JOURNAL OFVIROLOGY,Apr. 1975,p.812-819
Copyright 0 1975 AmericanSociety for Microbiology
Vol. 15, No.4 Printed in U.S.A.
Temperature-Sensitive
Host
Range
Mutants of
Herpes Simplex
Virus
Type
2
ROGER W. KOMENT AND FRED RAPP*
Department of Microbiology, College of Medicine,Milton S.HersheyMedical Centerof PennsylvaniaState
University, Hershey, Pennsylvania 17033
Received for publication 23 September 1974
Herpesviruses are capable of several types of infection of a host cell. To investigate the early events which ultimately determine the nature of the virus-host cellinteraction, asystemwasestablishedutilizing temperature-sensi-tive mutants ofherpes simplex virus type 2. Four mutants have been isolated which fail to induce cytopathic effects and donot replicate at 39 C inhamster embryo fibroblast cells. At least one mutant is virus DNA negative. Since
intracellular complementation is detectable between pairs of mutants, a virus function is knowntobetemperaturesensitive. However,all four mutantsinduce cytopathic effects and replicate toparentalvirus levels in rabbit kidney cells at 39 C. This suggests that a host cell function, lacking or nonfunctional in HEF cells but present inrabbitkidney cellsat39 C, isrequiredforthereplication of thesemutantsinhamsterembryo fibroblast cellsat39 C.Therefore,weconclude that these mutants are both temperature sensitive and exhibit host range properties.
Acharacteristicofherpes simplexvirus infec-tionof ahost cellisthediversityofrelationships which may be established within that cell.The most common effect is the
production
of the cytolytic cycle which has been described bymany investigators for many cell systems (7, 24). Virus latency has long been known tobea property ofthe herpesviruses (13). This prop-erty ofherpessimplexvirustype 1 (HSV-1) has
recently been characterized in both animal (3,
27-29) and human (1, 2) systems. Recurrent
infections due to herpes simplex virus type 2
(HSV-2) (18)arewell known.A recent reportby
Walz et al. (31) has given further evidence of
such HSV-2 latency in experimental systems.
The scope of herpesvirus latency has recently been reviewed (22).
The suspected transforming and oncogenic
potential ofthe herpesviruses, with all its im-portantimplications, hasbeendemonstrated in manylaboratories
(In
E. Kurstak and K. Mara-morosch (ed.), Viruses, evolution, and cancer, in press). Although these three manifestations oftransformation, latency, and cytolytic infec-tionby herpessimplex viruses are underinvesti-gation, little is known of the control mech-anisms regulating events leading to different interactions. To delineate such control mech-anismswould greatly facilitate research efforts into virus latency and cell transformation.
Since both virus latency and transformation 81
require a more subtle interaction than the
complete destruction of an infected cell, we
sought
to establishavirus-cell system utilizingmutantsofHSV-2 which didnotinduce
cytopa-thology under nonpermissive conditions.
In this report we describe the isolation and
preliminary characterization offour mutants of
HSV-2 strain333 whichare temperature sensi-tive in hamster embryo fibroblast (HEF) cells
butshow a surprising hostrange property.
MATERIALS AND METHODS
Cell cultures. HEF cells were prepared from
12-day-old LSH strain (Lakeview Hamster Colony)
ham-sterembryos.Thecells were grown at 37 C in 8-ounce (0.473liter) glass prescription bottles in medium 199 supplementedwith10%fetal calf serum, 10% tryptose phosphate broth, and 0.075% sodium bicarbonate. Onehundred units ofpenicillin per milliliter and 100
jig
ofstreptomycin/ml
were used in all cell culturemedia. At confluency, cell monolayers were trypsin-ized and reseeded into plastic microwell trays (Linbro Disposo Trays, Linbro Chemical Co., New Haven, Conn.) andthe growth medium containing increased sodium bicarbonate (0.23%) was replaced. These culturesweregrowntoconfluency at 37 C in a 5%CO2 atmosphere.
