Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Role of Single-Stranded DNA Binding Activity of T Antigen in
Simian Virus 40 DNA Replication
CHUNXIAO WU, RUPA ROY,ANDDANIEL T. SIMMONS*
Department of Biological Sciences, University of Delaware, Newark, Delaware 19716-2590
Received 2 October 2000/Accepted 22 December 2000
We have previously mapped the single-stranded DNA binding domain of large T antigen to amino acid residues 259 to 627. By using internal deletion mutants, we show that this domain most likely begins after residue 301 and that the region between residues 501 and 550 is not required. To study the function of this binding activity, a series of single-point substitutions were introduced in this domain, and the mutants were tested for their ability to support simian virus 40 (SV40) replication and to bind to single-stranded DNA. Two replication-defective mutants (429DA and 460EA) were grossly impaired in single-stranded DNA binding. These two mutants were further tested for other biochemical activities needed for viral DNA replication. They bound to origin DNA and formed double hexamers in the presence of ATP. Their ability to unwind origin DNA and a helicase substrate was severely reduced, although they still had ATPase activity. These results suggest that the single-stranded DNA binding activity is involved in DNA unwinding. The two mutants were also very defective in structural distortion of origin DNA, making it likely that single-stranded DNA binding is also required for this process. These data show that single-stranded DNA binding is needed for at least two steps during SV40 DNA replication.
Simian virus 40 (SV40) has been used extensively as a model to study eukaryotic DNA replication. The virus encodes a multifunctional protein, T antigen, which is the only viral tein required for DNA replication. All other factors are pro-vided by the host cell (63). To initiate DNA replication, T antigen binds to the viral origin, which has two pairs of GAGGC pentanucleotides arranged in a head-to-head orien-tation (15, 65, 66). In the presence of ATP, T antigen assem-bles into a double hexamer by binding to the pentanucleotides (13, 25, 33) and causes structural changes in the flanking DNA sequences. The DNA at the early palindrome (EP) is melted and the AT-rich region is untwisted (4, 5, 12, 41, 45). Then T antigen functions as a helicase to bidirectionally unwind the DNA in the presence of replication protein A (RPA) and topoisomerase I (14, 16, 21, 76). Other cellular factors, includ-ing DNA polymerase␣-primase, RFC, PCNA, and DNA poly-merase␦, are also recruited to form the replication fork (36, 39, 40, 44, 67, 70, 71).
Many of the functional domains of T antigen have been well characterized. The origin DNA binding domain is located near the N-terminal end from residues 147 to 247 (1, 35, 53, 54) and by itself has the ability to interact with origin DNA (1, 26, 35). As a helicase, T antigen unwinds DNA in a 3⬘to 5⬘direction coupled with ATP hydrolysis (21, 60, 75). The region required for this activity maps from residues 131 to 616 (78). The ATP binding domain is located between residues 418 and 528 (7), and the ATPase domain extends from residues 418 to 616 (9, 10).
We have recently started to study T antigen’s single-stranded DNA binding activity (77). The exact function of this activity is not known, but there are a number of reasons for believing that
it is required for DNA unwinding. When T antigen is incu-bated with a synthetic DNA fork in the presence of ATP, it assembles on the fork as a hexamer and unwinds it (50, 58, 74). During this process, T antigen interacts primarily with the 3⬘
single strand with slight protection of the duplex DNA close to the branch point (50). On a replication bubble, T antigen binds as a double hexamer with twofold symmetry and strongly pro-tects the same (3⬘) strand at each junction (58). It is proposed that during unwinding T antigen forms a double hexamer with each strand of DNA being pulled through the central channel of different hexamers (58). These observations point to an active role of single-stranded DNA binding during the unwind-ing process.
T antigen belongs to helicase superfamily 3, which includes helicases from small DNA and RNA viruses (22). Based on sequence alignments and secondary structure predictions, Masterson et al. (32) suggested that the C-terminal region of the helicases in this family serves as the helicase domain. This helicase domain on T antigen also shows homology and anti-body cross-reaction withEscherichia coliRecA (49). In recent years, the crystal structures of several helicases from super-families 1, 2, and 4 have been resolved. Monomeric or dimeric helicases, such asBacillus stearothermophilusPcrA (62, 69),E. coliRep (29), hepatitis C virus NS3 (28, 80), and UvrB (64) all contain two RecA-like domains that join to form a nucleotide binding site at their interface. Similarly, each subunit of hex-americ bacteriophage T7 gene 4 helicase contains a RecA-like domain (46, 57). Domains on adjacent subunits combine to form a nucleotide binding site. These similarities suggest that the ATP binding domain of RecA is the unifying structure of all helicases (2, 81). However, it does not appear that helicases use a common mechanism of action. PcrA binds and destabi-lizes double-stranded DNA ahead of the fork by using its 1B and 2B domains (59, 69), whereas NS3 passively unwinds the DNA by binding to single-stranded DNA generated from ther-* Corresponding author. Mailing address: Department of Biological
Sciences, University of Delaware, Newark, DE 19716-2590. Phone: (302) 831-8547. Fax: (302) 831-2281. E-mail: [email protected].
2839
on November 9, 2019 by guest
http://jvi.asm.org/
mal fluctuations at the fork (28, 43). During translocation, PcrA binds to single-stranded DNA as a monomer through the two RecA-like domains (69). For hexameric T7 gene 4 heli-case, it is proposed that each subunit sequentially binds to single-stranded DNA in the central channel, as suggested by the asymmetric arrangement of the six subunits in the hexamer (57).
To probe the possible role of the single-stranded DNA bind-ing domain of T antigen in DNA unwindbind-ing and to determine if it has other functions, we generated a series of internal deletion and single-point substitution mutants in this domain. The mutants’ abilities to bind single-stranded DNA and to perform other biochemical reactions related to DNA replica-tion were tested. The results demonstrate a strong correlareplica-tion between single-stranded DNA binding and unwinding the or-igin and a helicase substrate. We also show a correlation with the mutants’ abilities to structurally distort the origin.
MATERIALS AND METHODS
Oligonucleotide-directed mutagenesis.Mutations were generated as described by Loeber et al. (31). Oligonucleotides containing desired point mutations were phosphorylated and annealed to uridine-containing single-stranded pBS-SV40 (31) or pSK(⫺)SVTC (30). The complementary strand was synthesized by T4 polymerase and sealed by T4 ligase. The DNA was used to transformE. coli
JM109 cells. The colonies were screened for correct mutations by standard dideoxy DNA sequencing.
Construction of recombinant baculoviruses. For making internal deletion mutants, T antigen cDNAs with different internal deletions (27) (obtained from Judy Tevethia) were used as templates for PCR. DNA constructs were missing between 14 and 82 codons and contained, in their place, 3 extra non-T-antigen codons (27). An oligonucleotide with a start site at amino acid residue 246 was used as the up primer; the down primer had a stop codon after residue 708. The PCR DNAs were cut with appropriate restriction enzymes and inserted into pVL1393 baculovirus transfer vector (BD PharMingen). All clones were con-firmed by DNA sequencing.
