0022-538X/96/$04.00
1
0
Copyright
q
1996, American Society for Microbiology
Progression to Persistent Lymphocytosis and Tumor Development in
Bovine Leukemia Virus (BLV)-Infected Cattle Correlates with
Impaired Proliferation of CD4
1
T Cells in Response to
gag
- and
env
-Encoded BLV Proteins
OTO ORLIK
1†
ANDGARY A. SPLITTER*
Department of Animal Health and Biomedical Sciences, University of
Wisconsin—Madison, Madison, Wisconsin 53706
Received 22 May 1996/Accepted 2 August 1996
The mechanism of leukemogenesis and persistent lymphocytosis (PL; benign expansion of B lymphocytes)
in cattle infected with bovine leukemia virus (BLV; a retrovirus closely related to human T-cell leukemia virus
type 1) is unknown; however, the immune system likely plays an important role in controlling the outcome of
infection. In this study, we compared T-cell competence in serologically positive alymphocytotic (AL) animals
with T-cell functions in animals with progressive stages of infection, PL and tumor bearing (TB). Dramatic
differences were observed in lymphocyte proliferation to recombinant proteins encoded by BLV
gag
(p12, p15,
and p24) and
env
(gp30, and gp51) genes in different disease stages. Lymphocytes from AL cattle recognized
an average of three of five recombinant proteins per animal. Expansion of antigen pulsed lymphocytes in
interleukin-2 increased protein recognition to almost five per animal. In contrast, lymphocytes from PL and TB
animals failed to recognize any BLV recombinant proteins. Short-term T-cell cultures from the PL group
expanded in interleukin-2, as well as the PL and TB cells cultured in indomethacin (3 to 6
m
g/ml), increased
the average of recognized proteins per animal to one. Cells proliferating to BLV antigens were CD4
1T
lymphocytes, as shown by cell depletion studies. The positive effect of indomethacin suggests involvement of
prostaglandin E
2as a negative regulatory factor in the later stages of disease. Thus, for the first time,
advancing stages of BLV infection were correlated with decreased T-cell competence, providing deeper insight
into pathogenesis of retroviral infections.
Bovine leukemia virus (BLV), a lymphotropic retrovirus
structurally related to human T-cell leukemia virus type 1
(HTLV-1) (46), leads to development of enzootic
lymphosar-coma-leukemia in cattle (48). The low percentage (0.6 to 5%)
of infected animals developing tumors (11, 53) and long
la-tency (over 4 years) suggest that malignant transformation
involves mechanisms in addition to viral infection. A benign
consequence of BLV infection is persistent lymphocytosis
(PL), a disorder characterized by an increase in circulating B
lymphocytes (26, 35). Resistance or sensitivity to development
of PL was shown to be determined by genetic background (11)
and linked to the polymorphism in the BoLA-DRB3 allele
(55). The relationship between lymphosarcoma and PL is
un-clear, and lymphomas can occur in animals whether they
de-velop PL or not (1, 5, 11). However, the vast majority of
BLV-infected animals remain clinically healthy, and
serocon-version in AL animals (4) together with increased lymphocyte
count in PL animals are the only indications of infection.
The immune response, both humoral and cellular, plays a
role in protection from experimental BLV infection. Passive
immunization with anti-BLV antibodies conferred protection
from BLV infection (28). Resistance to BLV challenge after
vaccination of sheep with gp51 antigen (39) and cattle with
cells producing BLV
env
-encoded glycoproteins and p24 (2)
correlated with the presence of virus-specific antibodies. In
contrast, cell-mediated immunity was the major protective
mechanism in sheep inoculated with vaccinia virus expressing
gp51 (14, 37, 38, 45), and two CD8
1CTL epitopes on gp51
were identified (13). However, evaluation of cellular immunity
in naturally infected animals in different disease stages has not
been reported. Absence of these studies may be partially due
to extreme difficulties in assessing CD4
1T-cell function in
BLV-infected animals. Lymphocytes from BLV-infected
ani-mals, especially those in the PL stage, spontaneously
prolifer-ate and hinder antigen-specific responses. Previously, CD4
1T-cell epitopes were identified on gp51 by using lymphocytes
from BLV-negative cattle and sheep (13) and vaccinia
virus-vaccinated sheep (14); however, lymphocytes from
BLV-in-fected animals failed to produce specific responses.
Neverthe-less, B-cell depletion helped reduce background proliferation,
and several CD4
1determinants on gp51 (7) and p24 (31)
recognized by lymphocytes from infected cattle were identified.
Unfortunately, animals in the PL stage still could not be
eval-uated because of high spontaneous proliferation. Attempts to
evaluate T-cell function in animals with lymphosarcoma have
not been reported.
Structural proteins forming BLV virions are encoded by
gag
and
env
genes (15). The
gag
gene of BLV is translated as the
precursor Pr70 Gag and is processed into three mature proteins:
matrix protein p15, the most abundant capsid protein, p24, and
nucleocapsid protein p12. The product of the
env
gene is the
precursor Pr72 Env, which is processed into surface envelope
glycoprotein gp51 and transmembrane glycoprotein gp30.
The goal of this study was to determine and compare CD4
1T-cell competence in BLV-infected cattle in different disease
* Corresponding author. Mailing address: Department of Animal
Health and Biomedical Sciences, University of Wisconsin—Madison,
1655 Linden Dr., Madison, WI 53706. Phone: (608) 262-1837. Fax:
(608) 262-7420. Electronic mail address: [email protected].
† Permanent address: Cancer Research Institute, Slovak Academy
of Sciences, 812 32 Bratislava, Slovak Republic.
7584
on November 9, 2019 by guest
http://jvi.asm.org/
stages, i.e., alymphocytotic (AL), PL, and tumor bearing (TB).
A complete set of five BLV structural proteins was tested for
antigen-specific proliferation. Thus, a detailed analysis of the
contribution of viral proteins, lymphocyte response, and
dis-ease state of the animal can be defined in the natural host.
MATERIALS AND METHODS
Experimental animals.Fifteen cows comprising four AL (4, 17, 201, and 234), three PL (2, 182, and 612), according to the criterion of a 95% prediction limit for lymphocyte counts (52), and four TB (74, T2, 416, and 5184) animals natu-rally infected with BLV were evaluated. PL and TB animals characterized by leukocyte count/lymphocyte count/CD41T cells were as follows: animal 2, 12.5/ 10.5/1.373109/liter; animal 182, 11.1/9.3/1.113109/liter; animal 612, 16.7/15/ 1.833109/liter; animal 416, 15.1/6.1/0.813109/liter; animal 5184, 7.4/2.6/0.333 109/liter; animal T2, 77.7/48.1/3109/liter/not determined; and animal 74, not determined. TB animals had macroscopically enlarged prescapular and/or prefemoral lymph nodes. All animals were BLV seropositive as determined by agar gel immunodiffusion test. Two seronegative animals (3883 and 3993) and two animals (GS1 and GS2) immunized three times with inactivated BLV (BLVIm) were used as negative and positive controls, respectively.