Primary rabbit kidney (RK) cell cultures were preparedfromthe kidneys of 3-week-old New Zealand white rabbits. These cells were similarly grown at
37C in 8-ounce(0.473liter)glassprescription bottles
and passaged at confluency into plastic microwell
trays.Confluentmonolayersinplasticdishes(Falcon
on November 10, 2019 by guest
http://jvi.asm.org/
TEMPERATURE-SENSITIVE HOST RANGE MUTANTS
Plastics) (60 by15mm)wereusedfor infectiousvirus assay described below. All RK cells were grown in
Eagle minimal essential mediumsupplemented with
10%calfserum,0.075%or0.23%sodium bicarbonate,
and antibiotics. After virus inoculation, all cultures were maintained in Eagle medium supplemented
with2%fetalcalfserum,0.075%sodiumbicarbonate,
andantibiotics.
Virus. HSV-2 strain333 wasused astheparental
virus from which mutants were derived, and is
re-ferred to as the parental virus. This strain was
originally obtained from William Rawls (Baylor Col-lege of Medicine, Houston, Tex.) and had been
passaged in our laboratory four times in human embryokidney cells. A fifth passage washarvested,
quick frozen, and storedat -70Cforthese studies. Inoculation of cells. Confluent monolayers
con-sisting of 105 to 2 x 106 cells per microwell were
drained ofmedium andexposedto0.1mlof virus.For complementation studies, 0.05 ml each oftwo
mu-tantswasinoculated. Multiplicity of infectionforall
experiments was 2PFUpercell. After adsorption at roomtemperaturefor1h, cultureswerewashedtwice
with 1.0-ml volumes of 0.025 MTris-buffered saline (pH 7.4). One milliliter of Eagle medium containing 2%fetal calfserum, 0.23% sodiumbicarbonate, and
antibiotics was added to each microwell. Microwell
trayswereincubatedateither33,37, or39Cina5%
CO,atmosphere. The time ofvirus inoculation was
regardedas zerotime forallexperiments.
Plaque assay. Cultureswere frozen and thawed,
replicate cultureswerepooled, sonicallytreated for60 seachinaBransonSonifier(Cole-PalmerInstrument andEquipment Co.),andcelldebriswasremovedby centrifugation at200 x g. Serial 10-fold dilutions of each supernatantwere made in Tris-buffered saline
(pH 7.4) and 0.1 ml of each dilution of culture material wasinoculated intotwoplastic petri dishes
(60 by15mm) containing confluent monolayersof RK cells. After 1 h ofadsorptionatroomtemperatureall dishes were overlayed with 0.5% methylcellulose (Matheson, Coleman, and Bell) in Eagle medium supplementedwith5% fetalcalfserum,0.23% sodium
bicarbonate, and antibiotics. Infectious assays were
incubatedat33C in 5%CO2for4days, neutral red (diluted 1:7,500inTris-buffered saline)wasaddedfor
3h at37C, andplaqueswerecounted.
Mutagenesis by nitrosoguanidine (NTG).
Pa-rental virus(3.0x10 PFU)in 0.1 mlwasadded to0.9
mlof 50ug/mlofN-methyl-N-nitro-N-NTG (Kand K
Laboratories, Plainview,N.Y.) inmedium. At0.5, 1, 3, 5, 10, 20,and 60min,a0.1-mlsamplewasremoved from the incubation mixture and titrated directly in RK cells at 37 C as described above. At 4 days, plaqueswerecounted. Progenyfromplaquesof1mm
in diameteror less in size were picked and plaque purified at37Can additionaltwotimes.
Mutagenesis by UV irradiation. Parental virus
was diluted 1:4inTris-buffered saline and thevirus
suspension was sonicallytreated for 30 sto disrupt
virus aggregates. One milliliter ofthis virus
suspen-sion was placed into each of seven sterile plastic
dishes(60 by15mm)andexposedto4,000 ergs/sper
cm"of UV irradiation. At15-, 20-, 30-, 45-, 60-, 90-,
and 120-s exposure, one dish was removed and the contents weretitrated in RK cells at 37 C. Plaques of 1 mm diameter or less were picked and purified at
37C anadditionaltwotimes.