For making single-point-substitution-mutant T antigens, mutant pSK(⫺) SVTC DNA was cut withBamHI and ligated withBamHI-cleaved pVL941 DNA (BD PharMingen). The pVL1393 or pVL941 DNA with T-antigen inserts was cotransfected with BaculoGold DNA (BD PharMingen) in Sf9 insect cells ac-cording to the manufacturer’s protocol. T-antigen-expressing recombinants were detected by immunofluorescence analysis and purified by plaque assays.
T-antigen purification.Wild-type and mutant T antigens were expressed in baculovirus-infected insect cells and purified by immunoaffinity chromatography with monoclonal antibody PAb101 as previously described (19). The internal deletion mutants were eluted with buffer E (20 mM triethylamine [pH 10.8], 10% glycerol) (52), and the point substitution mutants were eluted with ethylene glycol elution buffer (50% ethylene glycol, 20 mM Tris [pH 8.5], 500 mM NaCl, 1mM EDTA, 10% glycerol) (33). The T antigens were dialyzed against dialysis storage buffer (10 mM Tris [pH 8.0], 100 mM NaCl, 1 mM EDTA, 1 mM dithiothreitol, 50% glycerol) and stored at⫺20°C.
Viral replication assay.pBS-SV40 DNA containing a point mutation was cut withBamHI to release the SV40 genomic DNA. The DNA was then ligated at low concentrations to get circular SV40 DNA. The DNA was transfected into BSC-1 monkey kidney cells by using DEAE-dextran (34). On day 10, the cells were stained with neutral red and the sizes of the plaques were compared to those formed by the wild type.
Gel shift assays.For single-stranded DNA binding, a32P-end-labeled 55-mer single-stranded DNA corresponding to the bottom strand of the fork substrate (50) was used. A 112-bp origin-containingHindIII-NcoI SV40 DNA fragment from pSKori (56) was end-labeled with32P and used in origin DNA binding reactions. The reactions were performed in replication buffer (30 mM HEPES-KOH [pH 7.5], 7 mM MgCl2, 40 mM creatine phosphate, 4 mM ATP, 1 mM dithiothreitol, 0.1 mg of bovine serum albumin per ml). After incubation of T antigen with DNA for 30 min at 37°C, glutaraldehyde was added to a final concentration of 0.1% and the reaction mixtures were incubated for an addi-tional 15 min at 37°C. The samples were loaded on composite gels (2.5% acrylamide and 0.6% agarose) and subjected to electrophoresis at 25 mA for 3 h
in Tris-borate-EDTA buffer at 4°C. The gels were dried and exposed to X-ray film.
Origin DNA unwinding.The substrate for origin DNA unwinding was a32 P-end-labeled 112-bpHindIII-to-NcoI origin-containing fragment. T antigen was incubated with the substrate in replication buffer supplemented with 50g of creatine phosphokinase per ml and 2.8g of E. colisingle-stranded DNA binding protein (Pharmacia) per ml. After 1 h at 37°C, the reaction was termi-nated by adding EDTA to 20 mM, sodium dodecyl sulfate to 0.5%, and protein-ase K to 0.2 mg/ml and the mixtures were incubated for an additional 30 min. After the samples were heated for 5 min at 65°C, they were loaded on 7% acrylamide gels and subjected to electrophoresis for 600 V䡠h at 4°C.
ATPase assays.Six hundred nanograms of T antigen was incubated with [␥-32P]ATP in ATPase buffer (8). After incubation at room temperature for 1 h, the mixture was spotted on polyethyleneimine-cellulose thin-layer chromatogra-phy plates and subjected to ascending chromatograchromatogra-phy in 0.75 M KH2PO4(6). The released inorganic phosphate and ATP were quantitated with a Phosphor-Imager (Molecular Dynamics).
Helicase assays.The helicase assay was based on the method of Stahl (60). The helicase substrate was made by annealing a 32-mer primer (5⬘CCAGGGT TTTCCCAGTCACGACGTTGTAAAAC 3⬘) to M13mp19 single-stranded DNA. The primer was extended by Klenow DNA polymerase in the presence of dCTP and [␣-32P]dATP at room temperature for 20 min, followed by an addi-tional 20-min incubation at room temperature with unlabeled dATP. The sub-strate was purified on a Sephadex G-50 column (Pharmacia). T antigen was incubated with the substrate in helicase buffer (60) for 30 min at 37°C. The reaction was stopped by adding sodium dodecyl sulfate to a final concentration of 0.2% and EDTA to 50 mM. The displaced primer was resolved by electro-phoresis on 10% acrylamide gels for 360 V䡠h at 4°C. The gels were dried and subjected to autoradiography.
KMnO4assays.KMnO4oxidation was used to detect the melting and untwist-ing of origin DNA as described by Borowiec and Hurwitz (4) and Parsons et al. (41). Four hundred to 2,000 ng of T antigen was incubated with 600 ng of origin-containing plasmid pSVO⌬1 (61) in replication buffer. After 30 min at 37°C, KMnO4 was added to a final concentration of 6 mM, followed by an additional 4-min incubation. The reaction was stopped by adding 2-mercapto-ethanol to a final concentration of 1 M. The DNA was purified through a Sephadex G-50 column and subjected to primer extension assays using a32 P-end-labeled pBR322EcoRI primer (New England Biolabs). The samples were loaded on a 7% sequencing gel, and the gel was dried and exposed to X-ray film. The melting and untwisting signals were quantitated with a PhosphorImager.
RESULTS
Single-stranded DNA binding activity of deletion mutants. The single-stranded DNA binding domain of T antigen has been mapped to amino acid residues 259 through 627 (77). In order to more precisely map this activity, we made a series of mutants with internal deletions (between 14 and 82 amino acids) within the single-stranded DNA binding domain. All of these mutant proteins began at amino acid residue 246 and ended at residue 708, with additional deletions spanning be-tween residues 251 and 550. Similar internal deletion mutants of T antigen were made by Kierstead and Tevethia (27) to map the p53 binding region. The mutants were expressed in recom-binant baculoviruses and purified by immunoaffinity chroma-tography. The single-stranded DNA binding activity of these 13 mutants was tested by gel shift experiments (Fig. 1). A mutant with a deletion of residues 251 to 300 could form hexamers on single-stranded DNA (Fig. 1, lane 2), although the binding was weaker than that with wild-type T antigen. Another mutant missing amino acids 501 to 550 also formed hexamers on single-stranded DNA at a level close to that of the wild type (Fig. 1, lane 14). This indicates that regions 251 to 300 and 501 to 550 are not required for single-stranded DNA binding. Mutants with deletions of residues 301 to 350, 354 to 400, 347 to 370, 369 to 400, 382 to 400, 401 to 436, 437 to 450, 451 to 465, 451 to 489, 451 to 500, and 451 to 532 failed to form
on November 9, 2019 by guest
http://jvi.asm.org/
hexamers on single-stranded DNA (Fig. 1, lanes 3 to 13). This result suggests that the region between residues 301 and 500 is important for single-stranded DNA binding.
