3PhbRNaeYOSF/4Hz+AJOFlEE=</latexit><latexit sha1_base64="cW1WjwCPyOJBY69CV/SqgjQrmD0=">AAAB+XicbVDLSsNAFJ34rPUVdelmsAiuSiKCLotuXFawD2hDmUxv2qGTSZi5KZbQP3HjQhG3/ok7/8Zpm4W2Hhg4nHMuc+8JUykMet63s7a+sbm1Xdop7+7tHxy6R8dNk2SaQ4MnMtHtkBmQQkEDBUpopxpYHEpohaO7md8agzYiUY84SSGI2UCJSHCGVuq5bhfhCXOhitC051a8qjcHXSV+QSqkQL3nfnX7Cc9iUMglM6bjeykGOdMouIRpuZsZSBkfsQF0LFUsBhPk882n9NwqfRol2j6FdK7+nshZbMwkDm0yZjg0y95M/M/rZBjdBPasNENQfPFRlEmKCZ3VQPtCA0c5sYRxLeyulA+ZZhxtD2Vbgr988ippXlZ9yx+uKrXboo4SOSVn5IL45JrUyD2pkwbhZEyeySt5c3LnxXl3PhbRNaeYOSF/4Hz+AJOFlEE=</latexit><latexit sha1_base64="cW1WjwCPyOJBY69CV/SqgjQrmD0=">AAAB+XicbVDLSsNAFJ34rPUVdelmsAiuSiKCLotuXFawD2hDmUxv2qGTSZi5KZbQP3HjQhG3/ok7/8Zpm4W2Hhg4nHMuc+8JUykMet63s7a+sbm1Xdop7+7tHxy6R8dNk2SaQ4MnMtHtkBmQQkEDBUpopxpYHEpohaO7md8agzYiUY84SSGI2UCJSHCGVuq5bhfhCXOhitC051a8qjcHXSV+QSqkQL3nfnX7Cc9iUMglM6bjeykGOdMouIRpuZsZSBkfsQF0LFUsBhPk882n9NwqfRol2j6FdK7+nshZbMwkDm0yZjg0y95M/M/rZBjdBPasNENQfPFRlEmKCZ3VQPtCA0c5sYRxLeyulA+ZZhxtD2Vbgr988ippXlZ9yx+uKrXboo4SOSVn5IL45JrUyD2pkwbhZEyeySt5c3LnxXl3PhbRNaeYOSF/4Hz+AJOFlEE=</latexit><latexit sha1_base64="cW1WjwCPyOJBY69CV/SqgjQrmD0=">AAAB+XicbVDLSsNAFJ34rPUVdelmsAiuSiKCLotuXFawD2hDmUxv2qGTSZi5KZbQP3HjQhG3/ok7/8Zpm4W2Hhg4nHMuc+8JUykMet63s7a+sbm1Xdop7+7tHxy6R8dNk2SaQ4MnMtHtkBmQQkEDBUpopxpYHEpohaO7md8agzYiUY84SSGI2UCJSHCGVuq5bhfhCXOhitC051a8qjcHXSV+QSqkQL3nfnX7Cc9iUMglM6bjeykGOdMouIRpuZsZSBkfsQF0LFUsBhPk882n9NwqfRol2j6FdK7+nshZbMwkDm0yZjg0y95M/M/rZBjdBPasNENQfPFRlEmKCZ3VQPtCA0c5sYRxLeyulA+ZZhxtD2Vbgr988ippXlZ9yx+uKrXboo4SOSVn5IL45JrUyD2pkwbhZEyeySt5c3LnxXl3PhbRNaeYOSF/4Hz+AJOFlEE=</latexit>duplication
<latexit sha1_base64="Uu6xFHee5Tqju7gt4TJ6azdplhk=">AAAB+3icbZBNS8NAEIY39avWr1iPXoJF8FQSEfRY9OKxgv2ANpTNZtIu3WzC7kRaQv+KFw+KePWPePPfuG1z0NYXFh7emWFm3yAVXKPrfluljc2t7Z3ybmVv/+DwyD6utnWSKQYtlohEdQOqQXAJLeQooJsqoHEgoBOM7+b1zhMozRP5iNMU/JgOJY84o2isgV3tI0wwDzOzbOnNBnbNrbsLOevgFVAjhZoD+6sfJiyLQSITVOue56bo51QhZwJmlX6mIaVsTIfQMyhpDNrPF7fPnHPjhE6UKPMkOgv390ROY62ncWA6Y4ojvVqbm//VehlGN37OZZohSLZcFGXCwcSZB+GEXAFDMTVAmeLmVoeNqKIMTVwVE4K3+uV1aF/WPcMPV7XGbRFHmZySM3JBPHJNGuSeNEmLMDIhz+SVvFkz68V6tz6WrSWrmDkhf2R9/gAM8JUU</latexit><latexit