Mutagenesis by BUdR. Concentrations of 5, 10, and 20
;g
of5-bromodeoxyuridine (BUdR)/ml were prepared in Eagle medium containing 10% dialyzed fetal calf serum, 0.23% sodium bicarbonate, and antibiotics. Parental virus wasinoculated, at a multi-plicity of 3PFU/cell,onto RK cells indishes (60 by 15 mm) andallowed to adsorb at roomtemperature for 1 h. After adsorption, 5.0 ml of medium containing BUdR was added to each culture and all were incubated at 37 C in 5%CO,atmosphere. All cultures wereharvested at 24 h at which time virus cytopathic effect(CPE)wascomplete at all drugconcentrations. Cultures were quick frozen and thawed, sonically treated for 45 s, and cell debris was removed by low-speed centrifugation. Supernatants were held at-70C asmutagenized stocks.
DNAassay.Confluent monolayers of HEF cells in 1-ounce'(ca. 0.0591 liter) bottles (-5.0 x 10' total cells) were inoculated, virus was allowed to adsorb, and 2.0 ml of maintenance medium per bottle was
added. At this time, 10
ACi
of 3'-methyl-tritiated thymidine (specific activity 14.1 Ci per mmol; Schwarz Mann Co.) in 0.1-ml volume was added. Cultures were incubated at 33 and 39C for anadditional 23 h,atwhich time CPEwasobserved,and all labeled cultures were harvested by freezing at -4C.
Cultures werethawed, and lysateswerecombined
at 37 C with 0.1% Sarkosyl NL 30 (Geigy Chemical Corp.), 0.02 M ethylenediaminetetraacetic acid, and 0.1% heat-inactivated Pronase (Calbiochem, B grade). Pronase and Sarkosylwerepreparedin0.1 x SSC (0.1 x SSC: 0.015 M sodium chloride and0.0015
Msodium citrate, pH 7.3). Thedigestion mixturewas
allowed to incubate at 37C for a minimum of4 h, afterwhich0.2ml ofeachsamplewasmixed with 3.8 ml of cesium chloride in 0.1 x SSC (density equals 1.745g/cc).Thesewerecentrifugedat30,000 rpmfor
60 h at 20C in a Beckman L2-65B ultracentrifuge with a 40.3 fixed-angle rotor. Eight drop fractions werecollectedbybottom puncture of centrifuge tubes
ontoWhatman filter paperdisks.Acid-insoluble
ma-terial was precipitated by three washes with 5%
trichloroacetic acidat roomtemperature. Filterswere
dehydrated byonewashingwith 95%ethanol andone
washing with acetone. After drying, filters were
placedin 10ml oftoluenecontaining0.25%Omnifluor (New England Nuclear Corp.) and counted in a
Beckman LS-250 liquid scintillation counter. Every tenth fractionwascollected fordensitydetermination utilizingaBausch and Lomb refractometer.
RESULTS
Selection
by plaque
characteristic. Theparental
strain of HSV-2 333 was known toproduceinRK cellsa
heterogeneous
mixture ofplaque sizes ranging in diameter from 1 to 3 mm,largeplaques (>2mm)
being
thepredomi-nanttype. After treatment of
parental
viruses813
VOL.15, 1975
on November 10, 2019 by guest
http://jvi.asm.org/
KOMENTANDRAPP
with mutagens, an increase in the number of smallplaques(< 1 mm)wasobserved.Although
this was not quantitated atthetime,itoccurred regardless ofthe mutagenused. Thisfinding is consistent with other reports
indicating
thatvariousmutagens caninduceanincreased
num-ber of small plaques within the mutagenized
virus population (5, 14, 25, 30).
To obtain a readily detectable genetic marker, only those
plaques
<1 mmindiameterwere picked and purified
by
further titrationand selectioninRKcells.
Figure
1demonstrates this difference inplaquesize between predomi-nantly largeplaque-forming parental virusand homogeneous smallplaque-forming
mutants.The stability of small
plaque
size has been reportedfornonmutagenized isolates ofHSV-1 (20) andHSV-2 (16).Isolation ofmutants. In three separate ex-periments the parental virus was
subjected
to treatment with one form of mutagenic agent:UV irradiation,
BUdR,
or NTG. Aftermuta-genesis, the virus wastitrated
by
10-fold serial dilutionasdescribedinMaterials andMethods, and small plaques (<1 mm) were picked, andthen suspended in 1.0 ml of Eagle growth medium. There was a greater than 1,000-fold decreaseininfectious virus detectable after 120 s of UV irradiation
(Fig.