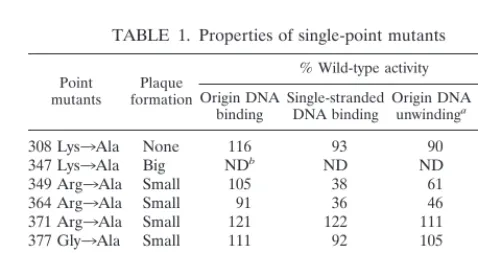
Viral replication of single-point mutants.To better charac-terize the function of the single-stranded DNA binding activity of T antigen, we generated mutants with single-point substitu-tions within the region identified above. The residues that were changed were selected on the basis of a weak sequence simi-larity with RPA, a single-stranded DNA binding protein. Since the residues in RPA that contact single-stranded DNA are known (3, 42), we chose nine corresponding amino acids in T antigen and mutagenized them to alanines. Mutations were first introduced into the entire virus genome (in pBS-SV40) to assess their effect on viral replication. The viral DNA was excised from the plasmid, religated into circular DNA, and transfected into BSC-1 monkey kidney cells. The ability of these mutants to form plaques was evaluated after 10 days (Table 1). Mutants 347KA and 387WA formed large plaques, just like the wild type. Mutants 349RA, 364RA, 371RA, and 377GA formed small plaques, and mutants 308KA, 429DA, and 460EA did not form any plaques under our experimental conditions. This showed that mutations of residues 308, 349, 364, 371, 377, 429, and 460 caused a defect in viral replication. Single-stranded DNA and origin DNA binding by single-point mutants. To determine whether the virus replication-defective T-antigen mutants were also replication-defective in binding to
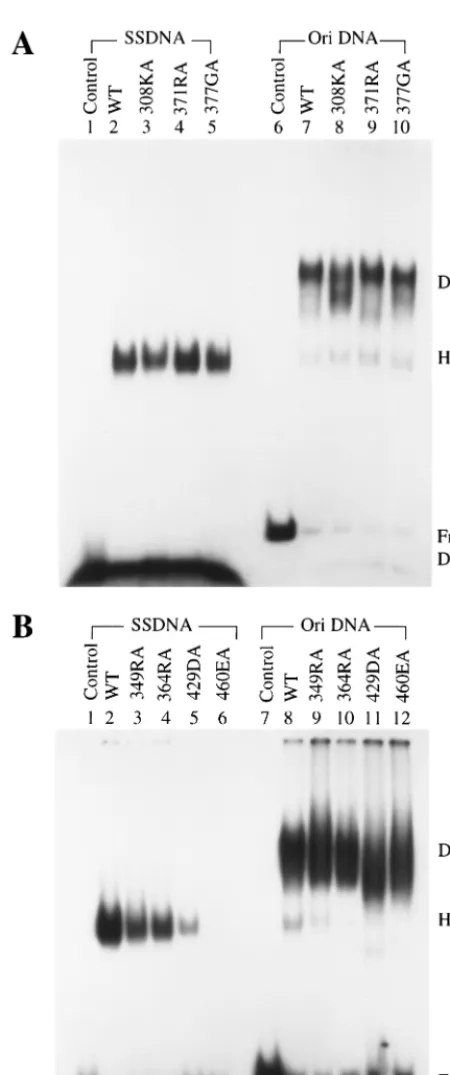
single-stranded DNA, seven equivalent mutations were intro-duced in the T-antigen cDNA to generate alanine substitution mutants at residues 308, 349, 364, 371, 377, 429, and 460. T antigens were expressed from recombinant baculoviruses and purified by immunoaffinity chromatography. Their binding to single-stranded DNA and origin DNA was tested in gel shift assays (Fig. 2 and Table 1). Mutants 308KA, 371RA, and 377GA had normal single-stranded DNA binding activity (Fig. 2A, lanes 3, 4, and 5), whereas that of mutants 349RA, 364RA, and 429DA was weaker than that of the wild type (Fig. 2B, lanes 3, 4, and 5), and mutant 460EA failed to bind to any single-stranded DNA (Fig. 2B, lane 6). Although mutants 349RA, 364RA, and 429DA bound single-stranded DNA poorly, they were still able to form hexamers on the DNA, just like the wild type (Fig. 2B, lanes 2 to 5). All of these mutants had the ability to bind to origin DNA as double hexamers (Fig. 2A, lanes 8 to 10, and B, lanes 9 to 12), although 308KA and 429DA might have formed slightly smaller complexes. These results are consistent with the previous observation (77) that the single-stranded DNA and origin DNA binding domains are distinct.
Unwinding of origin DNA by point mutants. The current helicase model shows that T-antigen hexamers encircle only one strand of DNA during unwinding (18, 58). This suggests that single-stranded DNA binding activity is needed for DNA unwinding. To investigate this possibility, mutant T antigens were examined for their ability to unwind an origin DNA fragment in the presence ofE. colisingle-stranded DNA bind-ing protein as previously described (55). The results are shown in Table 1. Mutants 308KA, 371RA, and 377GA were able to unwind double-stranded DNA into single-stranded DNA. Mu-tants 349RA and 364RA had significantly less unwinding ac-tivity, and mutant 429DA had very little activity. Unwinding activity by mutant 460EA was undetectable. By comparing these results with those measuring single-stranded DNA bind-ing (Fig. 2 and Table 1), it is apparent that mutants with impaired abilities to bind single-stranded DNA were also de-fective in origin DNA unwinding.
[image:3.612.83.266.72.343.2]ATPase activity of point mutants.For T antigen to unwind double-stranded DNA, it needs to hydrolyze ATP in order to provide the energy needed to separate the two strands. There are two possible reasons why mutants 349RA, 364RA, 429DA, FIG. 1. Single-stranded DNA binding assay of wild-type (WT) and
mutant T antigens. Ten picomoles (corresponding to 800 ng of wild-type T antigen) wild-wild-type or N-terminal deletion mutant T antigens (246 to 708) with an extra internal deletion was incubated with end-labeled single-stranded DNA for 30 min at 37°C, cross-linked with glutaraldehyde, and applied to a composite gel (0.6% agarose and 2.5% acrylamide) in Tris-borate-EDTA buffer. The positions of free DNA and bound DNA are shown.
TABLE 1. Properties of single-point mutants
Point
mutants formationPlaque
% Wild-type activity Origin DNA
binding Single-strandedDNA binding Origin DNAunwindinga ATPase
308 Lys3Ala None 116 93 90 50
347 Lys3Ala Big NDb ND ND ND
349 Arg3Ala Small 105 38 61 80
364 Arg3Ala Small 91 36 46 35
371 Arg3Ala Small 121 122 111 95
377 Gly3Ala Small 111 92 105 90
387 Trp3Ala Big ND ND ND ND
429 Asp3Ala None 88 11 9 75
460 Glu3Ala None 83 0 0 29
aThe reactions were performed by incubating 400 ng of T antigen with an
end-labeled 112-bp origin containing DNA fragment in the presence ofE. coli
single-stranded DNA binding protein.
bND, not done.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.311.550.568.694.2]and 460EA are defective in DNA unwinding. One is due to their impairment in single-stranded DNA binding; the other is as a result of an inability to hydrolyze ATP. To distinguish between them, the mutants were tested for their ATPase ac-tivity. It is clear that all mutants were able to hydrolyze ATP and release inorganic phosphate (Fig. 3 and Table 1). Since the level of ATPase activity did not correlate with unwinding, it is unlikely that the mutants’ defects in unwinding were due to altered ATPase activity.