Cells, virus and MAbs.BL3 (ATCC CRL 8037) and FLK BLV-infected cell lines maintained in RPMI 1640 supplemented with 2 mML-glutamine, 100 U of penicillin per ml, 100mg of streptomycin per ml, and 5% fetal serum (FS) were used to prepare BLV-containing supernatants. BLV from culture supernatant was isolated by ultracentrifugation and/or by sucrose gradient centrifugation. Virus production by BL3 cells was confirmed by flow cytometry analysis with anti-gp51 monoclonal antibodies (MAbs) (40), and the presence of viral proteins p24 and gp51 in BL3 supernatant was determined by Western blotting (immu-noblotting) and antigen capture assay with MAbs (40, 41). Before use, viral particles were disrupted by repeated cycles of freezing and thawing. MAb IL-A55 from cluster BoWC5 as well as anti-immunoglobulin M MAb 33 were used for B-cell identification. MAbs CC8 and IL-A11 with CD41specificity, MAbs CC63 and IL-A51 with CD81specificity, and anti-CD21MAb IL-A42 were used for T-cell subset characterization (23). Maltose-binding protein (MBP)-specific MAb 1E11 was prepared in our laboratory.
PCR.BLVgagandenvsequences were amplified by PCR. Plasmid pBLV913 was used to amplify p12, p15, p24, gp30, and gp51 genes. Furthermore, p12 and gp30 genes were amplified from genomic DNA, by using BLV-producing cell lines BL3 and FLK after extraction with TRI Reagent as recommended by the manufacturer (MRC, Cincinnati, Ohio). Oligo software (National Biosciences, Inc., Plymouth, Minn.) was used for primer design. The oligonucleotide se-quences, amplified fragment sizes, and nucleotide positions within the BLV genome are shown in Table 1. The nucleotides of BLV were numbered as specified by Sagata et al. (46). Each PCR mixture contained 5ml of 103 ther-mobuffer, 1.5 to 3 mM MgCl2, 0.8 mM deoxynucleoside triphosphates, 1mM each primer, 25 U ofTaqDNA polymerase (Promega, Madison, Wis.), and 0.01
mg of plasmid or 0.1mg of genomic DNA in a total of 50ml. The amplification sequence consisted of DNA denaturation at 948C for 1 min, annealing at 558C (p24 and gp51), 588C (gp30), or 618C (p12 and p15) for 1 to 2 min, and poly-merization at 728C for 1 to 1.5 min. This sequence was repeated for 35 cycles. Amplified DNA products (10 ml of reaction mixture) were evaluated by 2% agarose gel electrophoresis.
Construction of expression plasmids and purification of recombinant pro-teins. Target genes were produced as fusion proteins carrying S*Tag and a
thrombin cleavage site on the amino-terminal end, using the pET-29a expression vector (Novagen, Madison, Wis.). Purified PCR DNAs of p12, p24, and gp30 were digested withNcoI andEcoRI and ligated intoNcoI-EcoRI-digested pET-29a. Amplified p15 DNA was digested withAflIII andBamHI and ligated into NcoI-BamHI-digested pET29a, while gp51 PCR product was digested with BspHI andSacI and ligated intoNcoI-SacI-digested pET29a. Because of ineffi-cient affinity purification of pET29a expressed fusion proteins carrying S*Tag, all BLV genes were subcloned into pMAL-c2 expression vector (New England, Biolabs, Beverly, Mass.). The S*Tag portion of pET29a vector was transferred together with the BLV genes; thus, the resulting constructs contained both S*Tag (for easier detection) and MBP (for efficient affinity purification). pET-29a constructs containing p12, p15, and gp30 genes were digested withNdeI-HindIII (p12 DNA digest withNdeI was partial without cutting inside the p12 gene), and after Klenow treatment of protrudingNdeI ends, target genes were ligated into pMAL-c2 digested withBamHI-HindIII after the protrudingBamHI end was filled in with the Klenow polymerase. The p24 gene was subcloned by ligating the NspI-HindIII fragment from pET29a/p24 into NspI-HindIII-digested vector backbone from pMAL-c2/gp30. The full-length pMAL-c2/gp51 construct was obtained after ligation of theKpnI-HindIII fragment from pET29a/gp51 into the KpnI-HindIII vector backbone from pMAL-c2/gp30 DNA. Truncated gp51 (gp51/del; containing nucleotides 4913 to 5141) was constructed by blunt-end ligation of the pET29a/gp51 NdeI fragment into BamHI-digested vector pMAL-c2 following Klenow treatment. The described cloning resulted in pMAL-c2 constructs in which the MBP sequence was followed in frame by S*Tag and a particular BLV p12, p15, p24, gp30, gp51, or gp51/del gene. The resulting vectors were checked for proper orientation by restriction mapping and sequenc-ing. Ligated DNAs were transformed intoEscherichia coliDH5a(GIBCO BRL, Life Technologies, Gaithersburg, Md.), HMS174(DE3), and BL21(DE3) (No-vagen) competent cells. For protein expression, bacterial cells cultured in LB or TB medium were induced with 0.3 to 1 mM isopropylthiogalactopyranoside (IPTG) for 1 to 3 h. Bacteria were harvested by centrifugation (5,0003gfor 10 min), resuspended in phosphate-buffered saline (PBS) containing lysozyme (5 mg/ml), and lysed by five cycles of freezing and thawing. Bacterial lysates cleared by centrifugation (20,000 3gfor 1 h) were used for affinity purification of recombinant proteins by using an S*Tag purification kit (Novagen) or amylose resin (New England, Biolabs) as recommended by manufacturers. Because the fusion proteins were unstable and several degradation fragments were copuri-fied, full-length fusion proteins were recovered from sodium dodecyl sulfate (SDS)-polyacrylamide gels as described previously (19). MBP-LacZ protein was purified from the lysate of IPTG-induced DH5acells transformed with pMAL-c2. MBP was obtained after cleavage of MBP-LacZ with factor Xa (New En-gland, Biolabs).
Western blotting.Proteins were separated by SDS-polyacrylamide gel electro-phoresis (PAGE) (8 or 10% gel), and after transfer to nitrocellulose membranes, nonspecific binding sites were blocked with 5% nonfat milk in PBS for 20 min. To detect MBP-containing fusion proteins, nitrocellulose strips were incubated with anti-MBP MAb 1E11 for 1 h and then with alkaline phosphatase (AP)-labeled secondary antibody (Promega) for 1 h. Between incubations, the mem-brane strips were washed extensively with PBS containing 0.05% Tween 20. To detect recombinant proteins carrying S*Tag, nitrocellulose strips were incubated with S-protein–AP conjugate (Novagen) for 20 min. Protein bands with bound AP conjugates were visualized by nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolylphosphatep-toluidine salt.
[image:2.612.62.559.78.243.2]Lymphocyte proliferation assay. Modifications of a standard proliferation protocol using low serum (0.5 to 1%) and indomethacin (1.5 to 6mg/ml) allowed us to evaluate animals in all stages of BLV infection (42). Peripheral blood
TABLE 1. Oligonucleotide primers used for amplification of BLV genes
Amplified gene (size [bp]) and position in BLV genomea
Primer sequence (59- and 39-end primers)
p12 (262)
1565–1600 ...5
9
GGCCCCCAAGAACGAAACAGCCTGCCATGGTTGTCC
1791–1826 ...3
9
GGCGCTAAGTCCCCCCGAATTCAGTTTTTTGATTTG
p15 (394)
593–628 ...5
9
CGGCGGCATTTTGGGTCTCTCCTTACATGTATATCA
953–986 ...3
9
TGGCGGTTGCGGATCCCTTCAGATATGATTCACA
p24 (627)
941–965 ...5
9
CCCCCGCCATGGTGCCTATAATATC
1542–1567 ...3
9
GCCGAATTCTCCGCAAGCCTCAAGTT
gp30 (685)
5698–5721 ...5
9
CCGCCCTTACTTCCATGGTCAGAC
6360–6382 ...3
9
GATACGCGGGGAATTCAAGGACA
gp51 (837)
4903–4925 ...5
9
GTCGCCCGATCATGACTTGGAGA
5719–5739 ...3
9
GGGCCGCGAGAGCTCAACGTC
a
The nucleotide numbering for BLV described by Sagata et al. (46) was used.