2). Mutant 69 wasderived from a single plaque picked from the
titration inRKcellsofparentalvirusexposedto 90 sofUV irradiation. This dosage representsa 0.1% survival rate. Figure 3 indicates that at concentrations of 5 to 20,ug per ml ofBUdR, infectivityofparentalvirus 333 is inhibited by 100-fold. Mutant74wasobtainedfrom a single
plaque formed upon titration in RK cells of parental virus 333 previously replicated in the
PARFNTAL.
:VIU:1AIV;T 69 MUTANT .4
FIG. 1. Plaques in RK cells of parental HSV-2 strain333and mutants.
10
-2
z1
z
a
L)
106
4
10 PARENTAL
3 3 3
Q3
103 I
0 30 60 90 120
TIME IN SECONDS
FIG. 2. Inactivation ofparental HSV-2 strain 333 after exposureto UW irradiation.
-i
(2
-I
z D
0 6 10 .
105_ PARENTAL
3 3 3
5 10
O9g BUDR PER ML OF MEDIUM
FIG. 3. Inactivation of parental HSV-2 strain 333 by replication in the presence of 5'-BUdR.
814 J.- VIROL.
17
14I
13
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.507.266.452.72.338.2] [image:3.507.267.449.370.633.2] [image:3.507.64.248.482.624.2]TEMPERATURE-SENSITIVE HOST RANGE MUTANTS
presence of 20 gg ofBUdRper ml of nutrient
medium.
Exposure of parentalvirustoNTGfor 60min resulted in only a four-fold decrease in initial
virus titer. It wasatthis point that mutant 46 was isolated from a single plaque. Mutant 41
was isolated similarly from a single plaque
picked from the titration in RKcells of parental virus 333exposed to 10 minofNTG. Since the parental virus was not plaque purified before mutagenesis, the possibility that a
spontane-ously arising variant has been selected cannot
beruledout.
All plaques were purified by repeated titra-tion and selection. Small plaque-purified iso-lateswerethen screenedat33 and39 C inRK and HEF cells for their abilitytoinduce CPEat
the permissive but not at the nonpermissive temperature. Parental virus will induce
com-plete CPE at both temperatures with equal efficiency.
Cytopathologyand infectivity in HEFcells. Stocks of fourmutantswhichyieldednoCPEat
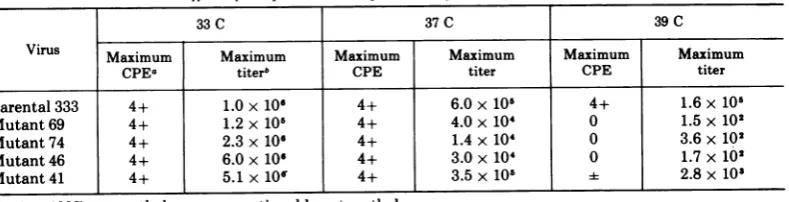
39 C inHEFcellswere growninRK cellsatthe permissive temperature and stored in aliquots at -70C. Table 1 demonstrates that these smallplaquemutants inducecomplete CPE by 48 h in HEF cells at 33C. Thiscytopathology correlates witha 1,000to10,000-fold increase in infectious virus between4 and 24 h (Fig. 4and 5), and is similar forall mutants tested.
Conversely, at 39 C in HEF cells, only the parental virus produces complete cytopathology by 24 h representing a 1,000-fold increase in infectious virus from the 4-h point (Fig. 4 and 5). Although there areslight amounts of leaki-ness, no mutant has progressed further than ±CPE at24 hor1+ at48 h.Lesssubjectiveare the actual titers representing infectious virus which demonstrate only a decrease in residual input virus. Table 1 further indicates that thesemutantsarecapable of inducing complete CPE but are less efficient in replicating at 37 thanat33 C.