Single-stranded DNA binding activity of 429DA and 460EA. Since mutants 429DA and 460EA were very defective in single-stranded DNA binding and origin unwinding and still had ATPase activity, we decided to use these two mutants to study the function of single-stranded DNA binding. The mutant pro-teins appeared to be stable on the basis of our ability to purify normal amounts from infected insect cells. We first studied their binding to single-stranded DNA in more detail. The sin-gle-stranded DNA binding assays described earlier were per-formed at low T-antigen concentrations, and we first asked if this defect could be compensated by high protein concentra-tions. Higher amounts of mutant 429DA T antigen did par-tially restore binding to single-stranded DNA (Fig. 4, lanes 5, 6, and 7), but binding was still weaker than that of the wild type (Fig. 4, lanes 2, 3, and 4). However, mutant 460EA showed no binding to single-stranded DNA at all T-antigen concentra-tions tested (Fig. 4, lanes 8, 9, and 10), demonstrating that this mutant had an intrinsic defect in its interaction with single-stranded DNA.
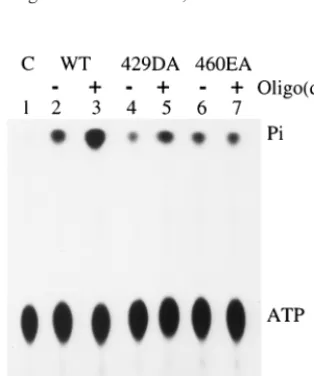
ATPase activity of 429DA and 460EA in the presence of single-stranded DNA.The ATPase activity of T antigen can be stimulated by single-stranded DNA (20, 47). If a mutant can-not bind to single-stranded DNA, one might predict that the addition of single-stranded DNA should have no effect on its ATPase activity. This was exactly what we observed with mu-tant 460EA. The addition of oligo(dT) did not cause an in-crease in ATPase activity (Fig. 5, lanes 6 and 7). In compari-FIG. 2. Single-stranded and origin DNA binding assays of
[image:4.612.314.543.72.262.2]wild-type (WT) and mutant T antigens. Four hundred nanograms of puri-fied wild-type and single-point-substitution mutant T antigens were tested for binding to end-labeled single-stranded DNA (SSDNA) or 112-bp origin-containing DNA (Ori DNA). (A) Binding by wild-type, 308KA, 371RA, and 377GA T antigens. (B) Binding by wild-type, 349RA, 364RA, 429DA, and 460EA T antigens. The positions of free DNA and DNA bound with T antigen in hexamers (H) and double hexamers (DH) are indicated.
FIG. 3. ATPase assay of wild-type (WT) and mutant T antigens. Six hundred nanograms of wild-type or mutant T antigens was incubated with 0.3 nmol of [␥-32P]ATP for 1 h at room temperature. The released free inorganic phosphate (Pi) was separated from the ATP substrate by ascending thin-layer chromatography.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.59.284.83.620.2]son, the ATPase activity of wild-type T antigen was stimulated 3.5-fold with oligo(dT), and that of mutant 429DA, which had weak binding to single-stranded DNA, was stimulated by two-fold (Fig. 5, lanes 2 to 5). This confirms that mutant 429DA effectively binds some single-stranded DNA, whereas mutant 460EA is completely defective.
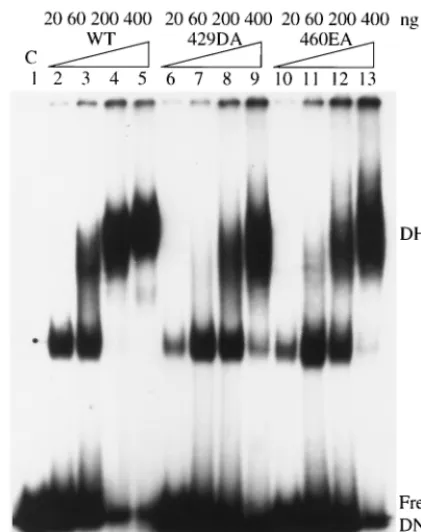
Unwinding activity of mutants 429DA and 460EA.We next studied the effects of higher T-antigen concentrations on DNA unwinding. Mutant 429DA showed very little origin unwinding at low T-antigen concentrations, but as its concentration was
increased, unwinding activity also increased (Fig. 6A, lanes 6, 7, and 8). However, mutant 460EA failed to unwind origin DNA even at high T-antigen concentrations (Fig. 6A, lanes 9, 10, and 11). Similar results were obtained with a standard helicase reaction measuring the ability to displace a primer from M13 single-stranded DNA. The helicase activity of mu-tant 429DA increased with increasing amounts of T antigen (Fig. 6B, lanes 6, 7, and 8), while mutant 460EA showed es-sentially no activity (Fig. 6B, lanes 9, 10, and 11). These results reveal a strong correlation between single-stranded DNA bind-ing and the ability to unwind DNA.
[image:5.612.321.542.72.370.2]Origin DNA binding by mutants 429DA and 460EA. We then asked if mutants 429DA and 460EA interact with origin DNA normally. At low T-antigen concentrations, wild-type T antigen bound to origin DNA as low-molecular-weight oli-gomers (Fig. 7, lane 2). As the concentration of T antigen increased, oligomers with higher molecular weights formed, eventually leading to double hexamers (Fig. 7, lanes 3, 4, and 5). First, we demonstrated that mutants 429DA and 460EA displayed similar patterns of origin binding with increasing amounts of T antigen (Fig. 7, lanes 6 to 13). At low T-antigen concentrations, binding was slightly weaker than that with the FIG. 4. Single-stranded DNA binding assay of wild-type (WT) and
mutant T antigens (Tag). Increasing amounts of wild-type, 429DA, and 460EA T antigens were tested for their ability to bind to single-stranded DNA. The positions of free DNA and bound DNA are shown. C, control.
[image:5.612.81.261.73.296.2]FIG. 5. Effect of oligo(dT) on the ATPase activity of wild-type (WT) and mutant T antigens. Six hundred nanograms of T antigen was incubated with 15 nmol of [␥-32P]ATP in the presence or absence of 40 ng of oligo(dT)32. C, control.
FIG. 6. Unwinding assays of wild-type (WT) and mutant T antigens (Tag). (A) Increasing amounts of wild-type, 429DA, and 460EA T antigens were tested for their ability to unwind a doubly end-labeled origin-containing DNA fragment in the presence ofE. coli single-stranded DNA binding protein. The positions of the single-single-stranded DNAs (SS) and double-stranded DNA (DS) are indicated. (B) In-creasing amounts of wild-type, 429DA, and 460EA T antigens were incubated with a helicase substrate. The positions of the annealed and displaced primers are indicated. C, control.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.95.252.499.687.2]wild type, suggesting that there is a partial defect in coopera-tive binding between individual monomers. Second, we showed that the abilities of mutants 429DA and 460EA to form double hexamers on origin DNA were stimulated with ATP just like the wild type (data not shown). These two experiments dem-onstrate that the origin binding and ATP binding domains of the mutants are functional and imply that the overall structure of each mutant is not dramatically altered.