sha1_base64="Uu6xFHee5Tqju7gt4TJ6azdplhk=">AAAB+3icbZBNS8NAEIY39avWr1iPXoJF8FQSEfRY9OKxgv2ANpTNZtIu3WzC7kRaQv+KFw+KePWPePPfuG1z0NYXFh7emWFm3yAVXKPrfluljc2t7Z3ybmVv/+DwyD6utnWSKQYtlohEdQOqQXAJLeQooJsqoHEgoBOM7+b1zhMozRP5iNMU/JgOJY84o2isgV3tI0wwDzOzbOnNBnbNrbsLOevgFVAjhZoD+6sfJiyLQSITVOue56bo51QhZwJmlX6mIaVsTIfQMyhpDNrPF7fPnHPjhE6UKPMkOgv390ROY62ncWA6Y4ojvVqbm//VehlGN37OZZohSLZcFGXCwcSZB+GEXAFDMTVAmeLmVoeNqKIMTVwVE4K3+uV1aF/WPcMPV7XGbRFHmZySM3JBPHJNGuSeNEmLMDIhz+SVvFkz68V6tz6WrSWrmDkhf2R9/gAM8JUU</latexit><latexit sha1_base64="Uu6xFHee5Tqju7gt4TJ6azdplhk=">AAAB+3icbZBNS8NAEIY39avWr1iPXoJF8FQSEfRY9OKxgv2ANpTNZtIu3WzC7kRaQv+KFw+KePWPePPfuG1z0NYXFh7emWFm3yAVXKPrfluljc2t7Z3ybmVv/+DwyD6utnWSKQYtlohEdQOqQXAJLeQooJsqoHEgoBOM7+b1zhMozRP5iNMU/JgOJY84o2isgV3tI0wwDzOzbOnNBnbNrbsLOevgFVAjhZoD+6sfJiyLQSITVOue56bo51QhZwJmlX6mIaVsTIfQMyhpDNrPF7fPnHPjhE6UKPMkOgv390ROY62ncWA6Y4ojvVqbm//VehlGN37OZZohSLZcFGXCwcSZB+GEXAFDMTVAmeLmVoeNqKIMTVwVE4K3+uV1aF/WPcMPV7XGbRFHmZySM3JBPHJNGuSeNEmLMDIhz+SVvFkz68V6tz6WrSWrmDkhf2R9/gAM8JUU</latexit><latexit sha1_base64="Uu6xFHee5Tqju7gt4TJ6azdplhk=">AAAB+3icbZBNS8NAEIY39avWr1iPXoJF8FQSEfRY9OKxgv2ANpTNZtIu3WzC7kRaQv+KFw+KePWPePPfuG1z0NYXFh7emWFm3yAVXKPrfluljc2t7Z3ybmVv/+DwyD6utnWSKQYtlohEdQOqQXAJLeQooJsqoHEgoBOM7+b1zhMozRP5iNMU/JgOJY84o2isgV3tI0wwDzOzbOnNBnbNrbsLOevgFVAjhZoD+6sfJiyLQSITVOue56bo51QhZwJmlX6mIaVsTIfQMyhpDNrPF7fPnHPjhE6UKPMkOgv390ROY62ncWA6Y4ojvVqbm//VehlGN37OZZohSLZcFGXCwcSZB+GEXAFDMTVAmeLmVoeNqKIMTVwVE4K3+uV1aF/WPcMPV7XGbRFHmZySM3JBPHJNGuSeNEmLMDIhz+SVvFkz68V6tz6WrSWrmDkhf2R9/gAM8JUU</latexit>
translocation
<latexit sha1_base64="9evadiwIcdnOyNAgZCS2RLsrTGA=">AAAB/XicbZBNS8NAEIYn9avWr/hx87JYBE8lEUGPRS8eK9hWaEPZbLft0s0m7E7EGop/xYsHRbz6P7z5b9y2OWjrCwsP78wws2+YSGHQ876dwtLyyupacb20sbm1vePu7jVMnGrG6yyWsb4LqeFSKF5HgZLfJZrTKJS8GQ6vJvXmPddGxOoWRwkPItpXoicYRWt13IM28gfMUFNlZDxzxx237FW8qcgi+DmUIVet4361uzFLI66QSWpMy/cSDDKqUTDJx6V2anhC2ZD2ecuiohE3QTa9fkyOrdMlvVjbp5BM3d8TGY2MGUWh7YwoDsx8bWL+V2ul2LsIMqGSFLlis0W9VBKMySQK0hWaM5QjC5RpYW8lbEA1ZWgDK9kQ/PkvL0LjtOJbvjkrVy/zOIpwCEdwAj6cQxWuoQZ1YPAIz/AKb86T8+K8Ox+z1oKTz+zDHzmfP8Kxlg0=</latexit><latexit