on November 9, 2019 by guest
http://jvi.asm.org/
mononuclear cells (PBMC) were obtained by using IsoPrep as recommended by the manufacturer (Robbins Scientific, Sunnyvale, Calif.). Cells (105
per well in 96-well U-bottom plates) were resuspended in RPMI 1640 containing 0.5 to 1% FS, 531025
M 2-mercaptoethanol, and indomethacin (1.5mg/ml for AL animals and 1.5 to 6mg/ml for PL and TB animals) and cultured at 378C for 7 days with 5% CO2in the presence of antigen in a total volume of 200 ml. In some experiments, after 3 to 5 days of culture, the serum concentration was adjusted to 5 to 10% by autologous or heterologous serum. The day before harvest, [3
H]thymidine (1.0mCi/well) was added for the final 18 h, at which time the cultures were harvested onto glass filters and incorporated activity was deter-mined by a beta counter. All samples were tested at least in triplicate, and all animals were tested repeatedly over 1.5 years. The only exception was TB cow 74, which died before the initial experiments could be repeated. Lymphocyte pro-liferation to MBP and MBP-LacZ was low and used as control (background) proliferation in experiments with recombinant proteins. As in other studies (7, 13, 14), antigen-specific proliferation was considered positive if the stimulation index (SI; defined as mean counts of test sample divided by mean counts of control) was greater than 2.
Flow cytometric analysis and cell depletion.Cells (23105
) were incubated with MAb diluted 1:50 in PBS containing 1% bovine serum albumin (BSA) and 0.2% sodium azide on ice for 30 min and washed three times in cold dilution buffer. Another 30-min incubation on ice with a 1:200 dilution of a secondary dichlorotriazinyl amino fluorescein-conjugated goat anti-mouse antibody (Jack-son ImmunoResearch Laboratories Inc., West Grove, Pa.) was performed, and after three washings, cells were analyzed on an EPICS-C (Coulter Instruments, Hialeah, Fla.) apparatus. For cell depletion, PBMC (107
/ml) were incubated with MAb (IL-A55 for B-cell depletion, a mixture of IL-A11 and CC8 for CD41cell depletion, and a mixture of IL-A51 and CC63 for CD81cell depletion) diluted 1:100 in RPMI 1640 containing 1% BSA for 1 h on ice and, after washing, incubated with rabbit complement (1:10) for 1 h at 378C. Cell depletion was monitored by flow cytometric analysis on pre- and postdepletion cells. Cell viability postdepletion was determined by trypan blue exclusion.
T-cell lines.Freshly isolated PBMC (107
/ml) were incubated with 10% virus-containing BL3 supernatant for 7 to 10 days in RPMI 1640 supplemented with 1% FS and indomethacin (1.5mg/ml for AL animals and 3 to 6mg/ml for PL and TB animals). After antigen priming, cells were cultured in the presence of 2 U of human recombinant interleukin-2 (IL-2) (Boehringer Mannheim Corp., India-napolis, Ind.) per ml for an additional 10 to 20 days. After dead cells were removed by IsoPrep gradient centrifugation, T-cell cultures were restimulated by addition of antigen-presenting cells and 10% BL3 supernatant for 7 to 10 days and then incubated in IL-2-containing medium for another 10 to 20 days. This cycle was repeated one or two times. After the last expansion in IL-2, T-cell lines were cultured for 5 days in medium without IL-2 and antigen and subsequently used for proliferation assays. Adherent cells from PBMC obtained by 18-h culture were used as antigen-presenting cells.
RESULTS
Expression and purification of recombinant BLV proteins.
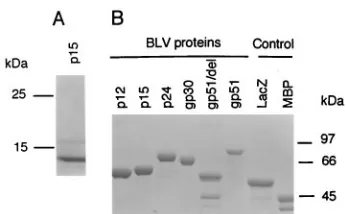
BLV structural proteins p12, p15, p24, gp30, and gp51 were
expressed by using the bacterial expression vector pET-29a as
fusion proteins carrying S*Tag. S-protein-coated Sepharose,
which selectively binds S*Tag, was used for affinity purification
and Western blot detection of recombinant proteins. Protein
p15 was successfully purified by this method, and full-length
product of 18 kDa and two major degradation products (15
and 13 kDa) were detected in Coomassie-stained
SDS-poly-acrylamide gels (Fig. 1A). Recombinant proteins p12, p24,
gp30, and gp51 did not bind the resin and could not be purified
by affinity chromatography (data not shown). Therefore, all
genes were subcloned into cloning vector pMAL-c2 and
ex-pressed as fusion proteins containing MBP (42 kDa) followed
by in-frame insertion of S*Tag and the relevant gene.
Recom-binant proteins were purified by affinity chromatography on
amylose resin columns, and proteins of the predicted
molecu-lar masses were isolated (p12 [56 kDa], p15 [59 kDa], p24 [68
kDa], gp51 [77 kDa], gp51/del [55 kDa], LacZ [51 kDa], and
MBP [42 kDa]) (Fig. 1B). The only exception was gp30, for
which the molecular mass was approximately 66 kDa instead of
the predicted 70 kDa. Therefore, the gp30 gene from
BLV-infected FLK and BL3 cells was cloned into the pMAL-c2
vector, and the molecular mass was compared with that of the
recombinant fusion gp30. Both constructs yielded proteins of
66 kDa (data not shown). These data suggested that changes in
molecular mass were due to anomalous protein migration in
SDS-PAGE rather than protein truncation. An additional
pro-tein band of 42 kDa in the gp51/del lane corresponded to
MBP, while the 38-kDa protein band in the MBP lane
corre-sponded to its major degradation product. Authenticity of
pu-rified proteins was determined by Western blot analysis (data
not shown). To separate proteins of interest from MBP-S*Tag,
factor Xa or thrombin was used. However, digestion of BLV
recombinant fusion products with factor Xa or thrombin
re-sulted in nonspecific cleavage and almost complete protein
degradation (data not shown). Therefore, the fusion proteins
purified by amylose resin were used as antigens for
prolifera-tion assays.
Lymphocyte proliferation induced by BLV proteins by AL
cattle.
In preliminary experiments, purified BLV as well as
supernatants from BLV-producing cell lines BL3 and FLK
elicited lymphocyte proliferation from seropositive animals
(data not shown). Therefore, a stock of BL3 supernatant was
prepared and used as a positive control throughout
experi-ments. Four BLV-seropositive animals (4, 17, 201, and 234)
were used to evaluate proliferative responses to BLV Gag and
Env proteins. The level of lymphocyte proliferation to BLV
and recombinant viral proteins differed among animals (Fig.
2). Lymphocytes from all AL animals proliferated after
stim-ulation with BL3 supernatant. However, the repertoire of
rec-ognized proteins varied among individuals. Animal 201
dis-played the highest SI in response to BLV and recognized all
five recombinant Gag and Env proteins. Animal 17 recognized
four recombinant proteins, while animals 4 and 234
prolifer-ated to two proteins. Control animals GS1 and GS2
immu-nized with BLV recogimmu-nized four and two proteins, respectively
(data not shown). BLV-negative animals did not proliferate
following culture with BL3 supernatant or recombinant
pro-teins. Data for one of two negative animals are shown in Fig.