Structural distortion activity of mutants 429DA and 460EA. When T antigen binds to the replication core origin, it melts the DNA at the EP on the early side of the central pentanucle-otides and untwists the AT track on the late side. These struc-tural alterations require ATP binding but not ATP hydrolysis (4). Since these changes may be accompanied by interactions with individual strands (51), we tested the possibility that sin-gle-stranded DNA binding is needed for structural distortion. Mutants 429DA and 460EA were therefore tested in a KMnO4
oxidation assay to detect DNA structure changes. Mutant 429DA caused very little EP or AT modification at low T-antigen concentrations (Fig. 8, lane 5), but at higher T con-centrations both EP and AT signals increased severalfold (Fig. 8, lanes 6 and 7). For comparison, the EP signal induced by wild-type T antigen remained about the same with increasing amounts of T antigen, while the AT signal increased several-fold (Fig. 8, lanes 2, 3, and 4). For mutant 460EA, there was no EP or AT signal (Fig. 8, lanes 8, 9, and 10) at all T-antigen concentrations. This shows a good correlation between single-stranded DNA binding and structure distortion.
DISCUSSION
The single-stranded DNA binding domain has been mapped previously to amino acid residues 259 to 627 (77). In this report we further mapped this domain by using internal deletion mutants. Our results showed that mutants with deletion of residues 251 to 300 or 501 to 550 still bound to single-stranded DNA; however, other mutants with deletions between residues 301 and 500 failed to bind to single-stranded DNA. This sig-nifies that the single-stranded DNA binding domain probably begins after residue 301 and that the region between 501 and 550 is dispensable. Our working model is that this domain lies between residues 301 and 500.
A number of single-point substitutions were introduced within this region, and several mutants were shown to be im-paired in single-stranded DNA binding. Two mutants, 429DA and 460EA, were the most defective. The mutant proteins appeared to assemble into double hexamers with origin DNA and displayed ATPase activity, suggesting that the origin bind-ing domain (residues 147 to 247) and ATP bindbind-ing domain (residues 418 to 528) were not seriously affected by the muta-tions. It was intriguing that 460EA was totally inactive in sin-gle-stranded DNA binding as well as in its ability to unwind DNA and to structurally distort the origin. Our interpretation is that this site is critical for an interaction with single-stranded DNA, perhaps because it is required for a structural change in T antigen during binding.
T antigen functions as a helicase during viral DNA replica-FIG. 7. Origin DNA binding assays of wild-type (WT) and mutant
[image:6.612.345.513.71.335.2]T antigens (Tag). Increasing amounts of wild-type, 429DA, and 460EA T antigens were incubated with an end-labeled DNA fragment con-taining the origin region. The positions of free DNA and DNA bound with T antigen in double hexamers (DH) are shown. C, control.
FIG. 8. KMnO4oxidation assay of wild-type (WT) and mutant T antigens (Tag). Increasing amounts of wild-type, 429DA, and 460EA T antigens were incubated with an origin-containing plasmid at 37°C for 30 min. The improperly paired thymine bases were modified by KMnO4and detected by primer extension using a32P-labeled primer. The melting signal at the EP and untwisting signal at the AT track are indicated. C, control.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.65.276.73.339.2]tion (60, 75). Based primarily on footprinting data, a model had been proposed describing how T antigen associates with DNA during unwinding. T antigen forms a double hexamer, with each strand of DNA passing through the central channel of one hexamer and wrapping around the other hexamer (see Fig. 9C) (18, 50, 51, 58). Thus, interactions with single-stranded DNA may be critical for DNA unwinding. Our ob-servation that there is a strong correlation between single-stranded DNA binding by 429DA and 460EA and origin unwinding as well as with helicase activity supports this hypoth-esis. Other hexameric helicases, like bacteriophage T7 heli-case, also form a ring structure with one strand of DNA threaded through the hexameric ring and the other lying out-side the ring (17, 23). Papilloma virus E1 probably forms a similarly shaped hexamer with single-stranded DNA (48). The crystal structures of helicases PcrA (69), hepatitis C virus NS3 (28), and Rep (29) bound to single-stranded DNA also dem-onstrate the importance of single-stranded DNA binding in helicase activity.
Based on these results, there are now at least five separate specific defects in T antigen that lead to an inability to unwind origin DNA. These include defects in origin binding (54, 55), in hexamer-hexamer interaction (37, 72), in oligomerization (73, 79), in ATPase activity (11, 38, 75), and now, in single-stranded DNA binding (this report).
Mutants 429DA and 460EA also showed correlation be-tween single-stranded DNA binding and the ability to cause structure distortion of origin DNA, suggesting that single-stranded DNA binding is also needed for structural distortion. There are several pieces of evidence that suggest that struc-tural distortion is a separate reaction from origin binding in-volving different regions of the protein. First, the DNA struc-ture of an origin fragment without the two pairs of GAGGC pentanucleotide binding sites can be altered by T antigen (41), indicating that origin binding by the origin binding domain is not required for structure changes. Second, T-antigen mutants that fail to bind to origin DNA can still cause structure distor-tion (55). Third, the interacdistor-tions between T antigen and the EP and AT regions are mostly through the sugar-phosphate back-bone (51), as they are between T antigen and single-stranded DNA (50), whereas interactions with the central pentanucle-otides involve direct binding to the bases (15, 24, 51, 65). Other single-stranded DNA binding proteins and helicases, such as RPA, PcrA, Rep, and NS3, also interact with the sugar-phos-phate backbone when they bind to single-stranded DNA (3, 28, 29, 69). Finally, the electron microscopy image of a T-antigen double hexamer bound to origin DNA (68) suggests that the two hexamers are arranged in a head-to-head orientation, with DNA in the central channel. Each hexamer has a bilobed structure, with the N-terminal DNA binding domain forming a small lobe that is bound to a pair of pentanucleotides and the remaining C-terminal region forming a larger lobe that covers the EP or AT region (see Fig. 9A). The single-stranded DNA binding domain is located in this second region. Our finding that single-stranded binding is involved in structure distortion is consistent with this model.