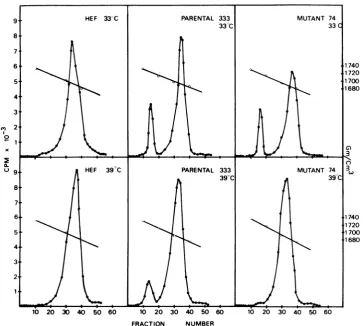
DNAresults. Mutant infectedHEF cultures werelabeled with 3'-methyltritiated-thymidine for24hatwhich time cellswereharvested and cell lysateswereanalyzed by isopycnic banding in cesium chloride gradients. By this method, virusDNA and host cell DNAcan beseparated basedontheircharacteristic densities. Figure 6 demonstrates that HEF cells inoculated with parental virus 333 synthesize virus DNAatboth 33 and 39C. However, mutant 74-inoculated HEFcells show littleor novirus DNAformation at 39C. Interestingly, at33 C, mutant 74
ap-pears todepress host cell DNAsynthesis (41%) but this characteristic is less marked at 39 C (7%).
Mutant 69 (Fig. 7) induces only small amountsof virus DNA in HEF cellsat33 Cover a 24-hcontinuous labelperiod. Moreover,some depression of host cell DNA synthesis is evident (21%). At 39 C, little or novirus DNA is made and there is a suggestion that host cell DNA synthesis is stimulated. Initial experiments also indicate that both mutants 46 and 41 (Fig. 7)
are capable of inducing reduced amounts of
virus DNAinHEFcellsat39 C anddepression of host cellDNAsynthesisoccursat both 33 and 39 C.
Cytopathology and infectivity of doubly infected HEF cells. To determine if the four
noncytopathic mutants derived from parental virus 333 were mutant in different cistrons, complementation studieswereperformed. HEF cells were mixedly infected with pairs of mu-tants,theviruswasallowed to adsorb for1 h at room temperature, the cell sheetswere washed twice with Tris-buffered saline, maintenance medium was added, and the cultures were
incubatedat33or39C.Inallcomplementation
experiments, singly infected HEF cells were included for calculation of complementation
levels.
Twoparameterswereutilized for the determi-nation of complementation: (i) formation of
[image:4.507.57.452.536.637.2]cytopathologyinmixedly infected cells and(ii)
TABLE 1. Effectoftemperatureonreplication ofHSV-2mutantsin HEF cells
33C 37C 39C
Virus Maximum Maximum Maximum Maximum Maximum Maximum
CPEa titerb CPE titer CPE titer
Parental333 4+ 1.0x10 4+ 6.0x105 4+ 1.6x105
Mutant69 4+ 1.2 x 106 4+ 4.0x 104 0 1.5x102
Mutant74 4+ 2.3x106 4+ 1.4x104 0 3.6x102
Mutant46 4+ 6.0x 106 4+ 3.0x 10' 0 1.7x102
Mutant41 4+ 5.1x10' 4+ 3.5x 105 2.8x10'
a4+, 100%cytopathology; -,questionablecytopathology.
t Maximumtiters inPFU per milliliter(checkedat24and48h
post-inoculation).
VOL.15,1975 815
on November 10, 2019 by guest
http://jvi.asm.org/
6.
105l
(2
z
10 a
'S
4 10
2
6
5
105
-j
2 4
Z 10
2
0
a
103
2
10
4 12 24 36 48 72 4 12 24 36 48 72
TIME IN HOURS TIME IN HOURS
FIG. 4. ReplicationofparentalHSV-2 strain 333 in FIG. 5. Replication of mutant 69 in HEF cells. HEF cells.
C',
0
C.)
10 20 30 40 50 60 10 20 30 40 50 60 10 20 30 40 50 60
FRACTION NUMBER
FIG. 6. DNA profiles'ofuninfected HEFcells and of cells infected withparental HSV-2 strain 333and
mutant74.