sha1_base64="9evadiwIcdnOyNAgZCS2RLsrTGA=">AAAB/XicbZBNS8NAEIYn9avWr/hx87JYBE8lEUGPRS8eK9hWaEPZbLft0s0m7E7EGop/xYsHRbz6P7z5b9y2OWjrCwsP78wws2+YSGHQ876dwtLyyupacb20sbm1vePu7jVMnGrG6yyWsb4LqeFSKF5HgZLfJZrTKJS8GQ6vJvXmPddGxOoWRwkPItpXoicYRWt13IM28gfMUFNlZDxzxx237FW8qcgi+DmUIVet4361uzFLI66QSWpMy/cSDDKqUTDJx6V2anhC2ZD2ecuiohE3QTa9fkyOrdMlvVjbp5BM3d8TGY2MGUWh7YwoDsx8bWL+V2ul2LsIMqGSFLlis0W9VBKMySQK0hWaM5QjC5RpYW8lbEA1ZWgDK9kQ/PkvL0LjtOJbvjkrVy/zOIpwCEdwAj6cQxWuoQZ1YPAIz/AKb86T8+K8Ox+z1oKTz+zDHzmfP8Kxlg0=</latexit><latexit sha1_base64="9evadiwIcdnOyNAgZCS2RLsrTGA=">AAAB/XicbZBNS8NAEIYn9avWr/hx87JYBE8lEUGPRS8eK9hWaEPZbLft0s0m7E7EGop/xYsHRbz6P7z5b9y2OWjrCwsP78wws2+YSGHQ876dwtLyyupacb20sbm1vePu7jVMnGrG6yyWsb4LqeFSKF5HgZLfJZrTKJS8GQ6vJvXmPddGxOoWRwkPItpXoicYRWt13IM28gfMUFNlZDxzxx237FW8qcgi+DmUIVet4361uzFLI66QSWpMy/cSDDKqUTDJx6V2anhC2ZD2ecuiohE3QTa9fkyOrdMlvVjbp5BM3d8TGY2MGUWh7YwoDsx8bWL+V2ul2LsIMqGSFLlis0W9VBKMySQK0hWaM5QjC5RpYW8lbEA1ZWgDK9kQ/PkvL0LjtOJbvjkrVy/zOIpwCEdwAj6cQxWuoQZ1YPAIz/AKb86T8+K8Ox+z1oKTz+zDHzmfP8Kxlg0=</latexit><latexit sha1_base64="9evadiwIcdnOyNAgZCS2RLsrTGA=">AAAB/XicbZBNS8NAEIYn9avWr/hx87JYBE8lEUGPRS8eK9hWaEPZbLft0s0m7E7EGop/xYsHRbz6P7z5b9y2OWjrCwsP78wws2+YSGHQ876dwtLyyupacb20sbm1vePu7jVMnGrG6yyWsb4LqeFSKF5HgZLfJZrTKJS8GQ6vJvXmPddGxOoWRwkPItpXoicYRWt13IM28gfMUFNlZDxzxx237FW8qcgi+DmUIVet4361uzFLI66QSWpMy/cSDDKqUTDJx6V2anhC2ZD2ecuiohE3QTa9fkyOrdMlvVjbp5BM3d8TGY2MGUWh7YwoDsx8bWL+V2ul2LsIMqGSFLlis0W9VBKMySQK0hWaM5QjC5RpYW8lbEA1ZWgDK9kQ/PkvL0LjtOJbvjkrVy/zOIpwCEdwAj6cQxWuoQZ1YPAIz/AKb86T8+K8Ox+z1oKTz+zDHzmfP8Kxlg0=</latexit>
Figure 2: Genetic variations at the molecular level. The main processes leading to a
genetic variation are substitution (point mutations), insertions (of one or more
char-acters), deletions (of one or more existing charchar-acters), translocations (the change of
location of a chunk of characters), duplications (the copying of a block of characters
one or many times, notably tandem repeats when the copies are placed at the
imme-diate adjacent locations) and inversions (the rearrangement in which a segment of
is reversed end to end).
At the molecular level the main processes leading to a genetic variation are
substitution
(point mutations),
insertions
(of one or more characters),
deletions
(of one or more existing characters),
translocations
(the change of location of
a chunk of characters),
duplications
(the copying of a block of characters one