When T antigen initially binds to origin DNA, the central channel of each hexamer is thought to contain double-stranded DNA. However, various models propose that there is only one strand of DNA in the central channel during unwinding. How
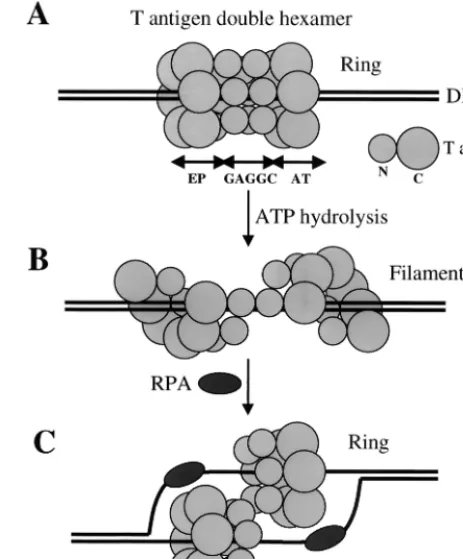
[image:7.612.315.547.76.356.2]one strand of DNA is displaced from the channel is not clear, but it probably involves a conformation change of the T-anti-gen hexamers. The recently resolved crystal structure of bac-teriophage T7 gene 4 helicase domain may provide an answer. T7 gene 4 helicase forms an asymmetric hexamer, where two subunits are rotated by 15° and two other subunits by 30° relative to the plane of the hexamer ring (57). A slightly smaller fragment of T7 gene 4 helicase domain forms a fila-ment instead of a closed ring (46).E. coliRecA, which has a similar nucleotide binding domain, can also form both rings and filaments (81), suggesting that rings and filaments are alternate structures that form during binding to DNA. Since the C-terminal region of T antigen also has homology with RecA (49), it is possible that the same changes occur in T antigen. In this model, a ring-to-filament conformation change would take place after the double hexamer forms on origin DNA (Fig. 9). This would permit one strand to be displaced and to associate with the single-stranded DNA binding protein RPA. Since there is evidence that T antigen interacts with only a single strand at the EP and AT regions (51), the DNA strand at each region might still be associated with T antigen through FIG. 9. A model showing possible conformation changes to T-an-tigen double hexamers bound to origin DNA. (A) T anT-an-tigen forms a double hexamer on origin DNA, with the DNA lying in the central channel of the closed hexameric ring on each hexamer. The N-terminal region of T antigen binds to one pair of GAGGC pentanucleotides, and the C-terminal region covers the EP or AT DNA element. The smaller and bigger circles represent the N-terminal (N) and C-terminal (C) regions of T antigen, respectively. (B) With ATP hydrolysis, the closed-ring structure of the double hexamer changes to an open fila-ment. (C) After one strand of DNA is displaced and bound by RPA, T antigen switches back to a closed ring to function as a helicase, with each hexamer encircling one strand of DNA.
on November 9, 2019 by guest
http://jvi.asm.org/
its single-stranded DNA binding activity after the transfer re-action. Afterwards, T antigen would reassemble into a hexam-eric closed ring to function as a helicase during DNA unwind-ing at replication forks.
In summary, there appear to be at least two steps during DNA replication where single-stranded DNA binding activity is required. One, this activity is needed shortly after DNA binding when the origin undergoes structure distortion (step A in Fig. 9). Two, it is needed during DNA unwinding in order to properly interact with the displaced strands (step C in Fig. 9).
ACKNOWLEDGMENTS
This work was supported by grants CA36118 from the National Cancer Institute and RR11820 from the National Center for Research Resources.
We are indebted to Judy Tevethia and Tim Kierstead for T-antigen internal deletion mutants. We thank Pamela Trowbridge for critical reading of the manuscript, and we are grateful to the University of Delaware cell biology facility for excellent cell culture and DNA se-quencing services.
REFERENCES
1.Arthur, A. K., A. Ho¨ss, and E. Fanning.1988. Expression of simian virus 40 T antigen inEscherichia coli: localization of T-antigen origin DNA-binding domain to within 129 amino acids. J. Virol.62:1999–2006.
2.Bird, L. E., H. S. Subramanya, and D. B. Wigley.1998. Helicases: a unifying structural theme? Curr. Opin. Struct. Biol.8:14–18.
3.Bochkarev, A., R. A. Pfuetzner, A. M. Edwards, and L. Frappier.1997. Structure of the single-stranded-DNA-binding domain of replication protein A bound to DNA. Nature385:176–181.
4.Borowiec, J. A., and J. Hurwitz. 1988. Localized melting and structural changes in the SV40 origin of replication induced by T-antigen. EMBO J. 7:3149–3158.
5.Borowiec, J. A., F. B. Dean, and J. Hurwitz.1991. Differential induction of structural changes in the simian virus 40 origin of replication by T antigen. J. Virol.65:1228–1235.
6.Bradley, M. K., J. Griffin, and D. M. Livingston.1982. Relationship of oligomerization to enzymatic and DNA binding properties of the SV40 large T antigen. Cell28:125–134.
7.Bradley, M. K., T. F. Smith, R. H. Lathrop, D. M. Livingston, and T. A. Webster.1987. Consensus topography in the ATP binding site of the simian virus 40 and polyomavirus large tumor antigens. Proc. Natl. Acad. Sci. USA 84:4026–4030.
8.Clark, R., D. P. Lane, and R. Tjian.1981. Use of monoclonal antibodies as probes of simian virus 40 T antigen ATPase activity. J. Biol. Chem.256: 11854–11858.
9.Clark, R., K. Peden, J. M. Pipas, D. Nathans, and R. Tjian.1983. Biochem-ical activities of T-antigen proteins encoded by simian virus 40 Agene deletion mutants. Mol. Cell. Biol.3:220–228.
10. Cole, C. N., J. Tornow, R. Clark, and R. Tjian.1986. Properties of the simian virus 40 (SV40) large T antigens encoded by SV40 mutants with deletions in geneA. J. Virol.57:539–546.
11. Collins, B. S., and J. M. Pipas.1995. T antigens encoded by replication-defective simian virus 40 mutants dl1135 and 5080. J. Biol. Chem.270:15377– 15384.
12. Dean, F. B., and J. Hurwitz.1991. Simian virus 40 large T antigen untwists DNA at the origin of DNA replication. J. Biol. Chem.266:5062–5071. 13. Dean, F. B., J. A. Borowiec, T. Eki, and J. Hurwitz.1992. The simian virus 40
T antigen double hexamer assembles around the DNA at the replication origin. J. Biol. Chem.267:14129–14137.
14. Dean, F. B., P. Bullock, Y. Murakami, C. R. Wobbe, L. Weissbach, and J. Hurwitz.1987. Simian virus 40 (SV40) DNA replication: SV40 large T antigen unwinds DNA containing the SV40 origin of replication. Proc. Natl. Acad. Sci. USA84:16–20.
15. DeLucia, A. L., B. A. Lewton, R. Tjian, and P. Tegtmeyer.1983. Topography of simian virus 40 A protein-DNA complexes: arrangement of pentanucle-otide interaction sites at the origin of replication. J. Virol.46:143–150. 16. Dodson, M., F. B. Dean, P. Bullock, H. Echols, and J. Hurwitz. 1987.
Unwinding of duplex DNA from the SV40 origin of replication by T antigen. Science238:964–967.
17. Egelman, H. H., X. Yu, R. Wild, M. M. Hingorani, and S. S. Patel.1995. Bacteriophage T7 helicase/primase proteins form rings around single-stranded DNA that suggest a general structure for hexameric helicases. Proc. Natl. Acad. Sci. USA92:3869–3873.
18. Falaschi, A.2000. Eukaryotic DNA replication: a model for a fixed double replisome. Trends Genet.16:88–92.
19. Gai, D., R. Roy, C. Wu, and D. T. Simmons.2000. Topoisomerase I associ-ates specifically with simian virus 40 large-T-antigen double hexamer-origin complexes. J. Virol.74:5224–5232.