816
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.507.68.452.36.285.2] [image:5.507.66.453.55.509.2] [image:5.507.84.448.313.639.2]TEMPERATURE-SENSITIVE HOST RANGE MUTANTS
817
FRACTION NUMBER
FIG. 7. DNAprofiles of uninfected HEF cells and of cells infected with HSV-2mutants69, 46,and41.
replication of viruses mixedly infecting HEF cells at 39 C as compared to singly infected cells. To quantitate this latter parameter, the method of Burge andPfefferkorn (4)wasused. In thismethod, the 24-h infectious virus yields
were used. Complementation levels were de-rived by dividing infectious virus yields of mixedlyinfected cells by the sumofthe yields
of each mutant under identical conditions in singly infected cells. A complementation level greater than one indicates complementation. However,assuggested by Burge and Pfefferkorn (4), we considered only those values greater than two as indicative of complementation. Noncomplementing pairs of mutants produce complementation levels of one or less. Values less than one may indicate interference ofone mutant with the replication of the other. This meansofdetermining complementation values fortemperature-sensitivemutantsof HSV-1has alsorecentlybeen usedby Schaffer etal. (26).
By this method, two complementation groups, designatedAandB, have beendefined (Table 2). Thecombination of mutant 74 with
any other mutant induced complete
cytopa-TABLE 2. Complementation of HSV-2 mutants at
39C in HEF cells
Mutant Complementation Complementation
combination level group
74x 69 6.25
74x46 19.23 A 74
74x41 2.31
46x41 0.69 69
46x69 0.11 B 46
69x41 0.35 41
thology, and replication approaching parental virus yields under nonpermissive conditions. Anytwocombinations ofmutants41,46,and69 did not induce cytopathology nor replication
under nonpermissive conditions. In additionto
demonstrating that at least two different mu-tant genotypes exist, intracellular complemen-tation also indicates that these mutants are capableofadsorbing, penetrating, and uncoat-ing within the HEF cell, thereby makuncoat-ing availa-ble virus genetic information for interaction with the hostgenome.
VOL.15, 1975
C,,
I0
A.
C.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.507.63.432.59.396.2] [image:6.507.259.454.417.533.2]KOMENT AND RAPP
Regulation of replication by host celltype.
Tothis point, the mutantsdescribed havebeen considered as to their virus temperature-sensi-tive nature in HEF cells; the permissive tem-perature was 33 C and the nonpermissive tem-perature 39 C. It has been demonstrated that selected pairs of mutants are capable of com-plementation in mixedly infected HEF cells at 39 C, further supporting the conclusion that a temperature-sensitive virus function is in-volved.
The first suggestion that cell type is a deter-minant of permissiveness was observed when small plaque-purified isolates were screened at 33 and 39C in RK and HEF cells for their
ability to induce cytopathology at 39C. It was noted that while little or no cytopathology
occurred in HEFcells, complete cytopathology
eventually occurred in RK cells at 39C. In
returning to further investigate this unusual
behavior ofthetemperature-sensitive mutants,
itwasfoundthatmutants ofparentalvirus333, inadditiontoinducing complete cytopathology,
could also replicate in RK cells to levels
ap-proachingthat of parental virus under identical
conditions (Table3). This implies a host regula-tory mechanism aswellas a
temperature-sensi-tive virus function is involved.
DISCUSSION
The purpose of this study was to establish a system to study interactions between herpes
simplex virus and host cells. The approach taken was to isolate conditional lethal mutants ofHSV-2defective, undernonpermissive
condi-tions, in theirability to induce cytopathology. Before the development of this mutant virus system, studies on herpes virus latency in cell culture have been confined to observation of infected cultures to which the DNA inhibitor
TABLE 3. Replication of HSV-2in HEF andRK cells
at 39C
Hamsterembryo Rabbitkidney fibroblastcells cells
Virus Maximum Maximum
cytopa- taiterm cytopa- taiterb
thologya itr thologya itr
Parental 4+ 4.5 x 104 4- 2.8 x 105 333
Mutant 69 0 1.0 x 101 3- 2.9 x 104
Mutant74 0 1.5 x 102 4+ 3.5 x 104 Mutant46 0 3.1 x 102 3-- 8.5 x 104
Mutant41 1- 2.0 x 103 4- 7.0x 105 a4+, 100%; 3+,75%;1+,25%cytopathology. 'MeasuredinPFUper milliliter.
cytosine arabinoside has been added (12, 19). Preliminary studiesonvirus latencyhave been
attempted by infecting HEF cells with
non-cytopathic mutants and maintaining cultures at39C.Cultures have been maintained as
long
as6weekspost-inoculation. When cultureswere shifted downto permissivetemperature (33
C),
complete cytopathology was observed. The re-covered virus had mutant properties(Koment
and Rapp, unpublished data) and may there-forebeuseful in thestudyofviruslatencyin cell culture.