2. Indomethacin treatment did not influence BLV-specific
pro-liferation in seronegative animals (data not shown).
To reveal low-frequency antigen-specific lymphocytes not
detectable by the standard 7-day assay, T-cell lines prepared as
described in Materials and Methods were tested in
prolifera-tion assays with recombinant proteins. Indeed, addiprolifera-tional viral
antigens were recognized by IL-2-treated cultures from
indi-vidual animals (Fig. 3). Animal 17 responded to all
recombi-nant proteins, animals 4 and 234 responded to two proteins,
and animals GS1 and GS2 responded to one (data not shown)
protein previously not recognized by fresh PBMC.
Interest-ingly, despite proliferation to recombinant proteins,
prolifera-tion to BLV antigen was negative in animals 4, 234 (Fig. 3), and
201 (data not shown). However, when short-term T-cell
cul-FIG. 1. Analysis of purified recombinant BLV proteins on an SDS–10% polyacrylamide gel stained with Coomassie blue. (A) Expression of BLV p15 protein from the pET29a vector. (B) Expression of BLV recombinant proteins from pMAL-c2. gp51/del, truncated gp51 containing 76 amino acid residues from the amino-terminal end of gp51; LacZ, fusion protein containingb-galactosidase
afragment and MBP.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.349.522.67.174.2]tures from animal 201 were pulsed again with BLV and
cul-tured with 2 U of recombinant IL-2 per ml, high proliferation
to BL3 supernatant (SI 26.0) was detected (data not shown).
Lymphocyte proliferation was antigen dose dependent, and
differences were observed among individuals and with different
Gag and Env proteins. Representative data for lymphocyte
proliferation in animal 17 to different concentrations of
recom-binant p24, gp30, and gp51 proteins are shown in Fig. 4.
Gen-erally, proteins at concentrations ranging from 1 to 20
m
g/ml
elicited antigen-specific proliferation, while proteins at
concen-FIG. 2. Lymphoproliferation of PBMC from a seronegative animal (3883) and AL cattle (4, 17, 201, and 234) to BLV proteins. Freshly isolated PBMC were stimulated with the indicated recombinant proteins (1 to 20mg/ml) or BLV (10% BL3 supernatant) for 7 days. gp51/tr represents the truncated form of gp51. Results are expressed as counts and represent the means of at least triplicate wells. Standard deviations are represented by vertical lines. All values above the horizontal lines representing SI 2 and marked by asterisks are considered positive (SI5mean counts in the tested sample divided by mean counts in the control MBP sample).
FIG. 3. Proliferative responses of short-term lymphocyte cultures from AL animals 4, 17, and 234 to BLV proteins. T-cell line cells (53103to 53104per well) were stimulated with the indicated recombinant proteins or 10% BL3 super-natant for 7 to 10 days. Results represent the means of at least triplicate wells. Standard deviations are represented by vertical lines. All values above the hor-izontal lines representing SI 2 and marked by asterisks are considered positive.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.328.551.66.553.2]trations higher than 50
m
g/ml were suppressive (data not
shown). In conclusion, the mean number of recognized
pro-teins per animal in the AL group, rounded to the closest
integer, was approximately 5.
Phenotypes of cells proliferating to BLV antigens.
To
deter-mine the phenotypes of cells proliferating to BLV antigens,
two sets of experiments were performed. First, PBMC from
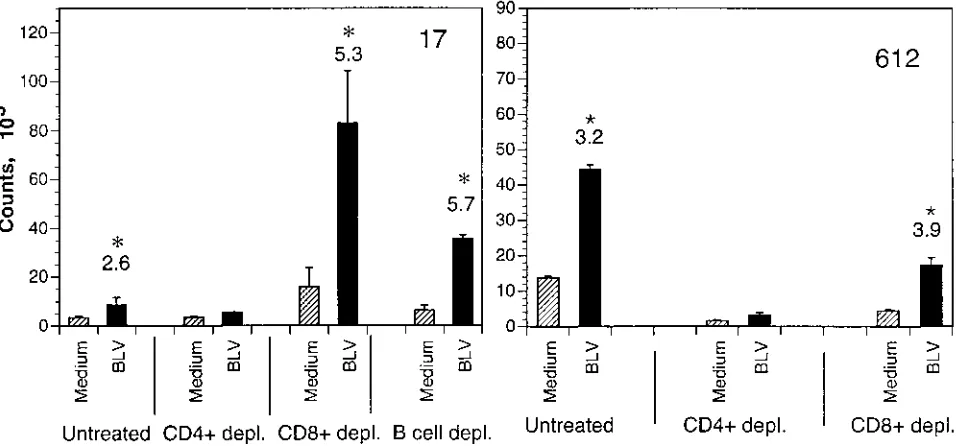
animals 17, 201, and 612 were depleted of CD4
1and CD8
1T-cell subsets or B cells by antibody and complement lysis, and
the remaining cells were cultured in the presence of antigen.
CD4
1depletion abolished or markedly reduced lymphocyte
proliferation to BLV and recombinant proteins, while CD8
1and B-cell depletion did not interfere with cell proliferation, as
shown for animals 17 and 612 in Fig. 5. Second, PBMC were
cultured in the presence of BL3 supernatant and after 10 days
evaluated by flow cytometry. The phenotypes of cells from
animal 17 before and after in vitro culture were compared
(Table 2). After culture, CD4
1cells increased nearly twofold
whereas CD8
1and B cells decreased. Thus, CD4
1T cells
represent the foremost population responsible for in vitro
pro-liferation to BLV antigens.
Response of lymphocytes from PL animals to BLV proteins.
Fresh PBMC from PL animals 2 and 182 proliferated when
stimulated with BL3 supernatant; however, PL animal 612 did
not respond to BL3 supernatant. Interestingly, in marked
con-trast to AL animals, fresh lymphocytes from all three PL
ani-mals failed to proliferate to recombinant BLV proteins (data
not shown). Previously, we found that high doses of
indometh-acin (3 to 6
m
g/ml) increased antigen-specific proliferation of
lymphocytes from PL animals (42). Therefore, cell cultures
from PL animals were supplemented with increasing doses of
indomethacin. Lymphocyte proliferation to BL3 supernatant
in animals 2 and 182 increased following addition of
indometh-acin, the highest level being found at a concentration of 6
m
g/ml. However, proliferation to all recombinant proteins
re-mained insignificant (Fig. 6A). In animal 612, addition of
in-domethacin (6
m
g/ml) elicited proliferation to BL3 supernatant
and gp30 (Fig. 6B). Next, PBMC from all three PL animals
were expanded in IL-2 (2 U/ml) and reexamined for
prolifer-ation to individual BLV proteins. IL-2-expanded PBMC from
animal 2 proliferated to three recombinant proteins (Fig. 6B).
IL-2-expanded PBMC from animals 182 and 612 did not
re-spond to any of the recombinant proteins or BLV (data not
shown). In conclusion, fresh PBMC from PL animals did not
proliferate to recombinant proteins. Treatment of cells with
low-dose IL-2 and indomethacin increased the mean number
of recognized proteins per animal to approximately 1.
Lymphoproliferation to BLV antigens in TB animals.