20. Giacherio, D., and L. P. Hager.1979. A poly(dT)-stimulated ATPase activity associated with simian virus 40 large T antigen. J. Biol. Chem.254:8113– 8116.
21. Goetz, G. S., F. B. Dean, J. Hurwitz, and S. W. Matson.1988. The unwinding of duplex regions in DNA by the simian virus 40 large tumor antigen-associated DNA helicase activity. J. Biol. Chem.263:383–392.
22. Gorbalenya, A. E., E. V. Koonin, and Y. I. Wolf.1990. A new superfamily of putative NTP-binding domains encoded by genomes of small DNA and RNA viruses. FEBS Lett.262:145–148.
23. Hacker, K. J., and K. A. Johnson.1997. A hexameric helicase encircles one DNA strand and excludes the other during DNA unwinding. Biochemistry 36:14080–14087.
24. Jones, K. A., and R. Tjian.1984. Essential contact residues within SV40 large T antigen binding sites I and II identified by alkylation-interference. Cell 36:155–162.
25. Joo, W. S., H. Y. Kim, J. D. Purviance, K. R. Sreekumar, and P. A. Bullock. 1998. Assembly of T-antigen double hexamers on the simian virus 40 core origin requires only a subset of the available binding sites. Mol. Cell. Biol. 18:2677–2687.
26. Joo, W. S., X. Luo, D. Denis, H. Y. Kim, G. J. Rainey, C. Jones, K. R. Sreekumar, and P. A. Bullock.1997. Purification of the simian virus 40 (SV40) T antigen DNA-binding domain and characterization of its interac-tions with the SV40 origin. J. Virol.71:3972–3985.
27. Kierstead, T. D., and M. J. Tevethia.1993. Association of p53 binding and immortalization of primary C57BL/6 mouse embryo fibroblasts by using simian virus 40 T-antigen mutants bearing internal overlapping deletion mutations. J. Virol.67:1817–1829.
28. Kim, J. L., K. A. Morgenstern, J. P. Griffith, M. D. Dwyer, J. A. Thomson, M. A. Murcko, C. Lin, and P. R. Caron.1998. Hepatitis C virus NS3 RNA helicase domain with a bound oligonucleotide: the crystal structure provides insights into the mode of unwinding. Structure6:89–100.
29. Korolev, S., J. Hsieh, G. H. Gauss, T. M. Lohman, and G. Waksman.1997. Major domain swiveling revealed by the crystal structures of complexes of
E. coliRep helicase bound to single-stranded DNA and ADP. Cell90:635– 647.
30. Lin, H.-J. L., R. Upson, and D. T. Simmons.1992. Nonspecific DNA binding activity of simian virus 40 large T antigen: evidence for the cooperation of two regions for full activity. J. Virol.66:5443–5452.
31. Loeber, G., R. Parsons, and P. Tegtmeyer.1989. The zinc finger region of simian virus 40 large T antigen. J. Virol.63:94–100.
32. Masterson, P. J., M. A. Stanley, A. P. Lewis, and M. A. Romanos.1998. A C-terminal helicase domain of the human papillomavirus E1 protein binds E2 and the DNA polymerase alpha-primase p68 subunit. J. Virol.72:7407– 7419.
33. Mastrangelo, I. A., P. V. C. Hough, J. S. Wall, M. Dodson, F. B. Dean, and J. Hurwitz.1989. ATP-dependent assembly of double hexamers of SV40 T antigen at the viral origin of DNA replication. Nature338:658–662. 34. McCutchan, J. H., and J. S. Pagano.1968. Enhancement of the infectivity of
simian vius 40 deoxyribonucleic acid with diethylaminoethyldextran. J. Natl. Cancer Inst.41:351–357.
35. McVey, D., M. Strauss, and Y. Gluzman.1989. Properties of the DNA-binding domain of the simian virus 40 large T antigen. Mol. Cell. Biol. 9:5525–5536.
36. Melendy, T., and B. Stillman.1993. An interaction between replication protein A and SV40 T antigen appears essential for primosome assembly during SV40 DNA replication. J. Biol. Chem.268:3389–3395.
37. Moarefi, I. F., D. Small, I. Gilbert, M. Hopfner, S. K. Randall, C. Schneider, A. A. Russo, U. Ramsperger, A. K. Arthur, H. Stahl, T. J. Kelly, and E. Fanning.1993. Mutation of the cyclin-dependent kinase phosphorylation site in simian virus 40 (SV40) large T antigen specifically blocks SV40 origin DNA unwinding. J. Virol.67:4992–5002.
38. Mohr, I. J., M. P. Fairman, B. Stillman, and Y. Gluzman.1989. Large T-antigen mutants define multiple steps in the initiation of simian virus 40 DNA replication. J. Virol.63:4181–4188.
39. Murakami, Y., and J. Hurwitz.1993. Functional interactions between SV40 T antigen and other replication proteins at the replication fork. J. Biol. Chem.268:11008–11017.
40. Murakami, Y., C. R. Wobbe, L. Weissbach, F. B. Dean, and J. Hurwitz.1986. Role of DNA polymerase alpha and DNA primase in simian virus 40 DNA replication in vitro. Proc. Natl. Acad. Sci. USA83:2869–2873.
41. Parsons, R., M. E. Anderson, and P. Tegtmeyer.1990. Three domains in the simian virus 40 core origin orchestrate the binding, melting, and DNA helicase activities of T antigen. J. Virol.64:509–518.
42. Pfuetzner, R. A., A. Bochkarev, L. Frappier, and A. M. Edwards.1997. Replication protein A. Characterization and crystallization of the DNA binding domain. J. Biol. Chem.272:430–434.
43. Porter, D. J.1998. A kinetic analysis of the oligonucleotide-modulated ATPase activity of the helicase domain of the NS3 protein from hepatitis C
on November 9, 2019 by guest
http://jvi.asm.org/
virus. The first cycle of interaction of ATP with the enzyme is unique. J. Biol. Chem.273:14247–14253.
44. Prelich, G., C. K. Tan, M. Kostura, M. B. Mathews, A. G. So, K. M. Downey, and B. Stillman.1987. Functional identity of proliferating cell nuclear anti-gen and a DNA polymerase-delta auxiliary protein. Nature326:517–520. 45. Roberts, J. M., and G. D’Urso. 1989. Cellular and viral control of the
initiation of DNA replication. J. Cell Sci. Suppl.12:171–182.
46. Sawaya, M. R., S. Guo, S. Tabor, C. C. Richardson, and T. Ellenberger.1999. Crystal structure of the helicase domain from the replicative helicase-pri-mase of bacteriophage T7. Cell99:167–177.
47. Scheffner, M., R. Knippers, and H. Stahl.1989. RNA unwinding activity of SV40 large T antigen. Cell57:955–963.
48. Sedman, J., and A. Stenlund.1998. The papillomavirus E1 protein forms a DNA-dependent hexameric complex with ATPase and DNA helicase activ-ities. J. Virol.72:6893–6897.