The unique observation that the mutants
replicated in RK cells at 39 C indicates that there may be a specific host component
re-quired for their replication. W. Munyon
(per-sonal communication) has made a similar ob-servation with mutants ofHSV-2. The extent of the host range property is underinvestigation.
The ability of herpes simplex viruses to in-duce heritable changes within cell populations
has been demonstrated in several laboratories
(6, 8, 9, 15, 17, 21, 23). This has been made possible in most cases by lethal treatment of virus by physical (UV irradiation) or chemical (neutral red) means before inoculation of cells tocircumvent the deleterious effects of cytopa-thology. Few transformation events have been reported using intact virus. Darai and Munk(6)
exposed human embryo lung cells to HSV-2,
shifted cultures to 42 C and recovered trans-formants. Garfinkle and McAuslan (10, 11)
"transformed" cells carrying Rous sarcoma virus with HSV type 1 or 2 and observed the expression of some virus functions. Utilizing
mutantsdescribedinthis study, transformation studies in HEF cells at 39 C could conceivably
identify a high frequency transforming mutant if the temperature-sensitive block occurred after transcription and translation of a "trans-forming" cistron (5). A variety of different
mutants,such asthose isolated, couldbe valua-ble in defining the virus information required for agiven transformational event.
ACKNOWLEDGMENTS
This work was supported by contract no. N01 CP53516 within the Virus Cancer Program of the National Cancer Instituteandby Public Health Serviceresearch grant no. 5 R01 CA11647 fromtheNational Cancer Institute.
LITERATURE CITED
1. Baringer, J. R., and P. Swoveland. 1973. Recovery of herpes-simplexvirusfrom human trigeminal ganglions. N.Engl.J.Med. 288:648-650.
2. Bastian,F. O., A. S. Rabson, C. L. Lee, and T. S. Tralka. 1972. Herpes-virus hominis: isolation from human tri-geminalganglion.Science178:306-307.
3. Benda, R., J. Cinatl, S. Petrovic, J. Roubal, and V.
818 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.507.63.255.503.634.2]TEMPERATURE-SENSITIVE HOST RANGE MUTANTS
Plaisner. 1973. Cultivation ofgangliaonmonofilfabric,
asuitable method for demonstration of latent herpes-virusinfection. Acta Virol. 17:305-309.
4. Burge, B. W., and E. R. Pfefferkorn.1966. Complemen-tation between temperature-sensitive mutants to
Sindbisvirus. Virology30:214-223.
5. Carp, R. I., and H. Koprowski. 1962.Mutationoftype3 polioviruswith nitrous acid. Virology17:99-109. 6. Darai, G., and K. Munk. 1973. Humanembryonic lung
cellsabortively infected with herpes virus hoministype
2show someproperties ofcell transformation. Nature
N.Biol. 241:268-269.
7. Darlington, R W., and A. Granoff. 1973. Replication-biological aspects, p. 93-220. In A. S. Kaplan (ed.), Theherpesviruses.Academic Press Inc., New York.
8. Duff, R., and F. Rapp. 1971. Oncogenic transformation of hamster cells after exposure to herpes simplex virus type2.Nature N. Biol. 233:48-50.
9. Duff, R., and F. Rapp. 1971. Properties of hamster embryo fibroblaststransformed in vitro afterexposure
toultraviolet-irradiated herpes simplex virustype2.J. Virol. 8:469-477.