PBMC from four TB animals (74, T2, 416, and 5184) were
evaluated. Fresh PBMC from TB animal 74 responded to
BLV; however, proliferation to recombinant gp51 was not
ob-served with or without indomethacin, and the remaining
pro-teins were not evaluated (Fig. 7A). Fresh PBMC from TB
animal T2 responded to BL3 supernatant but not to
recombi-nant proteins. Addition of indomethacin increased
prolifera-tion to BLV (Fig. 7B) and restored proliferaprolifera-tion to
[image:5.612.97.260.69.202.2]recombi-FIG. 4. Effects of peptide concentration on lymphocyte proliferation. Lym-phocytes from animal 17 were cultured with indicated concentrations of recom-binant proteins. Results represent the means of at least triplicate wells. Standard deviations are represented by vertical lines. Values above the horizontal line representing SI 2 are considered positive.
FIG. 5. Phenotypes of cells proliferating to BLV antigens in animals 17 and 612. PBMC nondepleted or depleted of CD41or CD81T cells or B cells by complement lysis were incubated with BL3 supernatant and cultured for 7 days. Untreated cells represent PBMC treated with complement without previous MAbs treatment. Standard deviations are represented by vertical lines. Asterisks and numbers above columns indicate statistically significant SI.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.71.549.477.700.2]nant gp51 (SI 2.6 [data not shown]), while proliferation to p15
and the remaining recombinant proteins was insignificant (Fig.
7B). Fresh cells from TB animal 416 did not proliferate to BLV
or recombinant proteins. However, addition of indomethacin
(4.5 to 6
m
g/ml) elicited proliferation to BL3 supernatant (SI
4.9 [data not shown]) and p24 but not the rest of the
recom-binant proteins (Fig. 7C). Fresh PBMC from cow 5184 did not
proliferate to BL3 supernatant or recombinant proteins.
Ad-dition of indomethacin (1.5 to 7.5
m
g/ml) did not influence
BLV-induced proliferation but did restore proliferation to p24,
while proliferation to another four recombinant proteins
re-mained insignificant (Fig. 7D). All attempts to expand T cells
from TB animals in IL-2 (2 U/ml) were unsuccessful (data not
shown).
[image:6.612.56.299.82.160.2]The inability of TB animal 5184 to proliferate to
BLV-containing supernatant and lower counts obtained with BLV
than with MBP in some experiments (Table 3) prompted us to
investigate whether BLV had a suppressive effect on
lympho-cyte proliferation. Surprisingly, concanavalin A (ConA)-induced
proliferation of PBMC from animal 5184 was suppressed to a
broad range of mitogen concentrations by supplementation
with BL3 supernatant. PBMC from TB animal 416 under the
same conditions did not reveal signs of suppression (Table 3).
[image:6.612.68.550.289.681.2]FIG. 6. Lymphocyte proliferation to BLV proteins in PL animals. (A) Fresh PBMC from PL animals 2 and 182 were cultured with BLV antigens at the indicated concentrations of indomethacin for 7 days. Horizontal lines indicate SI 2, and asterisks above columns indicate statistically significant SI. (B) T-cell lines from PL animal 2 and fresh PBMC from PL animal 612 were cultured in medium supplemented with indomethacin (3mg/ml for animal 2 and 6mg/ml for animal 612). Lymphocytes were stimulated with indicated recombinant proteins or BL3 supernatant for 7 days. Results represent the means of at least triplicate wells. Standard deviations are represented by vertical lines. Values above the horizontal line representing SI 2 are statistically significant (marked by asterisks).
TABLE 2. Phenotypes of lymphocytes proliferating to BLV
Cell surfacemarker
% Cellsa
Fresh BLV stimulated
CD2
50
71
CD4
36
67
CD8
13
8
Immunoglobulin M
16
3
WC3
b13
5
aPercentage of PBMC freshly isolated from AL animal 17 or following a
10-day culture with BLV expressing a given marker as determined by flow cytometry.
bWC3 (Workshop cluster 3) designates MAbs to B cells as determined at the
Second International Workshop on Leukocyte Antigens of Cattle and Sheep (23).
on November 9, 2019 by guest
http://jvi.asm.org/
In conclusion, fresh PBMC from TB animals did not
prolifer-ate to recombinant proteins, and indomethacin increased the
mean number of recognized proteins per animal to
approxi-mately 1.
Comparison of proliferative responses to BLV proteins from
animals in different stages of BLV disease.
Comparison of
lymphocyte proliferation to recombinant viral proteins and
BLV from animals in different stages of BLV infection is
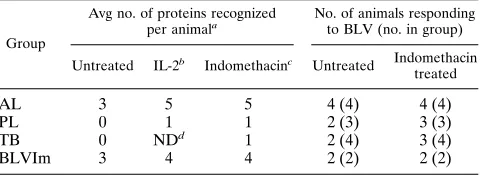
sum-marized in Table 4. The number of recognized proteins per
individual animal by fresh PBMC was highest in the AL group:
animals proliferated to an average of three proteins per
ani-mal. Culture of PBMC in recombinant IL-2 increased the
number of recognized proteins in this group to five per animal.
Fresh PBMC from PL and TB animals did not proliferate to
any of the recombinant proteins. IL-2 treatment and addition
of indomethacin (3 to 6
m
g/ml) to the culture medium
in-creased proliferation and protein recognition in PL and TB
groups to approximately one protein per animal. As found for
recombinant protein recognition, fresh PBMC from the AL
group responded better to BLV than PBMC from PL and TB
animals. Addition of indomethacin was required to elicit
pro-liferation in BLV-nonresponding animals (animal 5184 did not
respond at all) in the PL and TB groups. Responses of animals
from group BLVIm were comparable to those of AL animals.
In summary, proliferation to BLV viral proteins in PL and TB
animals was markedly less than in AL animals. Addition of
indomethacin had a beneficial effect on proliferation of
lym-phocytes from PL and TB animals and helped amplify
prolif-eration in previously negative animals to BLV or selected BLV
proteins.
DISCUSSION
This study demonstrates that CD4
1T-cell proliferation to
Gag and Env proteins was less in PL and TB animals than
in AL or BLVIm animals. Thus, for the first time, advancing
FIG. 7. Proliferation of lymphocytes from TB animals 74, T2, 416, and 5184 to BLV proteins. PBMC were cultured in medium supplemented with the indicated concentration of indomethacin (A to C) or 4.5mg of indomethacin per ml (D) and stimulated with recombinant proteins (4 to 16mg/ml) or 10% BLV-containing BL3 supernatant. Horizontal lines indicate SI 2, and asterisks above columns indicate statistically significant SI. Results represent the means of at least triplicate wells. Standard deviations are represented by vertical lines.
on November 9, 2019 by guest
http://jvi.asm.org/
stages of BLV infection were correlated with dysfunction in
lymphocyte recognition of BLV proteins, indicating that a loss
of T-cell competence may be a factor resulting in acceleration
of infection. AL animals differ from PL animals not only in
lymphocyte counts but also in control of BLV propagation.
The proportion of lymphocytes carrying provirus in PL animals
is 33 to 77% (27, 34), while less than 1% of lymphocytes from
AL animals carry provirus (34). Viral transcripts (18, 24) and
virus expression in vitro (10, 11) are detected most readily in
PL and TB cells and only infrequently in from AL cells. Thus,
our findings provide compelling evidence that less-stringent
virus control in PL and TB animals correlates with impaired
CD4
1T-cell immunity and that the immune system likely
influences the fate of infection. Similar to findings for BLV
infection, dysfunction in CD4
1T-cell antigen-specific
prolifer-ation in asymptomatic human immunodeficiency virus type 1
(HIV-1)-infected patients (9) and selective impairment of
HIV-1-specific cellular response with disease progression (8)
have been described.