49. Seif, R.1982. New properties of simian virus 40 large T antigen. Mol. Cell. Biol.2:1463–1471.
50. SenGupta, D. J., and J. A. Borowiec.1992. Strand-specific recognition of a synthetic DNA replication fork by the SV40 large tumor antigen. Science 256:1656–1661.
51. SenGupta, D. J., and J. A. Borowiec.1994. Strand and face: the topography of interactions between the SV40 origin of replication and T-antigen during the initiation of replication. EMBO J.13:982–992.
52. Simanis, V., and D. P. Lane.1985. An immunoaffinity purification procedure for SV40 large T antigen. Virology144:88–100.
53. Simmons, D. T., K. Wun-Kim, and W. Young.1990. Identification of simian virus 40 T antigen residues important for specific and nonspecific binding to DNA and for helicase activity. J. Virol.64:4858–4865.
54. Simmons, D. T., G. Loeber, and P. Tegtmeyer.1990. Four major sequence elements of simian virus 40 large T antigen coordinate its specific and nonspecific DNA binding. J. Virol.64:1973–1983.
55. Simmons, D. T., R. Upson, K. Wun-Kim, and W. Young.1993. Biochemical analysis of mutants with changes in the origin-binding domain of simian virus 40 tumor antigen. J. Virol.67:4227–4236.
56. Simmons, D. T., R. Roy, L. Chen, D. Gai, and P. W. Trowbridge.1998. The activity of topoisomerase I is modulated by large T antigen during unwinding of the SV40 origin. J. Biol. Chem.273:20390–20396.
57. Singleton, M. R., M. R. Sawaya, T. Ellenberger, and D. B. Wigley.2000. Crystal structure of T7 gene 4 ring helicase indicates a mechanism for sequential hydrolysis of nucleotides. Cell101:589–600.
58. Smelkova, N. V., and J. A. Borowiec.1998. Synthetic DNA replication bub-bles bound and unwound with twofold symmetry by a simian virus 40 T-antigen double hexamer. J. Virol.72:8676–8681.
59. Soultanas, P., M. S. Dillingham, P. Wiley, M. R. Webb, and D. B. Wigley. 2000. Uncoupling DNA translocation and helicase activity in PcrA: direct evidence for an active mechanism. EMBO J.19:3799–3810.
60. Stahl, H., P. Droge, and R. Knippers.1986. DNA helicase activity of SV40 large tumor antigen. EMBO J.5:1939–1944.
61. Stillman, B., R. D. Gerard, R. A. Guggenheimer, and Y. Gluzman.1985. T antigen and template requirements for SV40 DNA replication in vitro. EMBO J.4:2933–2939.
62. Subramanya, H. S., L. E. Bird, J. A. Brannigan, and D. B. Wigley.1996. Crystal structure of a DExx box DNA helicase. Nature384:379–383. 63. Tegtmeyer, P.1972. Simian virus 40 deoxyribonucleic acid synthesis: the viral
replicon. J. Virol.10:591–598.
64. Theis, K., P. J. Chen, M. Skorvaga, B. Van Houten, and C. Kisker.1999.
Crystal structure of UvrB, a DNA helicase adapted for nucleotide excision repair. EMBO J.18:6899–6907.
65. Tjian, R.1978. Protein-DNA interactions at the origin of simian virus 40 DNA replication. Cold Spring Harbor Symp. Quant. Biol.43:655–662. 66. Tjian, R.1978. The binding site of SV40 DNA for a T antigen-related
protein. Cell13:165–179.
67. Tsurimoto, T., and B. Stillman.1989. Purification of a cellular replication factor, RF-C, that is required for coordinated synthesis of leading and lag-ging strands during simian virus 40 DNA replication in vitro. Mol. Cell. Biol. 9:609–619.
68. Valle, M., C. Gruss, L. Halmer, J. M. Carazo, and L. E. Donate.2000. Large T-antigen double hexamers imaged at the simian virus 40 origin of replica-tion. Mol. Cell. Biol.20:34–41.
69. Velankar, S. S., P. Soultanas, M. S. Dillingham, H. S. Subramanya, and D. B. Wigley.1999. Crystal structures of complexes of PcrA DNA helicase with a DNA substrate indicate an inchworm mechanism. Cell97:75–84. 70. Waga, S., and B. Stillman.1994. Anatomy of a DNA replication fork
re-vealed by reconstitution of SV40 DNA replication in vitro. Nature (London) 369:207–212.
71. Weinberg, D. H., K. L. Collins, P. Simancek, A. Russo, M. S. Wold, D. M. Virshup, and T. J. Kelly.1990. Reconstitution of simian virus 40 DNA replication with purified proteins. Proc. Natl. Acad. Sci. USA87:8692–8696. 72. Weisshart, K., P. Taneja, A. Jenne, U. Herbig, D. T. Simmons, and E. Fanning.1999. Two regions of simian virus 40 T antigen determine cooper-ativity of double-hexamer assembly on the viral origin of DNA replication and promote hexamer interactions during bidirectional origin DNA unwind-ing. J. Virol.73:2201–2211.
73. Weisshart, K., M. K. Bradley, B. M. Weiner, C. Schneider, I. Moarefi, E. Fanning, and A. K. Arthur.1996. An N-terminal deletion mutant of simian virus 40 (SV40) large T antigen oligomerizes incorrectly on SV40 DNA but retains the ability to bind to DNA polymerase alpha and replicate SV40 DNA in vitro. J. Virol.70:3509–3516.
74. Wessel, R., J. Schweizer, and H. Stahl.1992. Simian virus 40 T-antigen DNA helicase is a hexamer which forms a binary complex during bidirectional unwinding from the viral origin of DNA replication. J. Virol.66:804–815. 75. Wiekowski, M., M. W. Schwarz, and H. Stahl.1988. Simian virus 40 large T
antigen DNA helicase. Characterization of the ATPase-dependent DNA unwinding activity and its substrate requirements. J. Biol. Chem.263:436– 442.
76. Wold, M. S., J. J. Li, and T. J. Kelly.1987. Initiation of simian virus 40 DNA replication in vitro: large-tumor-antigen- and origin-dependent unwinding of the template. Proc. Natl. Acad. Sci. USA84:3643–3647.
77. Wu, C., D. Edgil, and D. T. Simmons.1998. The origin DNA-binding and single-stranded DNA-binding domains of simian virus 40 large T antigen are distinct. J. Virol.72:10256–10259.
78. Wun-Kim, K., and D. T. Simmons.1990. Mapping of helicase and helicase substrate binding domains on simian virus 40 large T antigen. J. Virol. 64:2014–2020.
79. Wun-Kim, K., R. Upson, W. Young, T. Melendy, B. Stillman, and D. T. Simmons.1993. The DNA-binding domain of simian virus 40 tumor antigen has multiple functions. J. Virol.67:7608–7611.
80. Yao, N., T. Hesson, M. Cable, Z. Hong, A. D. Kwong, H. V. Le, and P. C. Weber.1997. Structure of the hepatitis C virus RNA helicase domain. Nat. Struct. Biol.4:463–467.
81. Yu, X., and E. H. Egelman.1997. The RecA hexamer is a structural homo-logue of ring helicases. Nat. Struct. Biol.4:101–104.