10. Garfinkle, B., and B. R. McAuslan. 1973. Non-cyto-pathic, non-productive infection by herpes simplex virusestypes1and2.Intervirology 1:362-375. 11. Garfinkle, B., and B. R McAuslan.1974.Transformation
of cultured mammalian cellsby viable herpessimplex virussubtypes 1 and2. Proc. Nat. Acad. Sci. U.S.A. 71:220-224.
12. G6cz8l, E., and L. Vaczi. 1973.Cytomegalovirus latency incultured human cells. J. Gen. Virol. 18:143-151. 13. Goodpasture, E. W. 1929. Herpetic infection, with
espe-cialreference to involvement of the nervoussystem. Medicine 8:223-243.
14. Granoff, A. 1961. Induction of Newcastle disease virus
mutantswith nitrousacid. Virology 13:402-408. 15. Kutinovi, L., V. Vonka, and J. Broucek. 1973.Increased
oncogenicity and synthesis of herpes virus antigens in hamstercells exposedtoherpes simplex type-2virus. J.Nat.Cancer Inst. 50:759-765.
16. Munk, K., and G. Ludwig. 1972. Properties of plaque variants ofherpes virus hominis strains ofgenital origin. Arch.Gesamte Virusforsch. 37:308-315.
17. Munyon, W., E. Kraiselburd, D. Davis, and J. Mann. 1971.Transferofthymidine kinasetothymidine kinase-less L cells by infection with ultraviolet-irradiated herpes simplex virus. J. Virol. 7:813-820.
18. Naib, Z. M., A. J. Nahmias, W. E. Josey, and J. H.
Kramer. 1969. Genital herpetic infection. Association with cervical dysplasia and carcinoma. Cancer 23:940-945.
19. O'Neill, F. J., R. J. Goldberg, and F. Rapp.1972.Herpes simplexviruslatencyinculturedhumancellsfollowing
treatment with cytosine arabinoside. J. Gen. Virol. 14:189-197.
20. Rapp, F.1963.Variants of herpessimplexvirus: isolation characterization, and factors influencing plaque forma-tion.J.Bacteriol. 86:985-991.
21. Rapp,F., and R. Duff. 1973. Transformation of hamster embryo fibroblasts by herpes simplex viruses type 1 andtype2.CancerRes.33:1527-1534.
22. Rapp, F., and M. A. Jerkofsky. 1973. Persistent and latent infections, p. 271-289, InA. S. Kaplan (ed.),
Theherpesviruses. Academic Press Inc.,NewYork. 23. Rapp, F., J. H. Li, and M. Jerkofsky. 1973.
Transforma-tion of mammalian cells by DNA-containing viruses followingphotodynamic inactivation. Virology 55:339-346.
24. Roizman, B., P.G. Spear, and E.D.Kieff.1972.Herpes simplex viruses I and II: A biochemical definition, p.
129-188. In M. Pollard(ed.), Perspectives invirology VIII.Academic Press Inc., New York.
25. Ross, L. J.N., P. Wildy, and K. R Cameron. 1971. For-mation of small plaques by herpes viruses irradiated withultraviolet light. Virology 45:808-812.
26. Schaffer, P. A., G.M.Aron,N.Biswal, andM. Benyesh-Melnick. 1973. Temperature-sensitive mutants of herpessimplex virustype1:isolationcomplementation, andpartial characterization. Virology 52:57-71. 27. Stevens, J. G., and M. L. Cook. 1971. Latent herpes
simplex virusinspinalganglia of mice. Science 173:843-845.
28. Stevens, J.G., and M. L. Cook. 1973. Latentinfections induced by herpes simplex viruses. Cancer Res. 33: 1399-1401.
29. Stevens, J. G., A. B. Nesburn, and M. L. Cook. 1972. Latent herpes simplex virus from trigeminal ganglia of rabbits with recurrent eye infection. Nature N. Biol.235:216.
30. Thiry, L. 1963. Chemical mutagenesis of Newcastle diseasevirus.Virology 19:225-236.
31. Walz, M. A., R W. Price, and A. L. Notkins. 1974.
Latent ganglionic infection with herpes simplexvirus types1and2:viralreactivationinvivoafterneurectomy. Science184:1185-1187.
VOL.15,1975