CD4
1lymphocytes are pivotal to the immune system. CD4
1T cells regulate cellular and humoral immunity, depending
whether type 1 or type 2 cytokines are produced (25), mediate
protection from viral infection (36), and reject virally induced
tumors (16, 30). Furthermore, antigen-specific CD4
1T
lym-phocytes reportedly sustain CD8
1cytotoxic T-cell responses
during chronic viral infections (33). Therefore, CD4
1T-cell
dysfunction likely leads to a disturbance in CD4
1-driven
im-mune functions that may contribute to progression of BLV
infection with development of PL and tumors. In fact, high
numbers of provirus carrying cells in PL and TB animals
indi-cate that spread of BLV infection to uninfected cells is
facili-tated and/or BLV-infected cells are not effectively eliminated.
Clinically, BLV-infected animals do not exhibit overt
immu-nodeficiency. However, opportunistic infections of
BLV-in-fected cattle with
Trichophyton verrucosum
(6) and increased
frequency of PL animals infected with
Trypanosoma theileri
(21) indicate that disturbances in the immune status of
BLV-infected animals, especially in the PL stage, may be more
profound. Thus, the decrease in CD4
1T-cell competency to
BLV antigens has significant consequences in ameliorating of
BLV infection.
Indomethacin, a cyclo-oxygenase inhibitor, is a potent
sup-pressor of prostaglandin E
2(PGE
2) production by
macro-phages. PGE
2is an immunosuppressant, inhibiting IL-12
pro-duction by macrophages (54), propro-duction of type 1 (IL-2 and
gamma interferon) but not type 2 (IL-4 and IL-5) cytokines by
CD4
1T cells and suppressing mitogen-induced T-cell
prolif-eration (3, 44). Therefore, the beneficial effect of
indometha-cin on antigen-specific lymphocyte proliferation in PL and TB
animals suggests involvement of PGE
2and macrophages in
down-regulating CD4
1T-lymphocyte functions. A similar
find-ing was reported for HIV-1 infection (12), and PGE
2produced
by HIV-1-infected monocytes inhibited T-cell proliferation to
recall antigens. Because infection of monocytes with BLV in
vivo has been reported (47), mechanisms inducing macrophage
PGE
2production by BLV and HIV-1 may be similar.
Interest-ingly, involvement of macrophages as possible mediators of
BLV-induced pathology is supported by our recent finding that
macrophages from PL and TB but not AL animals produce
high amounts of IL-10 mRNA (43). Since IL-10 has a strong
suppressive effect on proliferation of Th1 cells, PGE
2together
with IL-10 may constitute a down-regulatory loop suppressing
immune responses in PL and TB cattle.
Lymphocytes from AL animals proliferated to recombinant
viral proteins, and antigen-pulsed T cells expanded in the
pres-ence of IL-2 increased the repertoire of recognized proteins,
suggesting the expansion of low-frequency antigen-specific
precursor cells. B lymphocytes from BLV-infected animals are
activated cycling cells responsive to IL-2 (32). Therefore, the
superior effect of a low level of recombinant IL-2 (2 U/ml)
compared with high levels (10 and 20 U/ml [data not shown])
in cultures from AL animals could be explained by the selective
IL-2 action on antigen-activated T cells expressing high-affinity
IL-2 receptor (IL-2R) but not B cells that fail to express the
high-affinity receptor. However, IL-2 treatment of PL and TB
cells did not result in expansion of antigen-specific T cells
except in one PL animal. Presumably, B cells from PL and TB
animals are more responsive to IL-2, and 2 U/ml was sufficient
for B-cell proliferation and expansion. This explanation is
sup-ported by the finding that B-cell expression of the IL-2R
a
chain in mitogen-stimulated cells from PL animals is greater
than in AL animals (50). Alternatively, CD4
1T-cell
antigen-specific proliferation from IL-2-expanded PL and TB cells may
not have been detected because T cells from PL animals
re-portedly have altered proliferation and IL-2 production (49).
In HTLV-1 infection, dysregulation and overexpression of the
IL-2R
a
chain and autocrine stimulation involving IL-2 are
hypothesized to be the early events leading to leukemogenesis
(56). Transactivation of IL-2 and IL-2R genes by BLV Tax has
not been reported; however, suppression of type 1 IL-2-driven
activation and the preferential bias of type 2 cytokines
pro-voked by PGE
2and IL-10 could alter the regulatory
equilib-rium in T-T cell and T-B cell interactions and contribute to
BLV pathogenesis.
[image:8.612.59.299.91.159.2]Despite difficulties in proliferation to individual BLV
pro-teins, PL and TB animals (with the exception of animal 5184)
proliferated when BLV-containing medium was used as the
TABLE 3. Suppression of PBMC proliferation from
TB animal 5184 by BLV
Animal
SIa
Expt 1b Expt 2,
c
with ConA concn (mg/ml) of:
2.5 1.25 0.6 0.3 0.15
5184
0.4
0.76
0.74
0.77
0.98
0.70
416
1.36
1.13
1.48
1.33
1.13
1.27
a
Calculated as counts obtained with ConA plus BLV divided by counts ob-tained with ConA alone.
b
PBMC were stimulated by 10% BLV containing BL3 supernatant, and MBP was used to determine background proliferation.
c
PBMC cultured in 10% FS were stimulated with indicated concentrations of ConA or ConA plus 25% BL3 supernatant.
TABLE 4. Comparison of CD4
1T-cell proliferative responses to
BLV proteins from animals in different stages of BLV infection
Group
Avg no. of proteins recognized
per animala No. of animals responding to BLV (no. in group)
Untreated IL-2b Indomethacinc Untreated Indomethacin treated
AL
3
5
5
4 (4)
4 (4)
PL
0
1
1
2 (3)
3 (3)
TB
0
ND
d1
2 (4)
3 (4)
BLVIm
3
4
4
2 (2)
2 (2)
aValues were rounded to the closest integer.
bSum of proliferative responses from untreated and IL-2-treated cells.
cSum of proliferative responses from untreated, IL-2-treated, and
indometh-acin-treated cells. dND, not determined.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.317.557.598.686.2]antigen. The exact mechanism(s) leading to poor proliferation
to recombinant proteins is unclear; however, several
possibil-ities exist. First, macrophages may be deficient in processing
soluble peptides but not particulate antigens. BL3 supernatant
was subjected to repeated freeze-thaw cycles to disintegrate
the viral particles; however, this procedure may destroy BLV
infectivity mostly by removing surface glycoproteins, and
de-struction of the viral core (particulate antigen form) could
remain incomplete. Interestingly, in patients with AIDS, a
fail-ure of lymphocytes to recognize and respond to soluble antigen
has been found (29). Second, the precursor frequency of
anti-gen specific CD4
1T cells to individual proteins may be low,
and weak proliferation may remain undetectable. However,
when viral particles containing several proteins are added,
synergistic responses to individual proteins may result in
de-tectable proliferation. Third, proliferation to BLV virions may
result from additional proteins in BLV virions, such as
pro-tease and reverse transcriptase/integrase. Even though the
rel-ative content of these proteins in BLV virions is low, this
possibility cannot be excluded. Interestingly, reverse
transcrip-tase from HIV-1 was identified as a source of CD4
1and CD8
1T-cell epitopes (17, 22). Inhibition of immune functions often
accompanying retroviral infections can be partially attributed
to an immunosuppressive nature of certain retroviral proteins
(20, 51). BLV inhibition of ConA proliferation in lymphocytes
from TB animal 5184 may be caused by similar mechanisms.
In conclusion, our data demonstrate that progression of
BLV infection to advancing stages, PL and lymphosarcoma,
was correlated with dysfunction of CD4
1T cells in response to
BLV antigens. This finding strongly indicates that subversion
of cellular immunity by BLV is a factor triggering progression
of infection, thus providing further insight into the
pathogen-esis of BLV infection.
ACKNOWLEDGMENTS
This research was supported by the College of Agricultural and Life
Sciences and grants RO1.CA59127 from the National Institutes of
Health and BARD 95-34339-2556 from the Binational Agricultural
Research and Development Fund.
REFERENCES
1.Abt, D. A., R. R. Marshak, H. W. Kulp, and R. J. Pollock, Jr.1970. Studies on the relationship between lymphocytosis and bovine leukemia. Bibl. Haemat.36:527–536.
2.Altaner, C., J. Ban, V. Altanerova, and V. Janik.1991. Protective vaccination against bovine leukemia virus infection by means of cell-derived vaccine. Vaccine9:889–895.
3.Betz, M., and B. S. Fox.1991. Prostaglandin E2inhibits production of Th1 lymphokines but not of Th2 lymphokines. J. Immunol.146:108–113. 4.Bex, F., C. Bruck, M. Mammerickx, D. Portetelle, J. Ghysdael, Y. Cleuter, M.
Leclercq, D. Dekegel, and A. Burny.1979. Humoral antibody response to bovine leukemia virus infection in cattle and sheep. Cancer Res.39:1118– 1123.
5.Birkebak, T. A., G. H. Palmer, W. C. Davis, D. P. Knowles, and T. F. McElwain.1994. Association of gp51 expression and persistent CD51 B-lymphocyte expansion with lymphomagenesis in bovine leukemia virus in-fected sheep. Leukemia8:1890–1899.
6.Brenner, J., M. Van-Haam, D. Savir, and Z. Trainin.1989. The implications of BLV infection in the productivity, reproductive capacity and survival rate of a dairy cow. Vet. Immunol. Immunopathol.22:299–305.
7.Callebaut, I., V. Voneche, A. Mager, O. Fumiere, V. Krchnak, M. Merza, J. Zavada, M. Mammerickx, A. Burny, and D. Portetelle.1993. Mapping of B-neutralizing and T-helper cell epitopes on the bovine leukemia virus external glycoprotein gp51. J. Virol.67:5321–5327.
8.Carmichael, A., X. Jin, P. Sissons, and L. Borysiewicz.1993. Quantitative analysis of the human immunodeficiency virus type 1 (HIV-1)-specific cyto-toxic T lymphocyte (CTL) response at different stages of HIV-1 infection: differential CTL responses to HIV-1 and Epstein-Barr virus in late disease. J. Exp. Med.177:249–256.
9. Clerici, M., N. I. Stocks, R. A. Zajac, R. N. Boswell, D. R. Lucey, C. S. Via, and G. M. Shearer.1989. Detection of three distinct patterns of T helper cell dysfunction in asymptomatic, human immunodeficiency virus-seropositive
patients. J. Clin. Invest.84:1892–1899.
10. Cockerell, G. L., and J. Rovnak.1988. The correlation between the direct and indirect detection of bovine leukemia virus infection in cattle. Leukemia Res.12:465–469.
11. Ferrer, J. F., R. R. Marshak, D. A. Abt, and S. J. Kenyon.1978. Persistent lymphocytosis in cattle: its cause, nature and relation to lymphosarcoma. Ann. Rech. Vet.9:851–857.
12. Foley, P., F. Kazazi, R. Biti, T. C. Sorrell, and A. L. Cunningham.1992. HIV infection of monocytes inhibits the T-lymphocyte proliferative response to recall antigens, via production of eicosanoids. Immunology75:391–397. 13. Gatei, M. H., M. F. Good, R. C. W. Daniel, and M. F. Lavin.1993. T-cell
responses to highly conserved CD4 and CD8 epitopes on the outer mem-brane protein of bovine leukemia virus: relevance to vaccine development. J. Virol.67:1796–1802.
14. Gatei, M. H., H. M. Naif, S. Kumar, D. B. Boyle, R. C. W. Daniel, M. F. Good, and M. F. Lavin.1993. Protection of sheep against bovine leukemia virus (BLV) infection by vaccination with recombinant vaccinia viruses ex-pressing BLV envelope glycoproteins: correlation of protection with CD4 T-cell response to gp51 peptide 51-70. J. Virol.67:1803–1810.
15. Ghysdael, J., R. Kettmann, and A. Burny.1978. Translation of bovine leu-kemia virus genome information in heterologous protein synthesizing sys-tems programmed with virion RNA and in cell-lines persistently infected by BLV. Ann. Rech. Vet.9:627–634.
16. Greenberg, P. D., M. A. Cheever, and A. Feffer.1981. Eradication of dissem-inated murine leukemia by chemoimmunotherapy with cyclophosphamide and adoptively transferred immune syngeneic Lyt-1122lymphocytes. J. Exp. Med.154:952–963.
17. Haas, G., R. David, R. Frank, H. Gausepohl, C. Devaux, J. M. Claverie, and M. Pierres.1991. Identification of a major human immunodeficiency virus-1 reverse transcriptase epitope recognized by mouse CD41T lymphocytes. Eur. J. Immunol.21:1371–1377.
18. Haas, L., T. Divers, and J. W. Casey.1992. Bovine leukemia virus gene expression in vivo. J. Virol.66:6223–6225.
19. Hager, D. A., and R. R. Burgess.1980. Elution of proteins from sodium dodecyl sulfate-polyacrylamide gels, removal of sodium dodecyl sulfate, and renaturation of enzymatic activity: results with sigma subunit ofEscherichia coliRNA polymerase, wheat germ DNA topoisomerase, and other enzymes. Anal. Biochem.109:76–86.
20. Haraguchi, S., R. A. Good, and N. K. Day.1995. Immunosuppressive retro-viral peptides: cAMP and cytokine patterns. Immunol. Today16:595–603. 21. Hare, W. C. D., E. J. L. Soulsby, and D. A. Abt.1969. Bovine trypanosomiasis
and lymphocytosis parallel studies. Bibl. Haemat.36:504–517.
22. Hosmalin, A., M. Clerici, R. Houghten, C. D. Pendleton, C. Flexner, D. R. Lucey, B. Moss, R. N. Germain, G. M. Shearer, and J. A. Berzofsky.1990. An epitope in human immunodeficiency virus 1 reverse transcriptase recognized by both mouse and human cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. USA87:2344–2348.
23. Howard, C. J., and J. Naessens.1993. Summary of workshop findings for cattle. Vet. Immunol. Immunopathol.39:25–48.
24. Jensen, W. A., J. Rovnak, and G. L. Cockerell.1991. In vivo transcription of the bovine leukemia virustax/rexregion in normal and neoplastic lympho-cytes of cattle and sheep. J. Virol.65:2484–2490.
25. Kelso, A.1995. Th1 and Th2 subsets: paradigms lost? Immunol. Today
16:374–379.
26. Kenyon, S. J., and C. E. Piper.1977. Cellular basis of persistent lymphocy-tosis in cattle infected with bovine leukemia virus. Infect. Immun.16:891– 897.
27. Kettmann, R., Y. Cleuter, M. Mammerickx, M. Meunier-Rotival, G. Ber-nardi, A. Burny, and H. Chantrenne.1980. Genomic integration of bovine leukemia provirus: comparison of persistent lymphocytosis with lymph node tumor form of enzootic bovine leukosis. Proc. Natl. Acad. Sci. USA77:2577– 2581.
28. Kono, Y., K. Arai, H. Sentsui, S. Matsukawa, and S. Itohara.1986. Protec-tion against bovine leukemia virus infecProtec-tion in sheep by active and passive immunization. Jpn. J. Vet. Sci.48:117–125.
29. Lane, H. C., J. M. Depper, W. C. Greene, G. Whalen, T. A. Waldmann, and A. S. Fauci.1985. Qualitative analysis of immune function in patients with the acquired immunodeficiency syndrome. Evidence for a selective defect in soluble antigen recognition. N. Engl. J. Med.313:79–84.
30. Leclerc, J. C., and H. Cantor.1980. T cell-mediated immunity to oncorna-virus-induced tumors. II. Ability of different T cell sets to prevent tumor growthin vivo. J. Immunol.124:851–854.
31. Mager, A., R. Masengo, M. Mammerickx, and J. J. Letesson.1994. T cell proliferative responses to bovine leukemia virus (BLV): identification of T cell epitopes on the major core protein (p24) in BLV-infected cattle with normal haematological values. J. Gen. Virol.75:2223–2231.
32. Matheise, J. P., M. Delcommenne, A. Mager, C. H. Didembourg, and J. J. Letesson.1992. CD51B cells from bovine leukemia virus infected cows are activated cycling cells responsive to interleukin 2. Leukemia6:304–309. 33. Matloubian, M., R. J. Concepcion, and R. Ahmed.1994. CD41cells are
required to sustain CD81cytotoxic T-cell responses during chronic viral infection. J. Virol.68:8056–8063.
on November 9, 2019 by guest
http://jvi.asm.org/
34. Mirsky, M. L., Y. Da, and H. A. Lewin.1993. Detection of bovine leukemia virus proviral DNA in individual cells. PCR Methods Appl.2:333–340. 35. Muscoplat, C., D. W. Johnson, K. A. Pomeroy, J. M. Olson, V. L. Larson,
J. B. Stevens, and D. K. Sorensen.1974. Lymphocyte surface immunoglob-ulin: frequency in normal and lymphocytotic cattle. Am. J. Vet. Res.35:593– 595.
36. Neal, Z., and G. A. Splitter.1995. Picornavirus-specific CD41T lymphocytes possessing cytolytic activity confer protection in the absence of prophylactic antibodies. J. Virol.69:4914–4923.
37. Ohishi, K., H. Suzuki, T. Yamamoto, T. Maruyama, K. Miki, Y. Ikawa, S. Numakunai, K. Okada, K. Ohshima, and M. Sugimoto.1991. Protective immunity against bovine leukemia virus (BLV) induced in carrier sheep by inoculation with a vaccinia virus-BLVenvrecombinant: association with cell-mediated immunity. J. Gen. Virol.72:1887–1892.
38. Ohishi, K., H. Suzuki, Y. Yasutomi, M. Onuma, K. Okada, S. Numakunai, K. Ohshima, Y. Ikawa, and M. Sugimoto.1992. Augmentation of bovine leukemia virus (BLV)-specific lymphocyte proliferation responses in rumi-nants by inoculation with BLVenv-recombinant vaccinia virus: their role in the suppression of BLV replication. Microbiol. Immunol.36:1317–1323. 39. Onuma, M., T. Hodatsu, S. Yamamoto, M. Higashihara, S. Masu, T.
Mi-kami, and H. Izawa.1984. Protection by vaccination against bovine leukemia virus infection in sheep. Am. J. Vet. Res.45:1212–1215.
40. Orlik, O., J. Ban, E. Gieciova, V. Altanerova, and C. Altaner.1993. Prepa-ration and characterization of monoclonal antibodies directed against gly-coproteins of bovine leukemia virus. Acta Virol.37:377–387.
41. Orlik, O., J. Ban, E. Gieciova, V. Altanerova, and C. Altaner.1993. Two immunodominant regions revealed by monoclonal antibodies on the main structural protein p24 of bovine leukemia virus. Viral Immunol.6:245–254. 42. Orlik, O., and G. A. Splitter.Optimization of lymphocyte proliferation assay for cells with high spontaneous proliferationin vitro: CD41T-cell prolifer-ation in bovine leukemia virus infected animals with persistent lymphocyto-sis. J. Immunol. Methods, in press.
43. Peyon, D., K. L. O’Reilly, and G. A. Splitter.1996. Increased interleukin-10 mRNA expression in tumor bearing or persistently lymphocytotic animals infected with bovine leukemia virus. J. Virol.70:5706–5710.
44. Phipps, R. P., S. H. Stein, and R. L. Roper.1991. A new view of prostaglan-din E regulation of the immune response. Immunol. Today12:349–352.
45. Portetelle, D., K. Limbach, A. Burny, M. Mammerickx, P. Desmettre, M. Riviere, J. Zavada, and E. Paoletti.1991. Recombinant vaccinia virus ex-pression of the bovine leukaemia virus envelope gene and protection of immunized sheep against infection. Vaccine9:194–200.
46. Sagata, N., T. Yasunaga, J. Tsuzuku-Kawamura, K. Ohishi, Y. Ogawa, and Y. Ikawa.1985. Complete nucleotide sequence of the genome of bovine leukemia virus: its evolutionary relationship to other retroviruses. Proc. Natl. Acad. Sci. USA82:677–681.
47. Schwartz, I., A. Bensaid, B. Polack, B. Perrin, M. Berthelemy, and D. Levy.
1994. In vivo leukocyte tropism of bovine leukemia virus in sheep and cattle. J. Virol.68:4589–4596.
48. Schwartz, I., and D. Levy.1994. Pathobiology of bovine leukemia virus. Vet. Res.25:521–536.
49. Sordillo, L. M., C. R. Hicks, and G. M. Pighetti.1994. Altered interleukin-2 production by lymphocyte populations from bovine leukemia virus-infected cattle. Proc. Soc. Exp. Biol. Med.207:268–273.
50. Stone, D. M., T. F. McElwain, and W. C. Davis.1994. Enhanced B-lympho-cyte expression of IL-2Ra associated with T lymphocytosis in BLV-infected persistently lymphocytotic cows. Leukemia8:1057–1061.
51. Subramanyam, M., W. G. Gutheil, W. W. Bachovchin, and B. T. Huber.
1993. Mechanism of HIV-1 Tat induced inhibition of antigen-specific T cell responsiveness. J. Immunol.150:2544–2553.
52. Thurmond, M. C., R. L. Carter, J. P. Picanso, and K. Stralka.1990. Upper-normal prediction limits of lymphocyte counts for cattle not infected with bovine leukemia virus. Am. J. Vet. Res.51:466–470.
53. Thurmond, M. C., C. H. Holmberg, and J. P. Picanso.1985. Antibodies to bovine leukemia virus and presence of malignant lymphoma in slaughtered California dairy cattle. J. Natl. Cancer Inst.74:711–714.
54. van der Pouw Kraan, T., L. C. M. Boeije, R. J. T. Smeenk, J. Wijdenes, and L. A. Aarden.1995. Prostaglandin-E2 is a potent inhibitor of human inter-leukin 12 production. J. Exp. Med.181:775–779.
55. Xu, A., M. J. T. van Eijk, C. Park, and H. A. Lewin.1993. Polymorphism in BoLA-DRB3 exon 2 correlates with resistance to persistent lymphocytosis caused by bovine leukemia virus. J. Immunol.151:6977–6985.
56. Yodoi, J., and T. Uchiyama.1992. Diseases associated with HTLV-I: virus, IL-2 receptor dysregulation and redox regulation. Immunol. Today13:405– 411.